Dill Extract Induces Elastic Fiber Neosynthesis and Functional Improvement in the Ascending Aorta of Aged Mice with Reversal of Age-Dependent Cardiac Hypertrophy and Involvement of Lysyl Oxidase-Like-1

Abstract
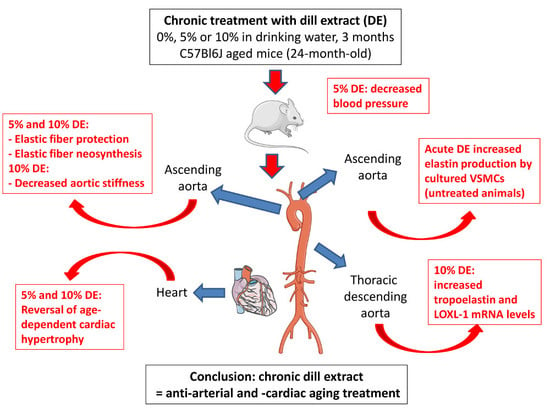
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Aqueous Extract of Dill and Treatment
2.3. Blood Pressure
2.4. Surgical and Post-Surgical Procedures
2.4.1. Heart Weight
2.4.2. Ascending Aorta Mechanics and Reactivity
2.4.3. Histological Examination
2.4.4. RNA Analyses
2.4.5. Elastin Production by Cultured VSMCs
Cell Culture
Extracellular Elastin Quantification
2.4.6. Statistics
3. Results
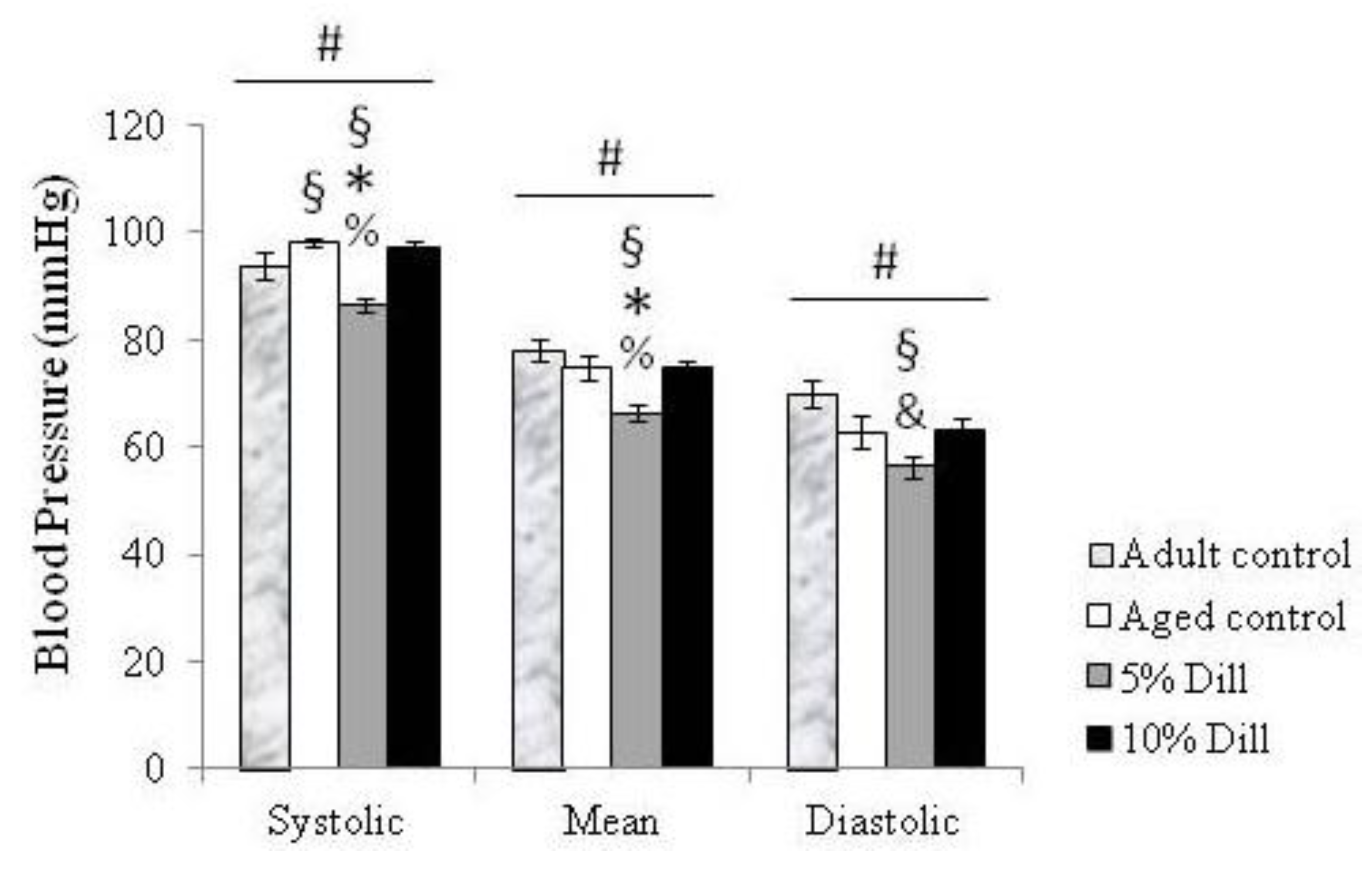
3.1. Blood Pressure
3.2. Body and Heart Weight Measurements
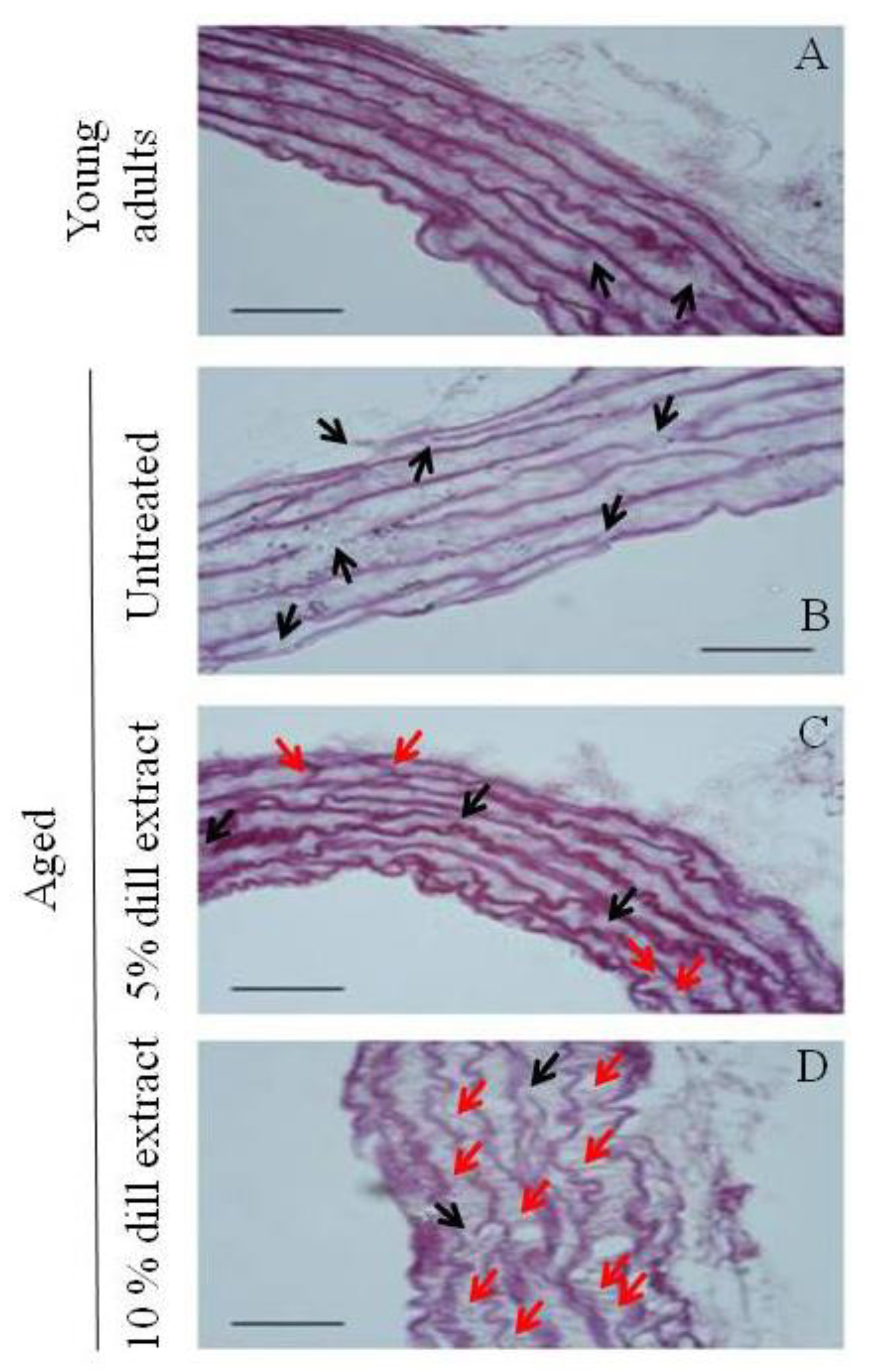
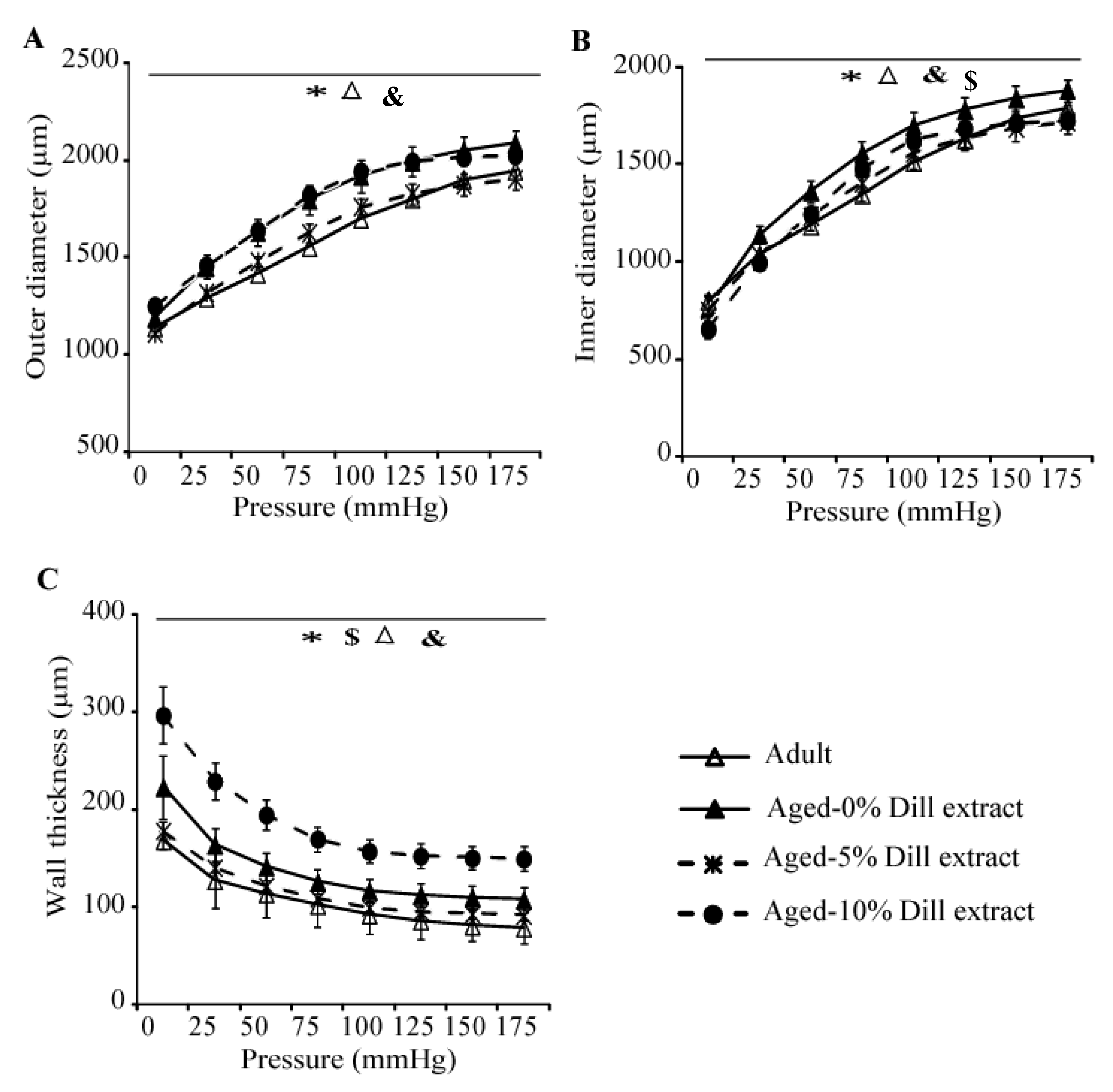
3.3. Ascending Aorta Morphology
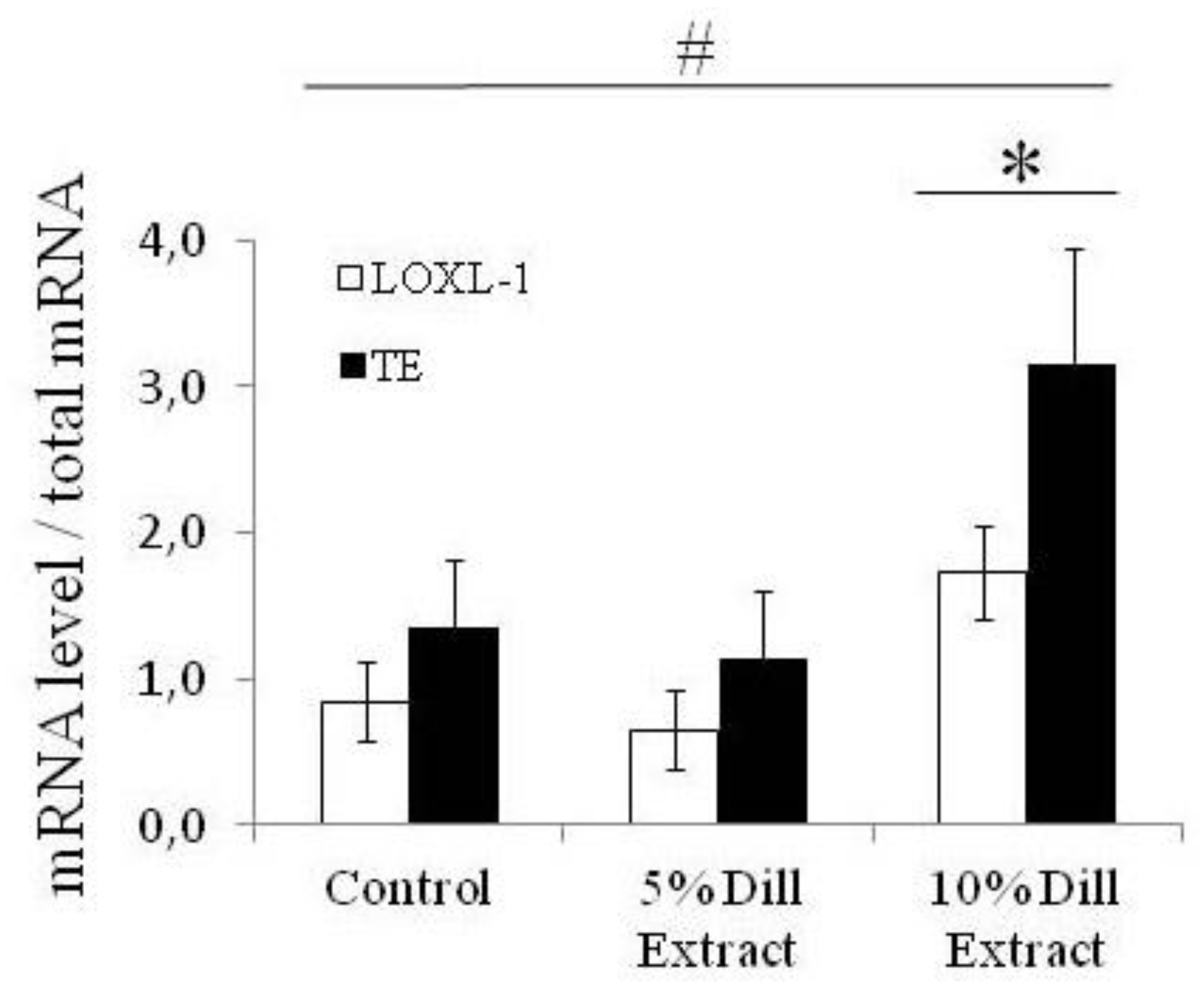
3.4. Tropoelastin and Lysyl-Oxidase Like-1 mRNA Levels
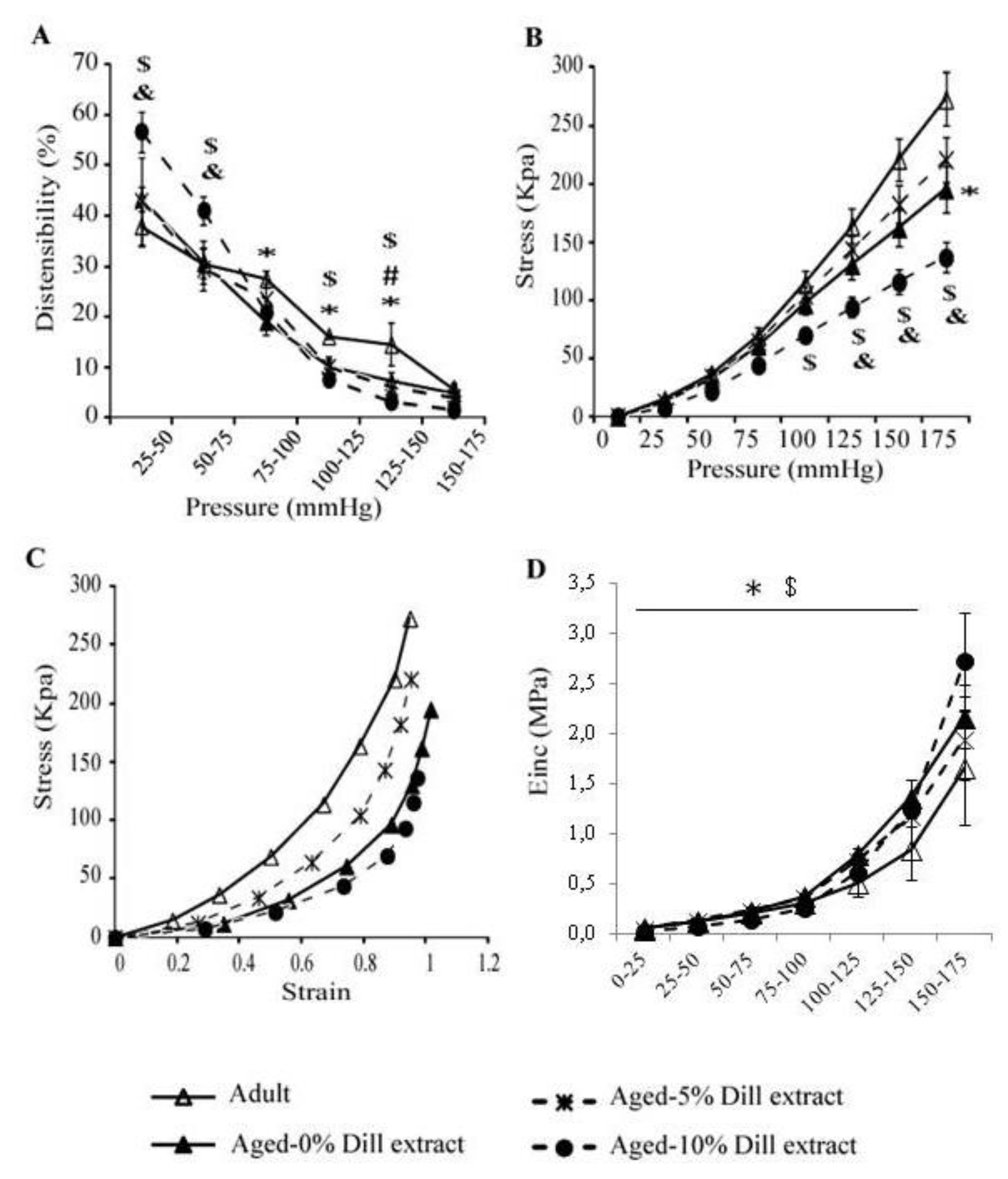
3.5. Biomechanical Properties of the Ascending Aorta
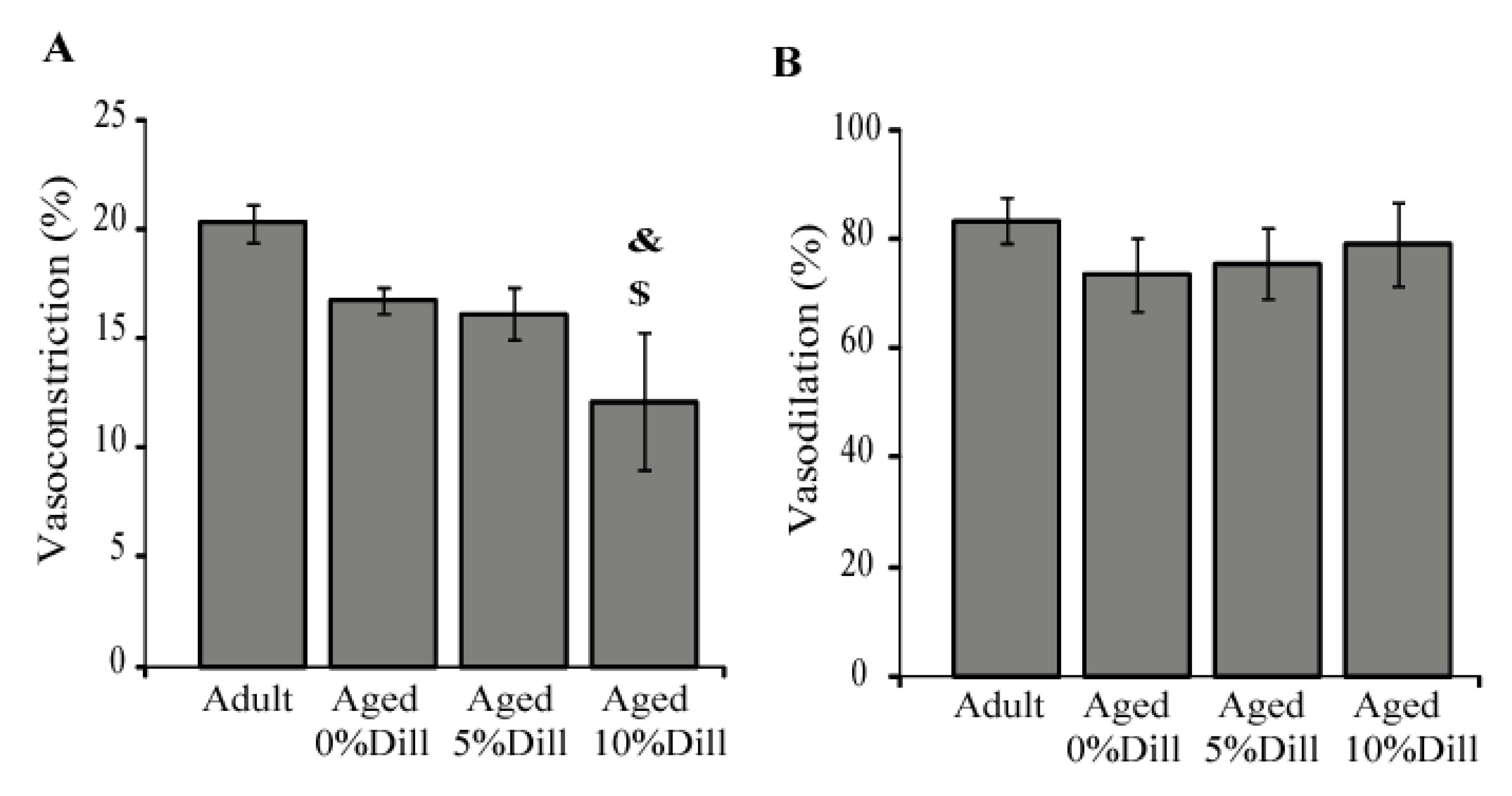
3.6. Ex Vivo Response of Ascending Aorta Segments to Vasoactive Agents
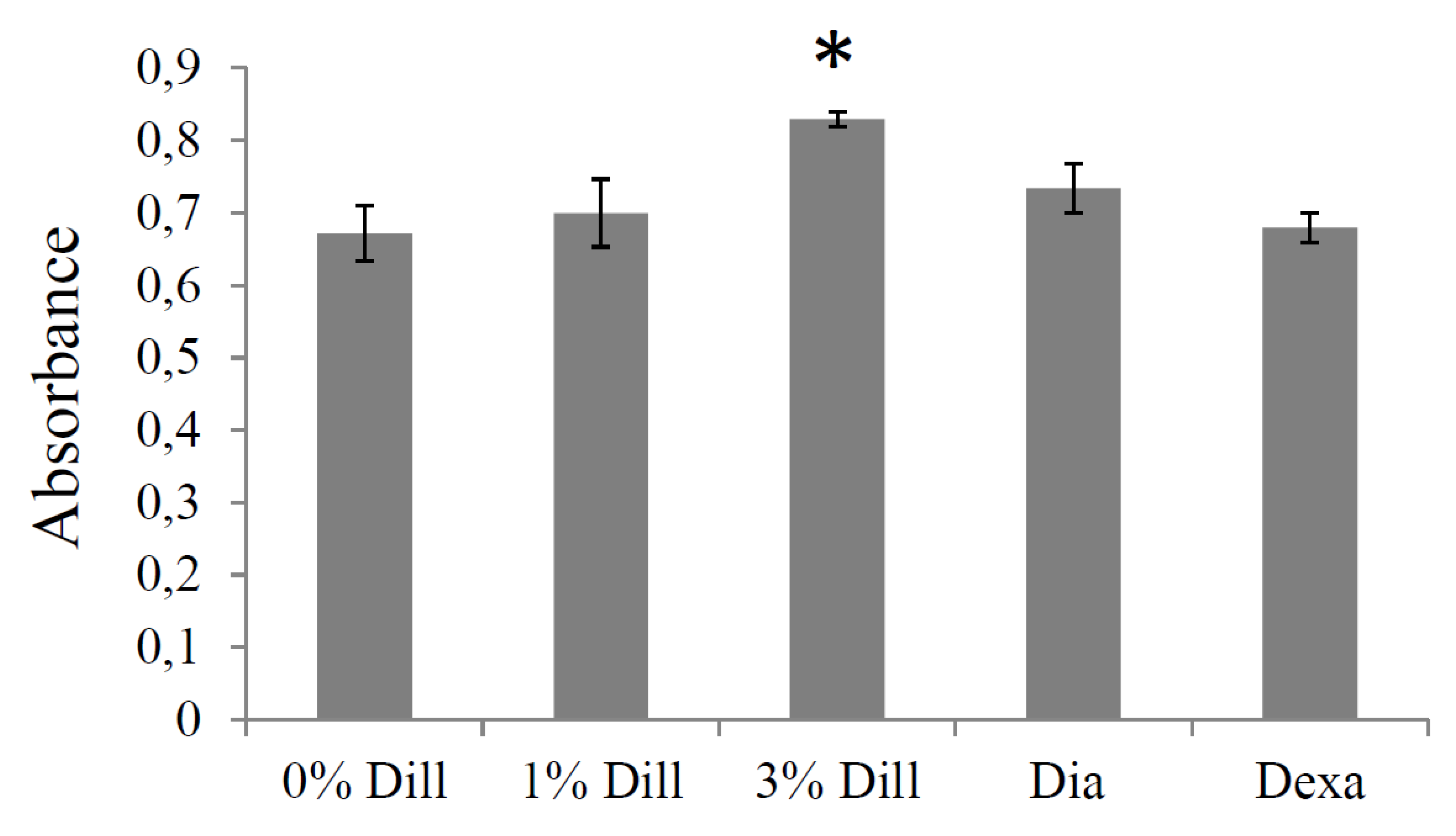
3.7. Impact of DE on Elastin Production by Cultured VSMCs
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A
Surgical Procedure and Mechanical Studies
References
- Zou, Y.; Zhang, Y. An experimental and theoretical study on the anisotropy of elastin network. Ann. Biomed. Eng. 2009, 37, 1572–1583. [Google Scholar] [CrossRef] [PubMed]
- Baldwin, A.K.; Simpson, A.; Steer, R.; Cain, S.A.; Kielty, C.M. Elastic fibres in health and disease. Expert Rev. Mol. Med. 2013, 15, e8. [Google Scholar] [CrossRef] [PubMed]
- Fhayli, W.; Ghandour, Z.; Mariko, B.; Pezet, M.; Faury, G. Elastin and microfibrils in vascular development and ageing: Complementary or opposite roles? Biol. Aujourdhui 2012, 206, 87–102. [Google Scholar] [CrossRef] [PubMed]
- Fhayli, W.; Boëté, Q.; Harki, O.; Briançon-Marjollet, A.; Jacob, M.-P.; Faury, G. Rise and fall of elastic fibers from development to aging. Consequences on arterial structure-function and therapeutical perspectives. Matrix Biol. 2019. [Google Scholar] [CrossRef] [PubMed]
- Dobrin, P.B. Mechanical properties of arteries. Physiol. Rev. 1978, 58, 397–460. [Google Scholar] [CrossRef] [PubMed]
- Mariko, B.; Pezet, M.; Escoubet, B.; Bouillot, S.; Andrieu, J.-P.; Starcher, B.; Quaglino, D.; Jacob, M.-P.; Huber, P.; Ramirez, F.; et al. Fibrillin-1 genetic deficiency leads to pathological ageing of arteries in mice. J. Pathol. 2011, 224, 33–44. [Google Scholar] [CrossRef]
