Misincorporation of Galactose by Chondroitin Synthase of Escherichia coli K4: From Traces to Synthesis of Chondbiuronan, a Novel Chondroitin-Like Polysaccharide

Abstract
1. Introduction
2. Materials and Methods
2.1. Chemicals and Molecular Biology Supplies and Services
2.2. Bacterial Strains and Recombinant Vectors
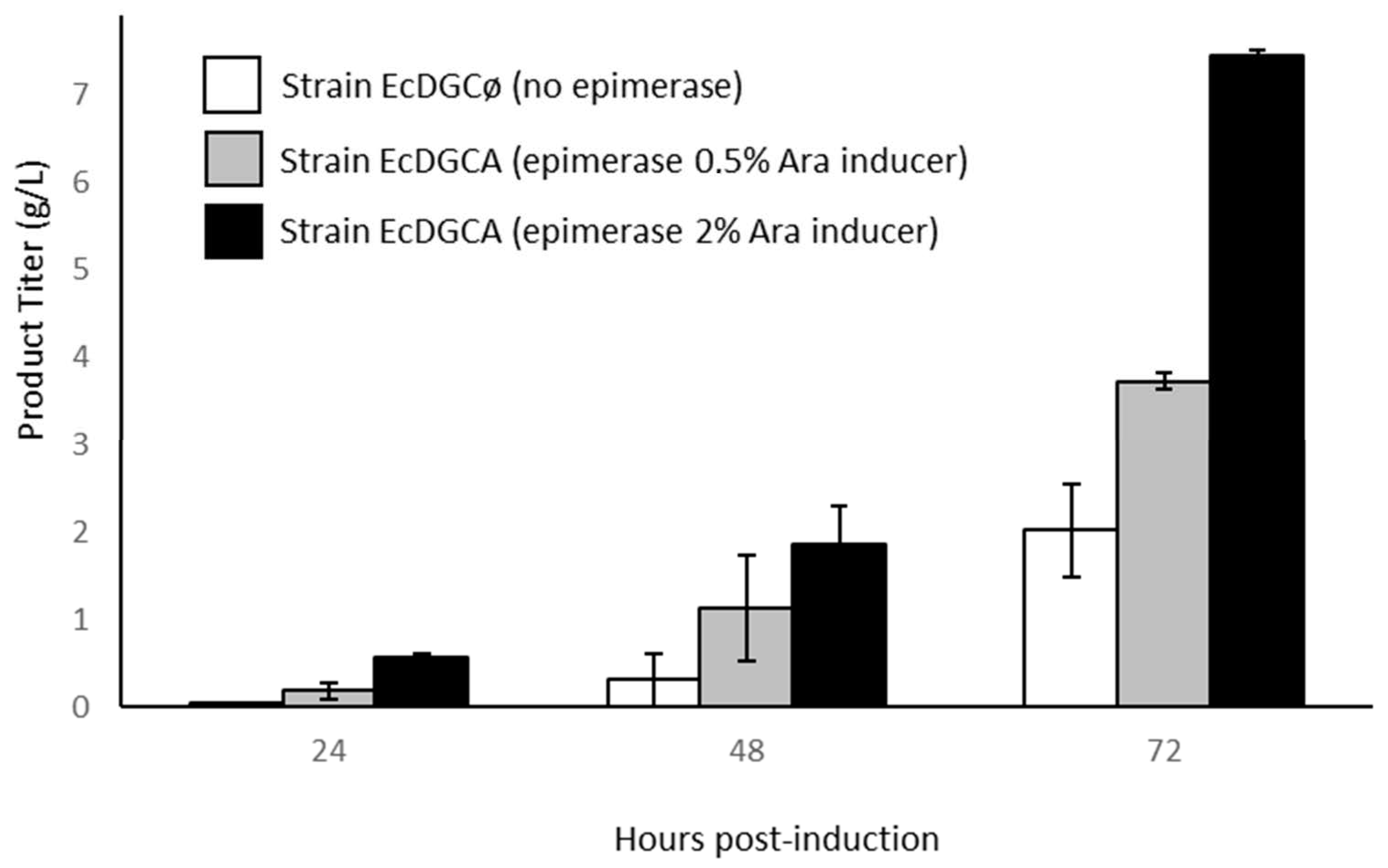
2.3. Culture Media and Culture Conditions for the Production of Polysaccharides
2.4. Extraction and Purification of Polysaccharides
2.5. Enzymatic Degradation
2.6. Carbohydrate Analyses
3. Results
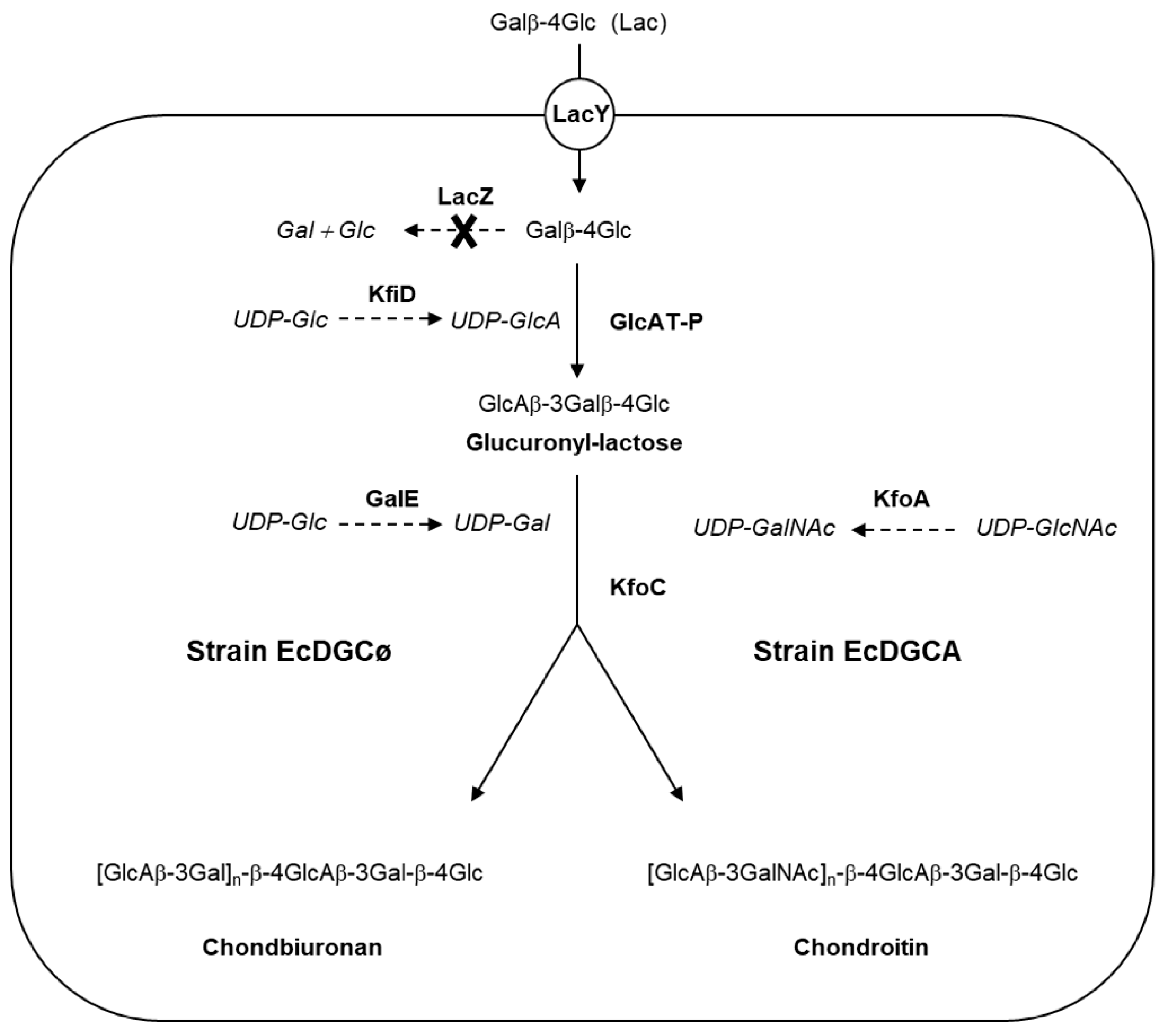
3.1. Engineering of E. Coli to Produce Chondroitin/Chondbiuronan Lactose
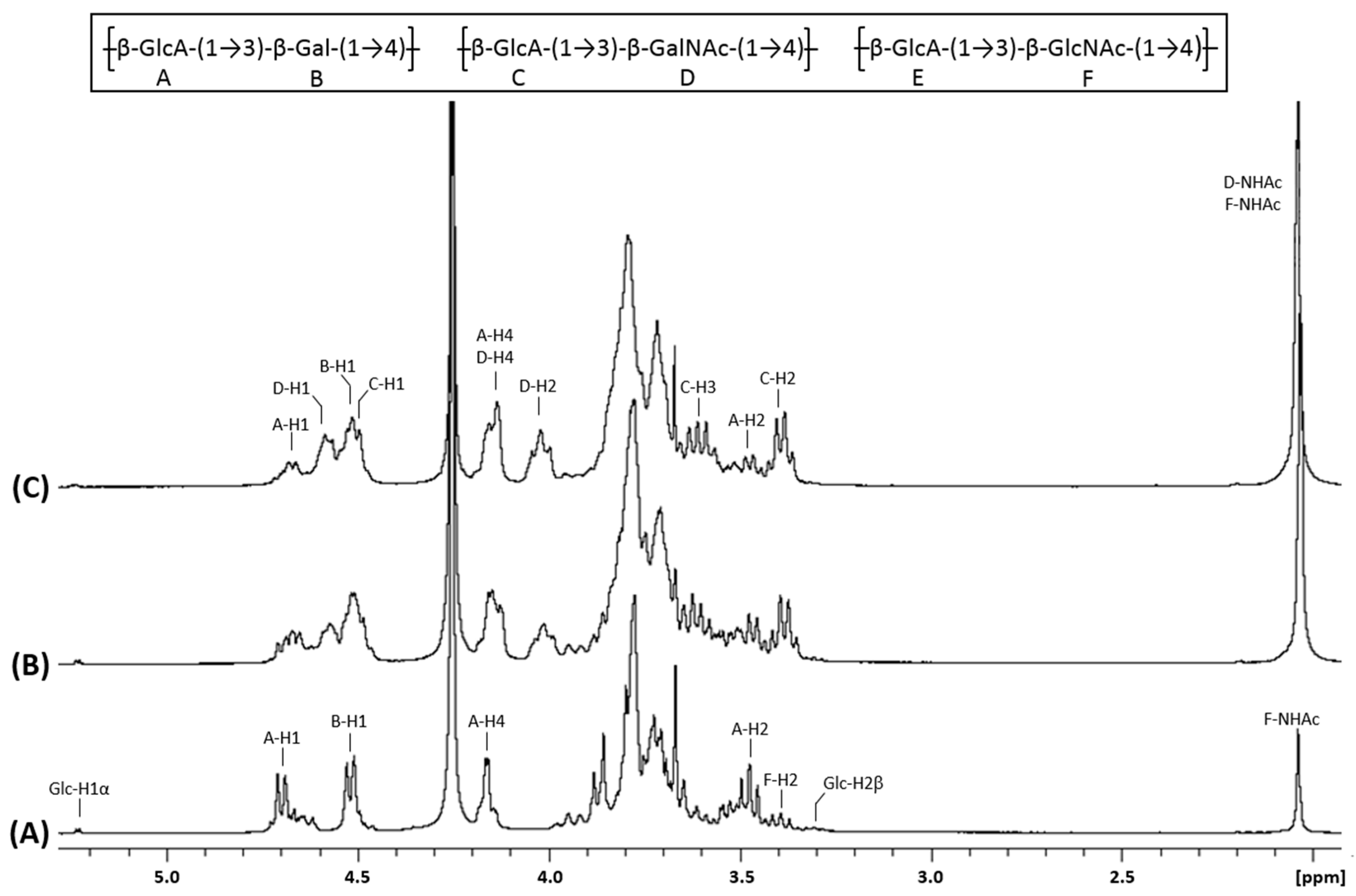
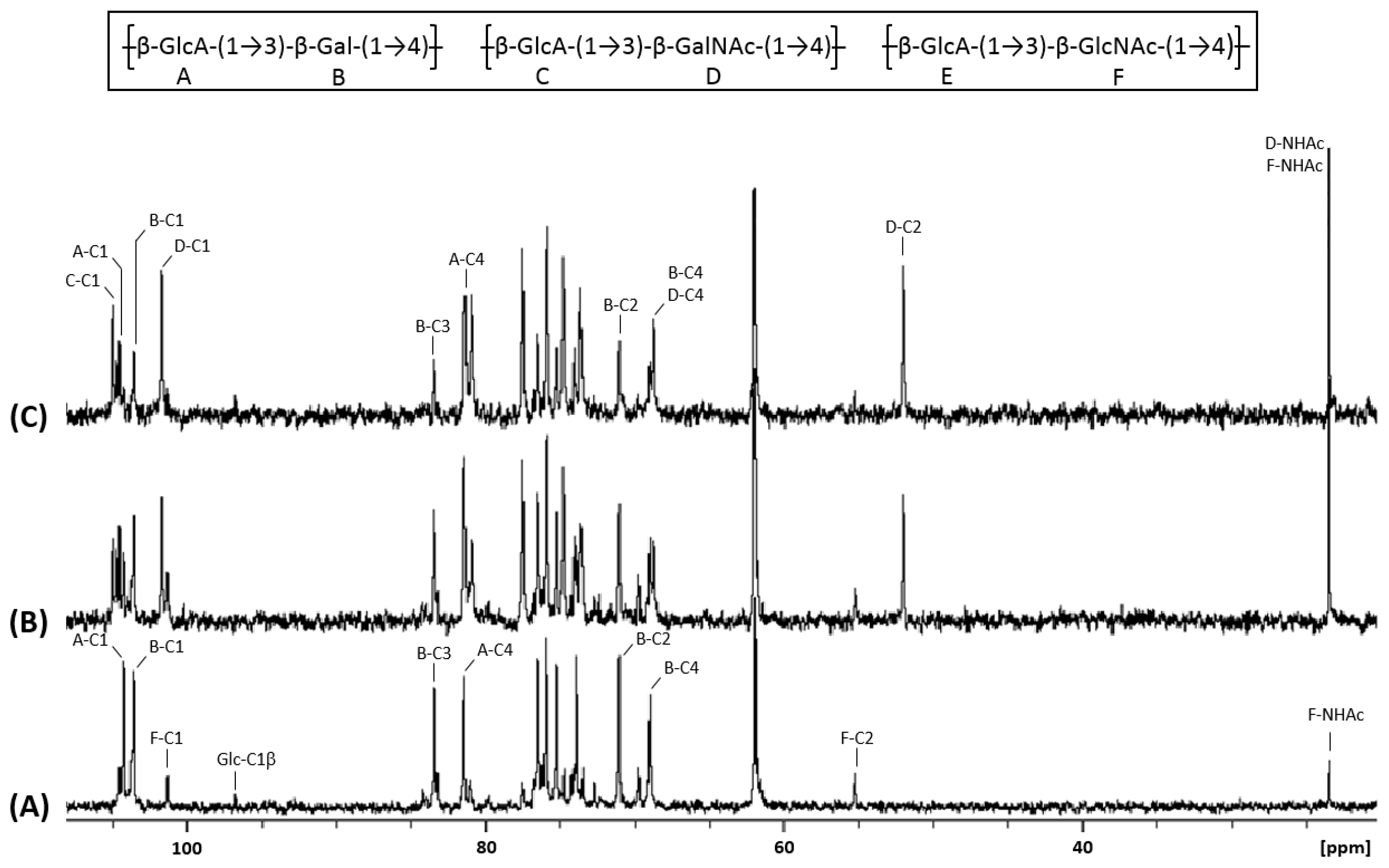
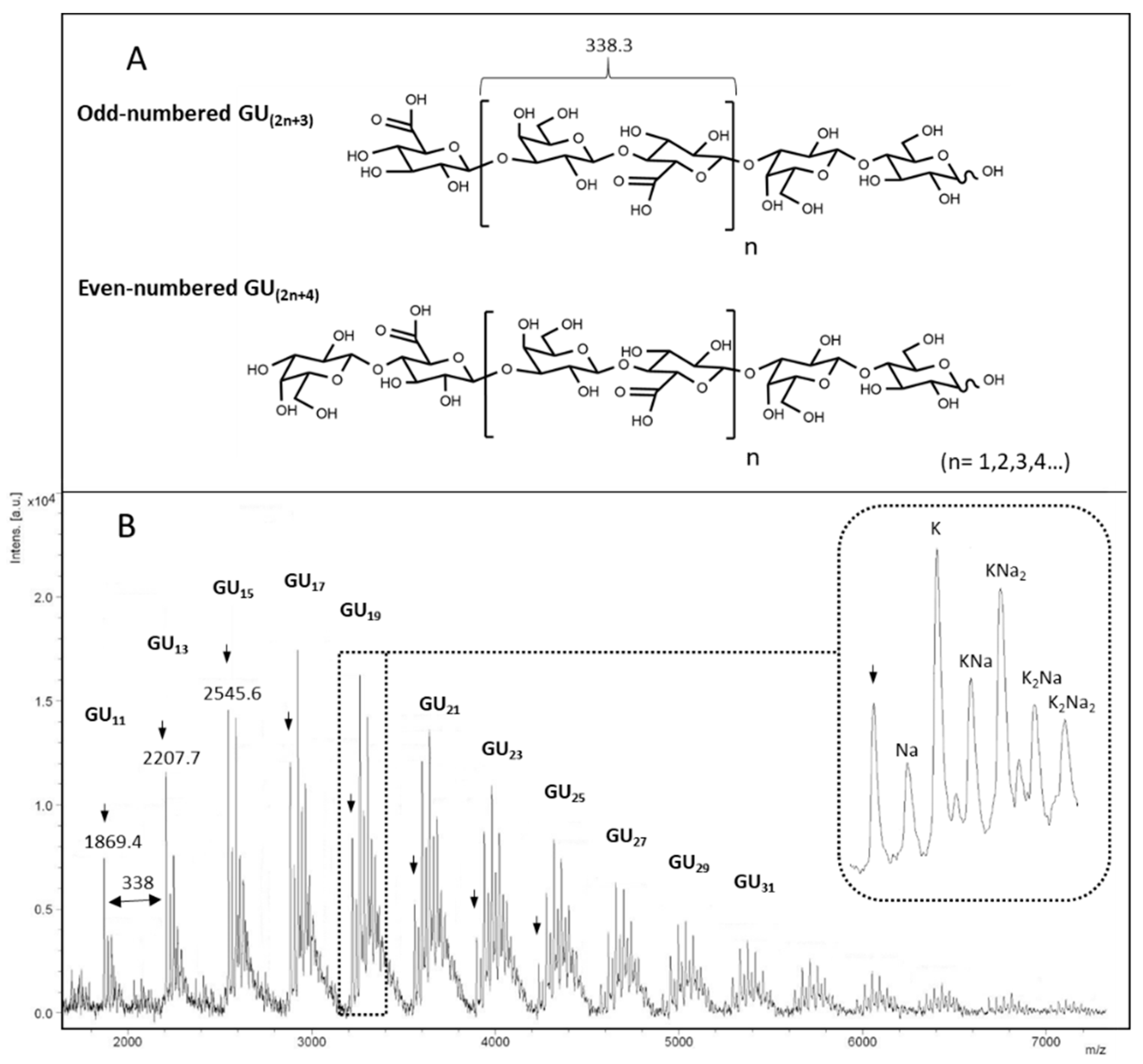
3.2. Structural Identification of Unknown Polymer Formed by KfoC in Absence of GalNAc
3.3. Galactose Incorporation Remains in Recombinant Chondroitin
3.4. N-Acetylglucosamine: Another Misincorporation of KfoC
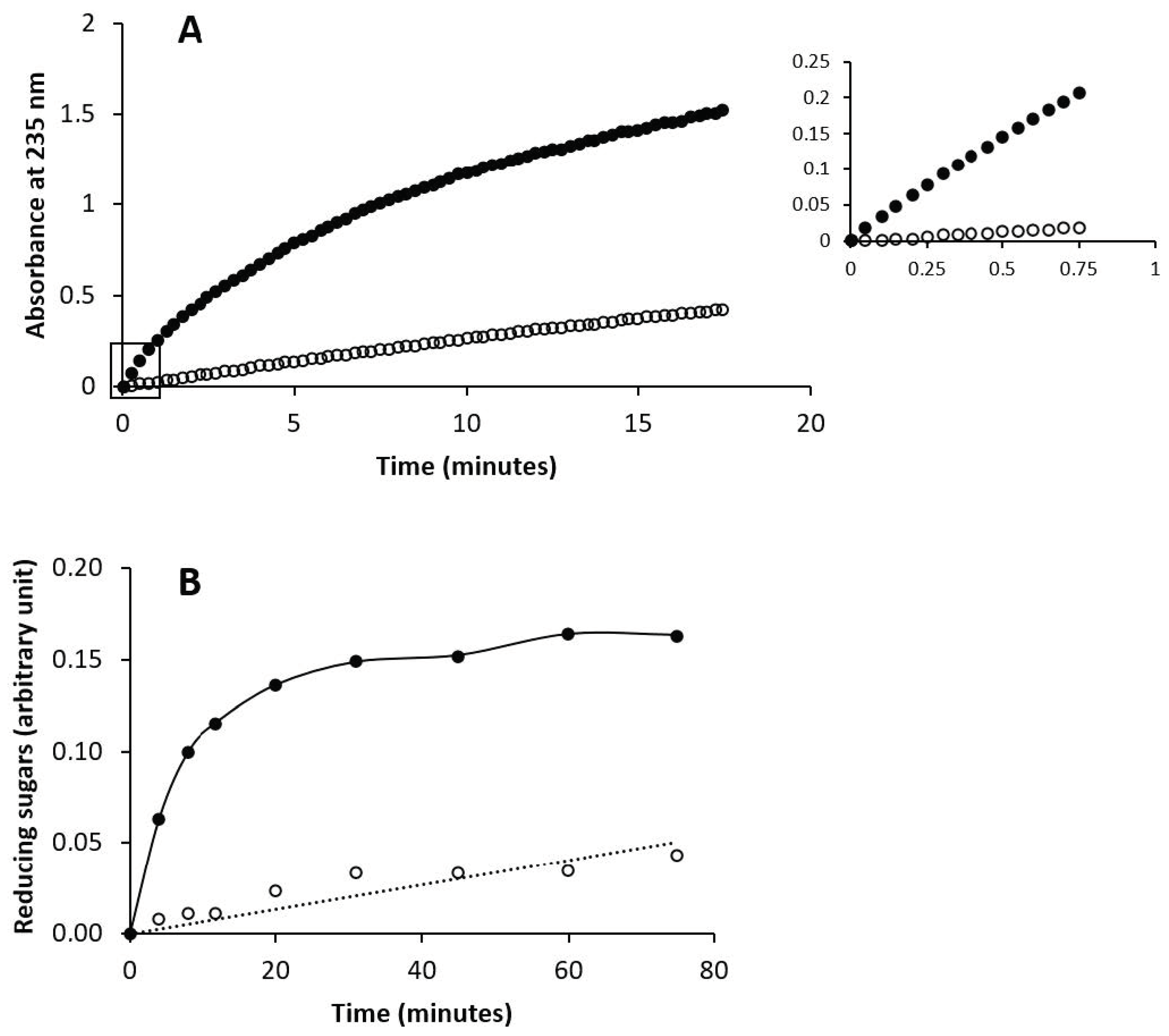
3.5. Enzymatic Susceptibility of Chondbiuronan
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Rodriguez, M.L.; Jann, B.; Jann, K. Structure and serological characteristics of the capsular K4 antigen of Escherichia coli 05:K4:H4, a fructose-containing polysaccharide with a chondroitin backbone. Eur. J. Biochem. 1988, 177, 117–124. [Google Scholar] [CrossRef]
