Identification and Validation of Carbonic Anhydrase II as the First Target of the Anti-Inflammatory Drug Actarit




{kind=link}
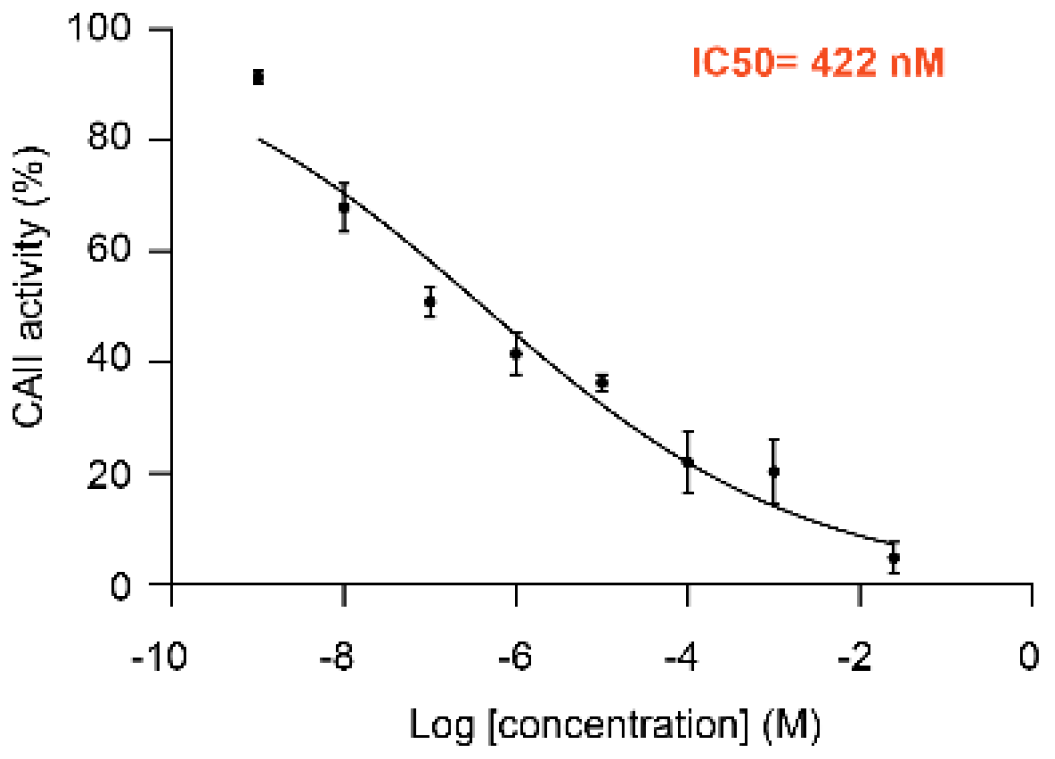{kind=link}
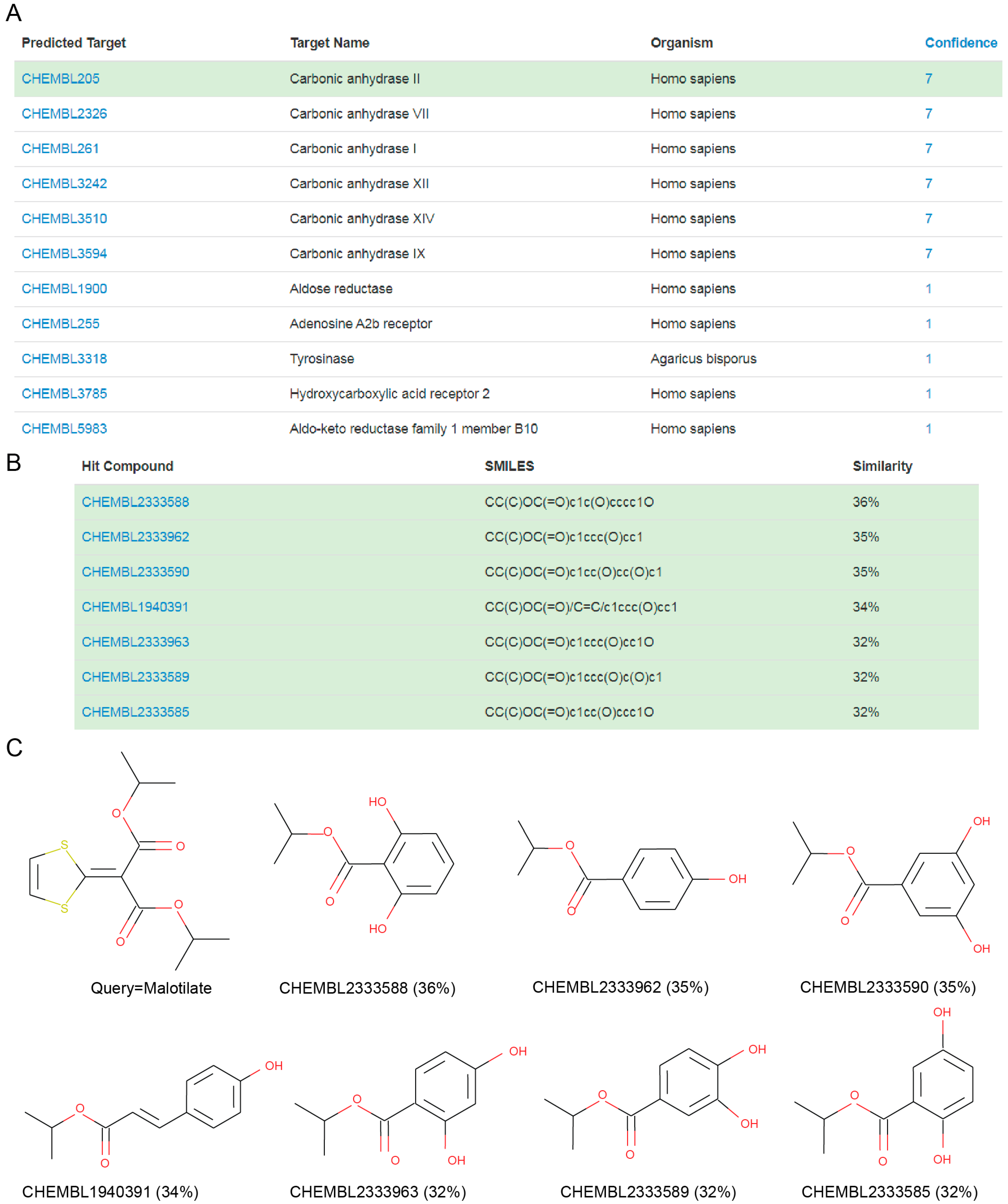{kind=link}
{kind=link}
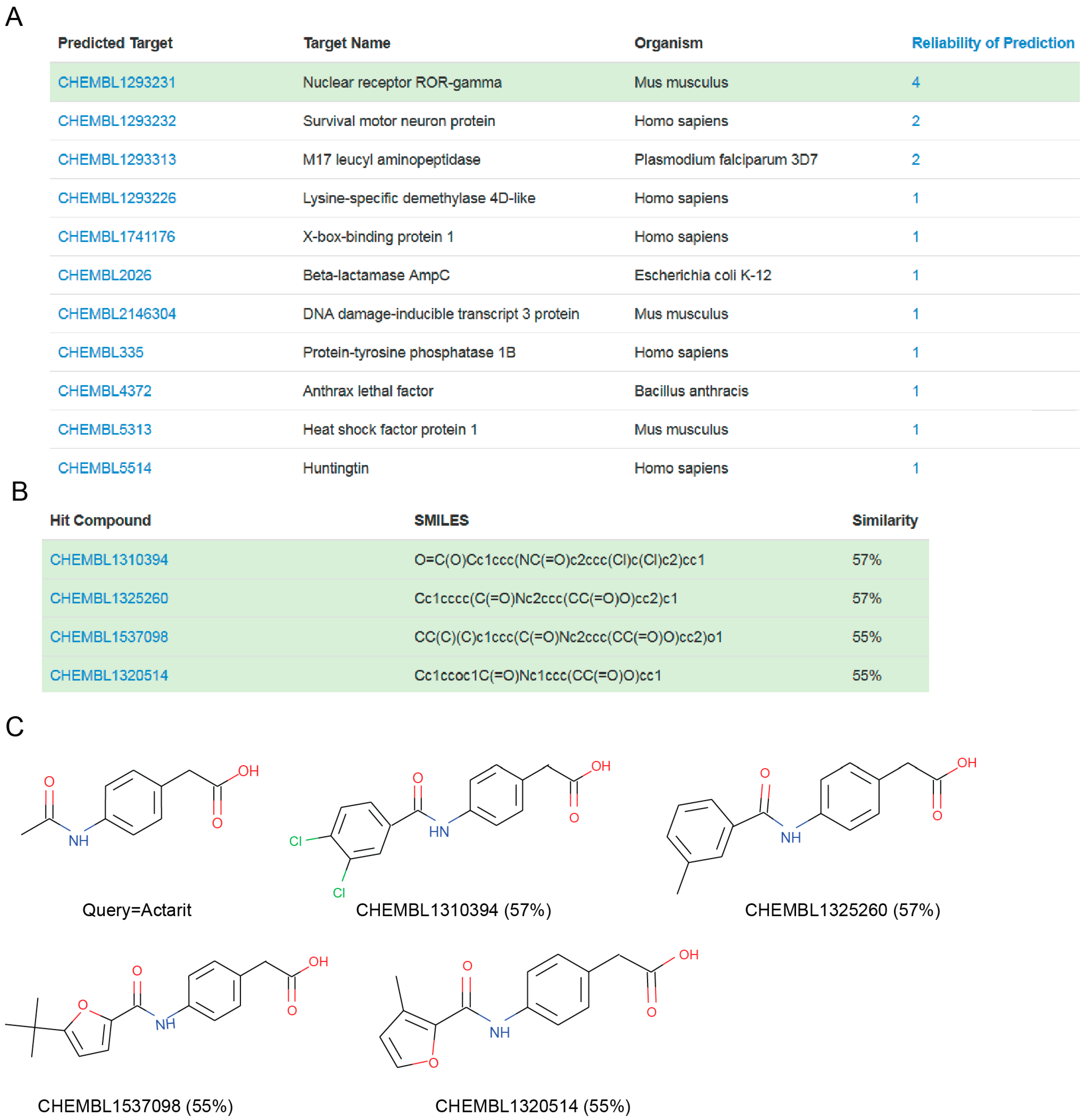{kind=link}
Abstract
- What Is Already Known:
- Computational target prediction methods complement and/or guide experimental approaches to characterise the polypharmacology of drugs.
- Ligand-centric chemical-similarity-based methods (e.g., MolTarPred, freely available at http://moltarpred.marseille.inserm.fr/) have been found to be particularly powerful.
- What This Study Adds:
- Experimental confirmation of MolTarPred prediction of CAII as a target of Actarit with a mid-nanomolar IC50.
- A proof-of-concept of MolTarPred’s utility for target-orphan drugs, providing other plausible target predictions for actarit (e.g., RORγ).
- Clinical Significance:
- The CAII-actarit association sheds light into its mechanism of action as a drug for RA (rheumatoid arthritis).