- Barodka, V.M.; Joshi, B.L.; Berkowitz, D.E.; Hogue, C.W.; Nyhan, D. Review article: Implications of vascular aging. Anesth. Analg. 2011, 112, 1048–1060. [Google Scholar] [CrossRef]
- Wagenseil, J.E.; Mecham, R.P. Vascular extracellular matrix and arterial mechanics. Physiol. Rev. 2009, 89, 957–989. [Google Scholar] [CrossRef]
- Laurent, S.; Cockcroft, J.; Van Bortel, L.; Boutouyrie, P.; Giannattasio, C.; Hayoz, D.; Pannier, B.; Vlachopoulos, C.; Wilkinson, I.; Struijker-Boudier, H.; et al. Expert consensus document on arterial stiffness: Methodological issues and clinical applications. Eur. Heart J. 2006, 27, 2588–2605. [Google Scholar] [CrossRef]
- Li, D.Y.; Brooke, B.; Davis, E.C.; Mecham, R.P.; Sorensen, L.K.; Boak, B.B.; Eichwald, E.; Keating, M.T. Elastin is an essential determinant of arterial morphogenesis. Nature 1998, 393, 276–280. [Google Scholar] [CrossRef]
- Li, D.Y.; Faury, G.; Taylor, D.G.; Davis, E.C.; Boyle, W.A.; Mecham, R.P.; Stenzel, P.; Boak, B.; Keating, M.T. Novel arterial pathology in mice and humans hemizygous for elastin. J. Clin. Investig. 1998, 102, 1783–1787. [Google Scholar] [CrossRef] [PubMed]
- Faury, G.; Pezet, M.; Knutsen, R.H.; Boyle, W.A.; Heximer, S.P.; McLean, S.E.; Minkes, R.K.; Blumer, K.J.; Kovacs, A.; Kelly, D.P.; et al. Developmental adaptation of the mouse cardiovascular system to elastin haploinsufficiency. J. Clin. Investig. 2003, 112, 1419–1428. [Google Scholar] [CrossRef] [PubMed]
- Pezet, M.; Jacob, M.-P.; Escoubet, B.; Gheduzzi, D.; Tillet, E.; Perret, P.; Huber, P.; Quaglino, D.; Vranckx, R.; Li, D.Y.; et al. Elastin haploinsufficiency induces alternative aging processes in the aorta. Rejuvenation Res. 2008, 11, 97–112. [Google Scholar] [CrossRef] [PubMed]
- Johnson, D.J.; Robson, P.; Hew, Y.; Keeley, F.W. Decreased elastin synthesis in normal development and in long-term aortic organ and cell cultures is related to rapid and selective destabilization of mRNA for elastin. Circ. Res. 1995, 77, 1107–1113. [Google Scholar] [CrossRef] [PubMed]
- Pasquali-Ronchetti, I.; Baccarani-Contri, M. Elastic fiber during development and aging. Microsc. Res. Tech. 1997, 38, 428–435. [Google Scholar] [CrossRef]
- Liu, X.; Zhao, Y.; Gao, J.; Pawlyk, B.; Starcher, B.; Spencer, J.A.; Yanagisawa, H.; Zuo, J.; Li, T. Elastic fiber homeostasis requires lysyl oxidase-like 1 protein. Nat. Genet. 2004, 36, 178–182. [Google Scholar] [CrossRef]