- Rimler, R.B. Presumptive identification of Pasteurella multocida serogroup A, serogroup D and serogroup F by capsule depolymerization with mucopolysaccharidases. Vet. Rec. 1994, 134, 191–192. [Google Scholar] [CrossRef]
- Rimler, R.B.; Register, K.B.; Magyar, T.; Ackermann, M.R. Influence of chondroitinase on indirect hemagglutination titers and phagocytosis of Pasteurella multocida serogroups A, D and F. Vet. Microbiol. 1995, 47, 287–294. [Google Scholar] [CrossRef]
- Wu, J.-R.; Chen, P.-Y.; Shien, J.-H.; Shyu, C.-L.; Shieh, H.K.; Chang, F.; Chang, P.-C. Analysis of the biosynthesis genes and chemical components of the capsule of Avibacterium paragallinarum. Vet. Microbiol. 2010, 145, 90–99. [Google Scholar] [CrossRef]
- Truppe, W.; Kresse, H. Uptake of Proteoglycans and Sulfated Glycosaminoglycans by Cultured Skin Fibroblasts. JBIC J. Biol. Inorg. Chem. 1978, 85, 351–356. [Google Scholar] [CrossRef]
- Osawa, T.; Sugiura, N.; Shimada, H.; Hirooka, R.; Tsuji, A.; Shirakawa, T.; Fukuyama, K.; Kimura, M.; Kimata, K.; Kakuta, Y. Crystal structure of chondroitin polymerase from K4. Biochem. Biophys. Res. Commun. 2009, 378, 10–14. [Google Scholar] [CrossRef]