- Hornstra, I.K.; Birge, S.; Starcher, B.; Bailey, A.J.; Mecham, R.P.; Shapiro, S.D. Lysyl oxidase is required for vascular and diaphragmatic development in mice. J. Biol. Chem. 2003, 278, 14387–14393. [Google Scholar] [CrossRef]
- Noblesse, E.; Cenizo, V.; Bouez, C.; Borel, A.; Gleyzal, C.; Peyrol, S.; Jacob, M.-P.; Sommer, P.; Damour, O. Lysyl oxidase-like and lysyl oxidase are present in the dermis and epidermis of a skin equivalent and in human skin and are associated to elastic fibers. J. Investig. Dermatol. 2004, 122, 621–630. [Google Scholar] [CrossRef]
- Thorin-Trescases, N.; Thorin, E. Lifelong Cyclic Mechanical Strain Promotes Large Elastic Artery Stiffening: Increased Pulse Pressure and Old Age-Related Organ Failure. Can. J. Cardiol. 2016, 32, 624–633. [Google Scholar] [CrossRef]
- McNulty, M.; Spiers, P.; McGovern, E.; Feely, J. Aging is associated with increased matrix metalloproteinase-2 activity in the human aorta. Am. J. Hypertens. 2005, 18, 504–509. [Google Scholar] [CrossRef]
- Jacob, M.P. Extracellular matrix remodeling and matrix metalloproteinases in the vascular wall during aging and in pathological conditions. Biomed. Pharmacother. 2003, 57, 195–202. [Google Scholar] [CrossRef]
- Duca, L.; Blaise, S.; Romier, B.; Laffargue, M.; Gayral, S.; El Btaouri, H.; Kawecki, C.; Guillot, A.; Martiny, L.; Debelle, L.; et al. Matrix ageing and vascular impacts: Focus on elastin fragmentation. Cardiovasc. Res. 2016, 110, 298–308. [Google Scholar] [CrossRef] [PubMed]
- Astrand, H.; Stalhand, J.; Karlsson, J.; Karlsson, M.; Sonesson, B.; Länne, T. In vivo estimation of the contribution of elastin and collagen to the mechanical properties in the human abdominal aorta: Effect of age and sex. J. Appl. Physiol. 2011, 110, 176–187. [Google Scholar] [CrossRef] [PubMed]
- Hodis, S.; Zamir, M. Mechanical events within the arterial wall: The dynamic context for elastin fatigue. J. Biomech. 2009, 42, 1010–1016. [Google Scholar] [CrossRef] [PubMed]
- Greenwald, S.E. Ageing of the conduit arteries. J. Pathol. 2007, 211, 157–172. [Google Scholar] [CrossRef] [PubMed]
- Basalyga, D.M.; Simionescu, D.T.; Xiong, W.; Baxter, B.T.; Starcher, B.C.; Vyavahare, N.R. Elastin degradation and calcification in an abdominal aorta injury model: Role of matrix metalloproteinases. Circulation 2004, 110, 3480–3487. [Google Scholar] [CrossRef] [PubMed]
- Mackenzie, I.S.; Wilkinson, I.B.; Cockcroft, J.R. Assessment of arterial stiffness in clinical practice. QJM 2002, 95, 67–74. [Google Scholar] [CrossRef]
- Lee, H.-Y.; Oh, B.-H. Aging and arterial stiffness. Circ. J. 2010, 74, 2257–2262. [Google Scholar] [CrossRef]