- Ninomiya, T. Molecular Cloning and Characterization of Chondroitin Polymerase from Escherichia coli strain K4. J. Biol. Chem. 2002, 277, 21567–21575. [Google Scholar] [CrossRef]
- Zanfardino, A.; Restaino, O.F.; Notomista, E.; Cimini, D.; Schiraldi, C.; De Rosa, M.; De Felice, M.; Varcamonti, M. Isolation of an Escherichia coli K4 kfoC mutant over-producing capsular chondroitin. Microb. Cell Factories 2010, 9, 34. [Google Scholar] [CrossRef]
- Jin, P.; Zhang, L.; Yuan, P.; Kang, Z.; Du, G.; Chen, J. Efficient biosynthesis of polysaccharides chondroitin and heparosan by metabolically engineered Bacillus subtilis. Carbohydr. Polym. 2016, 140, 424–432. [Google Scholar] [CrossRef]
- Sobhany, M.; Kakuta, Y.; Sugiura, N.; Kimata, K.; Negishi, M. The structural basis for a coordinated reaction catalysed by a bifunctional glycosyltransferase in chondroitin biosynthesis. J. Biol. Chem. 2012, 287, 36022–36028. [Google Scholar] [CrossRef]
- Sugiura, N.; Shimokata, S.; Minamisawa, T.; Hirabayashi, J.; Kimata, K.; Watanabe, H. Sequential synthesis of chondroitin oligosaccharides by immobilized chondroitin polymerase mutants. Glycoconj. J. 2008, 25, 521–530. [Google Scholar] [CrossRef]
- Xue, J.; Jin, L.; Zhang, X.; Wang, F.; Ling, P.; Sheng, J.-Z. Impact of donor binding on polymerization catalyzed by KfoC by regulating the affinity of enzyme for acceptor. Biochim. Biophys. Acta (BBA) Gen. Subj. 2016, 1860, 844–855. [Google Scholar] [CrossRef]