- Hayashi, A.; Suzuki, T.; Wachi, H.; Tajima, S.; Nishikawa, T.; Murad, S.; Pinnell, S.R. Minoxidil stimulates elastin expression in aortic smooth muscle cells. Arch. Biochem. Biophys. 1994, 315, 137–141. [Google Scholar] [CrossRef]
- Tsoporis, J.; Keeley, F.W.; Lee, R.M.; Leenen, F.H. Arterial vasodilation and vascular connective tissue changes in spontaneously hypertensive rats. J. Cardiovasc. Pharmacol. 1998, 31, 960–962. [Google Scholar] [CrossRef]
- Slove, S.; Lannoy, M.; Behmoaras, J.; Pezet, M.; Sloboda, N.; Lacolley, P.; Escoubet, B.; Buján, J.; Jacob, M.-P. Potassium channel openers increase aortic elastic fiber formation and reverse the genetically determined elastin deficit in the BN rat. Hypertension 2013, 62, 794–801. [Google Scholar] [CrossRef] [PubMed]
- Bouider, N.; Fhayli, W.; Ghandour, Z.; Boyer, M.; Harrouche, K.; Florence, X.; Pirotte, B.; Lebrun, P.; Faury, G.; Khelili, S. Design and synthesis of new potassium channel activators derived from the ring opening of diazoxide: Study of their vasodilatory effect, stimulation of elastin synthesis and inhibitory effect on insulin release. Bioorg. Med. Chem. 2015, 23, 1735–1746. [Google Scholar] [CrossRef] [PubMed]
- Coquand-Gandit, M.; Jacob, M.-P.; Fhayli, W.; Romero, B.; Georgieva, M.; Bouillot, S.; Estève, E.; Andrieu, J.-P.; Brasseur, S.; Bouyon, S.; et al. Chronic Treatment with Minoxidil Induces Elastic Fiber Neosynthesis and Functional Improvement in the Aorta of Aged Mice. Rejuvenation Res. 2017, 20, 218–230. [Google Scholar] [CrossRef] [PubMed]
- Bouhedja, M.; Peres, B.; Fhayli, W.; Ghandour, Z.; Boumendjel, A.; Faury, G.; Khelili, S. Design, synthesis and biological evaluation of novel ring-opened cromakalim analogues with relaxant effects on vascular and respiratory smooth muscles and as stimulators of elastin synthesis. Eur. J. Med. Chem. 2018, 144, 774–796. [Google Scholar] [CrossRef]
- Knutsen, R.H.; Beeman, S.C.; Broekelmann, T.J.; Liu, D.; Tsang, K.M.; Kovacs, A.; Ye, L.; Danback, J.R.; Watson, A.; Wardlaw, A.; et al. Minoxidil improves vascular compliance, restores cerebral blood flow, and alters extracellular matrix gene expression in a model of chronic vascular stiffness. Am. J. Physiol. Heart Circ. Physiol. 2018, 315, H18–H32. [Google Scholar] [CrossRef]
- Fhayli, W.; Boyer, M.; Ghandour, Z.; Jacob, M.P.; Andrieu, J.P.; Starcher, B.C.; Estève, E.; Faury, G. Chronic administration of minoxidil protects elastic fibers and stimulates their neosynthesis with improvement of the aorta mechanics in mice. Cell. Signal. 2019, 62, 109333. [Google Scholar] [CrossRef]