- Barreteau, H.; Richard, E.; Drouillard, S.; Samain, E.; Priem, B. Production of intracellular heparosan and derived oligosaccharides by lyase expression in metabolically engineered E. coli K-12. Carbohydr. Res. 2012, 360, 19–24. [Google Scholar] [CrossRef]
- Priem, B.; Peroux, J.; Colin-Morel, P.; Drouillard, S.; Fort, S. Chemo-bacterial synthesis of conjugatable glycosaminoglycans. Carbohydr. Polym. 2017, 167, 123–128. [Google Scholar] [CrossRef]
- Zhu, H.-M.; Sun, B.; Li, Y.-J.; Meng, D.-H.; Ju-Zheng, S.; Wang, T.-T.; Wang, F.; Sheng, J.-Z. KfoA, the UDP-glucose-4-epimerase of Escherichia coli strain O5:K4:H4, shows preference for acetylated substrates. Appl. Microbiol. Biotechnol. 2017, 102, 751–761. [Google Scholar] [CrossRef]
- Blumenkrantz, N.; Asboe-Hansen, G. New method for quantitative determination of uronic acids. Anal. Biochem. 1973, 54, 484–489. [Google Scholar] [CrossRef]
- Kidby, D.; Davidson, D. A convenient ferricyanide estimation of reducing sugars in the nanomole range. Anal. Biochem. 1973, 55, 321–325. [Google Scholar] [CrossRef]