- Sohm, B.; Cenizo, V.; André, V.; Zahouani, H.; Pailler-Mattei, C.; Vogelgesang, B. Evaluation of the efficacy of a dill extract in vitro and in vivo. Int. J. Cosmet. Sci. 2011, 33, 157–163. [Google Scholar] [CrossRef]
- Cenizo, V.; André, V.; Reymermier, C.; Sommer, P.; Damour, O.; Perrier, E. LOXL as a target to increase the elastin content in adult skin: A dill extract induces the LOXL gene expression. Exp. Dermatol. 2006, 15, 574–581. [Google Scholar] [CrossRef]
- Behmoaras, J.; Slove, S.; Seve, S.; Vranckx, R.; Sommer, P.; Jacob, M.-P. Differential expression of lysyl oxidases LOXL1 and LOX during growth and aging suggests specific roles in elastin and collagen fiber remodeling in rat aorta. Rejuvenation Res. 2008, 11, 883–889. [Google Scholar] [CrossRef]
- Faury, G.; Maher, G.M.; Li, D.Y.; Keating, M.T.; Mecham, R.P.; Boyle, W.A. Relation between outer and luminal diameter in cannulated arteries. Am. J. Physiol. 1999, 277, H1745–H1753. [Google Scholar] [CrossRef]
- Smith, J.; Kampine, J. Circulatory Physiology—The Essentials, 3rd ed.; Williams & Wilkins: Baltimore, MD, USA, 1990. [Google Scholar]
- Gibbons, C.A.; Shadwick, R.E. Functional similarities in the mechanical design of the aorta in lower vertebrates and mammals. Experientia 1989, 45, 1083–1088. [Google Scholar] [CrossRef] [PubMed]
- Barnett, C.P.; Chitayat, D.; Bradley, T.J.; Wang, Y.; Hinek, A. Dexamethasone normalizes aberrant elastic fiber production and collagen 1 secretion by Loeys-Dietz syndrome fibroblasts: A possible treatment? Eur. J. Hum. Genet. 2011, 19, 624–633. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Moravec, C.S.; Ruhe, T.; Cifani, J.R.; Milovanovic, M.; Khairallah, P.A. Structural and functional consequences of minoxidil-induced cardiac hypertrophy. J. Pharmacol. Exp. Ther. 1994, 269, 290–296. [Google Scholar] [PubMed]
- Lowenthal, D.T.; Onesti, G.; Mutterperl, R.; Affrime, M.; Martinez, E.W.; Kim, K.E.; Busby, P.; Shirk, J.; Swartz, C. Long-term clinical effects, bioavailability, and kinetics of minoxidil in relation to renal function. J. Clin. Pharmacol. 1978, 18, 500–508. [Google Scholar] [CrossRef]
- Jana, S.; Shekhawat, G.S. Anethum graveolens: An Indian traditional medicinal herb and spice. Pharmacogn. Rev. 2010, 4, 179–184. [Google Scholar]
- Jana, S.; Shekhawat, G. Phytochemical Analysis and Antibacterial Screening of in vivo and in vitro Extracts of Indian Medicinal Herb: Anethum graveolens. Res. J. Med. Plant. 2010, 4, 206–212. [Google Scholar] [CrossRef]
- Angeloni, C.; Leoncini, E.; Malaguti, M.; Angelini, S.; Hrelia, P.; Hrelia, S. Role of quercetin in modulating rat cardiomyocyte gene expression profile. Am. J. Physiol. Heart Circ. Physiol. 2008, 294, H1233–H1243. [Google Scholar] [CrossRef]
- Quiñones, M.; Miguel, M.; Aleixandre, A. Beneficial effects of polyphenols on cardiovascular disease. Pharmacol. Res. 2013, 68, 125–131. [Google Scholar] [CrossRef]
- Testai, L.; Martelli, A.; Cristofaro, M.; Breschi, M.C.; Calderone, V. Cardioprotective effects of different flavonoids against myocardial ischaemia/reperfusion injury in Langendorff-perfused rat hearts. J. Pharm. Pharmacol. 2013, 65, 750–756. [Google Scholar] [CrossRef]
- Schroeter, H.; Heiss, C.; Balzer, J.; Kleinbongard, P.; Keen, C.L.; Hollenberg, N.K.; Sies, H.; Kwik-Uribe, C.; Schmitz, H.H.; Kelm, M. (-)-Epicatechin mediates beneficial effects of flavanol-rich cocoa on vascular function in humans. Proc. Natl. Acad. Sci. USA 2006, 103, 1024–1029. [Google Scholar] [CrossRef]
- Thring, T.S.A.; Hili, P.; Naughton, D.P. Anti-collagenase, anti-elastase and anti-oxidant activities of extracts from 21 plants. BMC Complement. Altern. Med. 2009, 9, 27. [Google Scholar] [CrossRef] [PubMed]
- Jimenez, F.; Mitts, T.F.; Liu, K.; Wang, Y.; Hinek, A. Ellagic and tannic acids protect newly synthesized elastic fibers from premature enzymatic degradation in dermal fibroblast cultures. J. Investig. Dermatol. 2006, 126, 1272–1280. [Google Scholar] [CrossRef] [PubMed]
- Negrão, M.R.; Keating, E.; Faria, A.; Azevedo, I.; Martins, M.J. Acute effect of tea, wine, beer, and polyphenols on ecto-alkaline phosphatase activity in human vascular smooth muscle cells. J. Agric. Food Chem. 2006, 54, 4982–4988. [Google Scholar] [CrossRef] [PubMed]
- Lomashvili, K.A.; Cobbs, S.; Hennigar, R.A.; Hardcastle, K.I.; O’Neill, W.C. Phosphate-induced vascular calcification: Role of pyrophosphate and osteopontin. J. Am. Soc. Nephrol. 2004, 15, 1392–1401. [Google Scholar] [CrossRef]
- Le Lièvre, C.S.; Le Douarin, N.M. Mesenchymal derivatives of the neural crest: Analysis of chimaeric quail and chick embryos. J. Embryol. Exp. Morphol. 1975, 34, 125–154. [Google Scholar]
- Jiang, X.; Rowitch, D.H.; Soriano, P.; McMahon, A.P.; Sucov, H.M. Fate of the mammalian cardiac neural crest. Development 2000, 127, 1607–1616. [Google Scholar]
- Lin, P.H.; Chaikof, E.L. Embryology, anatomy, and surgical exposure of the great abdominal vessels. Surg. Clin. N. Am. 2000, 80, 417–433. [Google Scholar] [CrossRef]