- Stevenson, G.; Andrianopoulos, K.; Hobbs, M.; Reeves, P.R. Organization of the Escherichia coli K-12 gene cluster responsible for production of the extracellular polysaccharide colanic acid. J. Bacteriol. 1996, 178, 4885–4893. [Google Scholar] [CrossRef]
- Yavuz, E.; Drouillard, S.; Samain, E.; Roberts, I.; Priem, B. Glucuronylation in Escherichia coli for the bacterial synthesis of the carbohydrate moiety of nonsulfated HNK-1. Glycobiology 2007, 18, 152–157. [Google Scholar] [CrossRef][Green Version]
- Bastide, L.; Priem, B.; Fort, S. Chemo-bacterial synthesis and immunoreactivity of a brain HNK-1 analogue. Carbohydr. Res. 2011, 346, 348–351. [Google Scholar] [CrossRef]
- Studier, F.W.; Daegelen, P.; Lenski, R.E.; Maslov, S.; Kim, J.F. Understanding the differences between genome sequences of Escherichia coli B Strains REL606 and BL21(DE3) and comparison of the E. coli B and K-12 genomes. J. Mol. Biol. 2009, 394, 653–680. [Google Scholar] [CrossRef]
- Rye, C.S.; Withers, S.G. Elucidation of the Mechanism of Polysaccharide Cleavage by Chondroitin AC Lyase from Flavobacterium heparinum. J. Am. Chem. Soc. 2002, 124, 9756–9767. [Google Scholar] [CrossRef]
- Kakizaki, I.; Ibori, N.; Kojima, K.; Yamaguchi, M.; Endo, M. Mechanism for the hydrolysis of hyaluronan oligosaccharides by bovine testicular hyaluronidase. FEBS J. 2010, 277, 1776–1786. [Google Scholar] [CrossRef]