- Pfaltzgraff, E.R.; Bader, D.M. Heterogeneity in vascular smooth muscle cell embryonic origin in relation to adult structure, physiology, and disease. Dev. Dyn. 2015, 244, 410–416. [Google Scholar] [CrossRef]
- Gadson, P.F., Jr.; Dalton, M.L.; Patterson, E.; Svoboda, D.D.; Hutchinson, L.; Schram, D.; Rosenquist, T.H. Differential response of mesoderm- and neural crest-derived smooth muscle to TGF-beta1: Regulation of c-myb and alpha1 (I) procollagen genes. Exp. Cell Res. 1997, 230, 169–180. [Google Scholar] [CrossRef]
- Pfaltzgraff, E.R.; Shelton, E.L.; Galindo, C.L.; Nelms, B.L.; Hooper, C.W.; Poole, S.D.; Labosky, P.A.; Bader, D.M.; Reese, J. Embryonic domains of the aorta derived from diverse origins exhibit distinct properties that converge into a common phenotype in the adult. J. Mol. Cell Cardiol. 2014, 69, 88–96. [Google Scholar] [CrossRef]
- Simons, P.C.; Algra, A.; Bots, M.L.; Grobbee, D.E.; van der Graaf, Y. Common carotid intima-media thickness and arterial stiffness: Indicators of cardiovascular risk in high-risk patients. The SMART Study (Second Manifestations of ARTerial disease). Circulation 1999, 100, 951–957. [Google Scholar] [CrossRef] [PubMed]
- Arnett, D.K.; Evans, G.W.; Riley, W.A. Arterial stiffness: A new cardiovascular risk factor? Am. J. Epidemiol. 1994, 140, 669–682. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, Y.; Hamada, M.; Hiwada, K. Aortic distensibility is closely related to the progression of left ventricular hypertrophy in patients receiving hemodialysis. Angiology 2000, 51, 933–941. [Google Scholar] [CrossRef] [PubMed]
- Qin, F.; Siwik, D.A.; Luptak, I.; Hou, X.; Wang, L.; Higuchi, A.; Weisbrod, R.M.; Ouchi, N.; Tu, V.H.; Calamaras, T.D.; et al. The polyphenols resveratrol and S17834 prevent the structural and functional sequelae of diet-induced metabolic heart disease in mice. Circulation 2012, 125, 1757–1764. [Google Scholar] [CrossRef]
- Safar, M.E.; Levy, B.I.; London, G.M. Arterial structure in hypertension and the effects of angiotensin converting enzyme inhibition. J. Hypertens. Suppl. 1992, 10, S51–S57. [Google Scholar] [CrossRef]
- Bayer, I.M.; Adamson, S.L.; Langille, B.L. Atrophic remodeling of the artery-cuffed artery. Arterioscler. Thromb. Vasc. Biol. 1999, 19, 1499–1505. [Google Scholar] [CrossRef][Green Version]
- Bény, J.L.; Brunet, P.C. Electrophysiological and mechanical effects of substance P and acetylcholine on rabbit aorta. J. Physiol. 1988, 398, 277–289. [Google Scholar] [CrossRef]
- Brunner, F.; Kühberger, E.; Brockmeier, D.; Kukovetz, W.R. Evidence for muscarinic receptors in endothelial cells from combined functional and binding studies. Eur. J. Pharmacol. 1990, 187, 145–154. [Google Scholar] [CrossRef]
- Shetty, S.S.; Okada, T.; Webb, R.L.; DelGrande, D.; Lappe, R.W. Functionally distinct endothelin B receptors in vascular endothelium and smooth muscle. Biochem. Biophys. Res. Commun. 1993, 191, 459–464. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Adult | Aged | |||
|---|---|---|---|---|
| 0% Dill | 5% Dill | 10% Dill | ||
| Body weight (g) | 32.4 ± 1.5 | 30.4 ± 0.8 | 31.7 ± 0.8 | 32.5 ± 1 |
| HW/BW | 0.48 ± 0.01 | 0.56 ± 0.03 * | 0.47 ± 0.003 & | 0.48 ± 0.03 & |
| LV+S /BW | 0.37 ± 0.006 | 0.43 ± 0.009 * | 0.36 ± 0.005 & | 0.37 ± 0.02& |
| RV/BW | 0.1 ± 0.004 | 0.12 ± 0.006 | 0.1 ± 0.002 | 0.1 ± 0.007 |
| Adult | Aged | |||
|---|---|---|---|---|
| 0% Dill | 5% Dill | 10% Dill | ||
| Number of elastic lamellae (EL) | 6.81 ± 0.31 | 7.12 ± 0.36 | 7.58 ± 0.25 | 7.66 ± 0.33 |
| Number of disruptions per EL | 2.49 ± 0.11 | 5.93 ± 0.49 * | 4.58 ± 0.2 *& | 3.99 ± 0.46 *& |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fhayli, W.; Boëté, Q.; Kihal, N.; Cenizo, V.; Sommer, P.; Boyle, W.A.; Jacob, M.-P.; Faury, G. Dill Extract Induces Elastic Fiber Neosynthesis and Functional Improvement in the Ascending Aorta of Aged Mice with Reversal of Age-Dependent Cardiac Hypertrophy and Involvement of Lysyl Oxidase-Like-1. Biomolecules 2020, 10, 173. https://doi.org/10.3390/biom10020173
Fhayli W, Boëté Q, Kihal N, Cenizo V, Sommer P, Boyle WA, Jacob M-P, Faury G. Dill Extract Induces Elastic Fiber Neosynthesis and Functional Improvement in the Ascending Aorta of Aged Mice with Reversal of Age-Dependent Cardiac Hypertrophy and Involvement of Lysyl Oxidase-Like-1. Biomolecules. 2020; 10(2):173. https://doi.org/10.3390/biom10020173
Chicago/Turabian StyleFhayli, Wassim, Quentin Boëté, Nadjib Kihal, Valérie Cenizo, Pascal Sommer, Walter A. Boyle, Marie-Paule Jacob, and Gilles Faury. 2020. "Dill Extract Induces Elastic Fiber Neosynthesis and Functional Improvement in the Ascending Aorta of Aged Mice with Reversal of Age-Dependent Cardiac Hypertrophy and Involvement of Lysyl Oxidase-Like-1" Biomolecules 10, no. 2: 173. https://doi.org/10.3390/biom10020173
APA StyleFhayli, W., Boëté, Q., Kihal, N., Cenizo, V., Sommer, P., Boyle, W. A., Jacob, M.-P., & Faury, G. (2020). Dill Extract Induces Elastic Fiber Neosynthesis and Functional Improvement in the Ascending Aorta of Aged Mice with Reversal of Age-Dependent Cardiac Hypertrophy and Involvement of Lysyl Oxidase-Like-1. Biomolecules, 10(2), 173. https://doi.org/10.3390/biom10020173