- Wolf, S.; Warnecke, S.; Ehrit, J.; Freiberger, F.; Gerardy-Schahn, R.; Meier, C. Chemical synthesis and enzymatic testing of CMP-sialic acid derivatives. ChemBioChem 2012, 13, 2605–2615. [Google Scholar] [CrossRef]
- Pouilly, S.; Bourgeaux, V.; Piller, F.; Piller, V. Evaluation of analogues of GalNAc as substrates for enzymes of the mammalian GalNAc salvage pathway. ACS Chem. Biol. 2012, 7, 753–760. [Google Scholar] [CrossRef]
- Lane, R.S.; Ange, K.S.; Zolghadr, B.; Liu, X.; Schäffer, C.; Linhardt, R.J.; DeAngelis, P.L. Expanding glycosaminoglycan chemical space: Towards the creation of sulfated analogs, novel polymers and chimeric constructs. Glycobiology 2017, 27, 646–656. [Google Scholar] [CrossRef]
- Gagnon, S.M.L.; Meloncelli, P.J.; Zheng, R.B.; Haji-Ghassemi, O.; Johal, A.R.; Borisova, S.N.; Lowary, T.L.; Evans, S.V. High resolution structures of the human ABO(H) blood group enzymes in complex with donor analogs reveal that the enzymes utilize multiple donor conformations to bind substrates in a stepwise manner. J. Biol. Chem. 2015, 290, 27040–27052. [Google Scholar] [CrossRef]
- Fort, S.; Birikaki, L.; Dubois, M.-P.; Antoine, T.; Samain, E.; Driguez, H. Biosynthesis of conjugatable saccharidic moieties of GM2 and GM3 gangliosides by engineered E. coli. Chem. Commun. 2005, 20, 2558–2560. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Culture | Mn | Mw | Mw/Mn |
|---|---|---|---|
| EcDGCø + IPTG | 6000 ± 1400 | 8000 ± 2000 | 1.34 ± 0.01 |
| EcDGCA + IPTG + 0.5% Ara | 13,700 ± 1400 | 24,000 ± 4800 | 1.79 ± 0.41 |
| EcDGCA + IPTG + 2% Ara | 18,600 ± 1800 | 30,400 ± 3000 | 1.63 ± 0.01 |
| Sugar Residue | 1 | 2 | 3 | 4 | 5 | 6 (6a,6b) | NHAc (CH3, CO) | |
|---|---|---|---|---|---|---|---|---|
| Chondbiuronan EcDGCø | ||||||||
| A→4)-β−D-GlcA-(1→ | 1H | 4.70 | 3.47 | 3.65 | 3.78 | 3.86 | no | |
| 13C | 104.24 | 73.91 | 75.25 | 81.43 | 76.51 | 175.78 | ||
| B→3)-β−D-Gal-(1→ | 1H | 4.52 | 3.71 | 3.79 | 4.16 | 3.72 | 3.79, 3.79 | |
| 13C | 103.57 | 71.05 | 83.42 | 68.97 | 75.95 | 61.94 | ||
| Chondroitin EcDGCA | ||||||||
| A→4)−β−D-GlcA-(1→ | 1H | 4.68 | 3.47 | 3.65 | 3.78 | 3.85 | no | |
| 13C | 104.53 | 74.01 | 75.25 | 81.43 | 76.53 | 175.78 | ||
| B→3)−β−D-Gal-(1→ | 1H | 4.52 | 3.71 | 3.79 | 4.16 | 3.72 | 3.79, 3.79 | |
| 13C | 103.57 | 71.03 | 83.45 | 68.99 | 75.95 | 61.94 | ||
| C→4)−β−D-GlcA-(1→ | 1H | 4.51 | 3.39 | 3.61 | 3.78 | 3.72 | no | |
| 13C | 104.99 | 73.65 | 74.80 | 80.93 | 77.52 | 175.11 | ||
| D→3)-β-D-GalNAc-(1→ | 1H | 4.58 | 4.02 | 3.82 | 4.14 | 3.72 | 3.69, 3.94 | 2.04 |
| 13C | 101.71 | 52.01 | 81.36 | 68.76 | 75.88 | 62.00 | 23.47, 175.75 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Leroux, M.; Michaud, J.; Bayma, E.; Armand, S.; Drouillard, S.; Priem, B. Misincorporation of Galactose by Chondroitin Synthase of Escherichia coli K4: From Traces to Synthesis of Chondbiuronan, a Novel Chondroitin-Like Polysaccharide. Biomolecules 2020, 10, 1667. https://doi.org/10.3390/biom10121667
Leroux M, Michaud J, Bayma E, Armand S, Drouillard S, Priem B. Misincorporation of Galactose by Chondroitin Synthase of Escherichia coli K4: From Traces to Synthesis of Chondbiuronan, a Novel Chondroitin-Like Polysaccharide. Biomolecules. 2020; 10(12):1667. https://doi.org/10.3390/biom10121667
Chicago/Turabian StyleLeroux, Mélanie, Julie Michaud, Eric Bayma, Sylvie Armand, Sophie Drouillard, and Bernard Priem. 2020. "Misincorporation of Galactose by Chondroitin Synthase of Escherichia coli K4: From Traces to Synthesis of Chondbiuronan, a Novel Chondroitin-Like Polysaccharide" Biomolecules 10, no. 12: 1667. https://doi.org/10.3390/biom10121667
APA StyleLeroux, M., Michaud, J., Bayma, E., Armand, S., Drouillard, S., & Priem, B. (2020). Misincorporation of Galactose by Chondroitin Synthase of Escherichia coli K4: From Traces to Synthesis of Chondbiuronan, a Novel Chondroitin-Like Polysaccharide. Biomolecules, 10(12), 1667. https://doi.org/10.3390/biom10121667