- Repositioning of actarit can now be investigated for other CAII-linked indications.
1. Introduction
2. Materials and Methods
2.1. MolTarPred (Molecular Target Prediction)
2.2. CAII Activity Assay
2.3. Materials
3. Results
4. Discussion and Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| CAII | Carbonic anhydrase II |
| FDA | Food and Drug Administration |
| MolTarPred | Molecular Target Prediction |
| RA | Rheumatoid Arthritis |
| RORγ | RAR-related orphan receptor gamma |
| SEA | Similarity Ensemble Approach |
References
- Li, Y.Y.; Jones, S.J.M. Drug repositioning for personalized medicine. Genome Med. 2012, 4, 27. [Google Scholar] [CrossRef]
- Lysenko, A.; Sharma, A.; Boroevich, K.A.; Tsunoda, T. An integrative machine learning approach for prediction of toxicity-related drug safety. Life Sci. Alliance 2018, 1. [Google Scholar] [CrossRef]
- Mathai, N.; Chen, Y.; Kirchmair, J. Validation strategies for target prediction methods. Brief. Bioinform. 2020, 21, 791–802. [Google Scholar] [CrossRef]
- Peón, A.; Naulaerts, S.; Ballester, P.J. Predicting the Reliability of Drug-target Interaction Predictions with Maximum Coverage of Target Space. Sci. Rep. 2017, 7, 3820. [Google Scholar] [CrossRef]
- Peón, A.; Dang, C.C.; Ballester, P.J. How Reliable Are Ligand-Centric Methods for Target Fishing? Front. Chem. 2016, 4, 15. [Google Scholar] [CrossRef]
- Mathai, N.; Kirchmair, J. Similarity-Based Methods and Machine Learning Approaches for Target Prediction in Early Drug Discovery: Performance and Scope. Int. J. Mol. Sci. 2020, 21, 3585. [Google Scholar] [CrossRef]
- Gregori-Puigjané, E.; Setola, V.; Hert, J.; Crews, B.A.; Irwin, J.J.; Lounkine, E.; Marnett, L.; Roth, B.L.; Shoichet, B.K. Identifying mechanism-of-action targets for drugs and probes. Proc. Natl. Acad. Sci. USA 2012, 109, 11178–11183. [Google Scholar] [CrossRef]
- Mugumbate, G.; Mendes, V.; Blaszczyk, M.; Sabbah, M.; Papadatos, G.; Lelievre, J.; Ballell, L.; Barros, D.; Abell, C.; Blundell, T.L.; et al. Target Identification of Mycobacterium tuberculosis Phenotypic Hits Using a Concerted Chemogenomic, Biophysical, and Structural Approach. Front. Pharmacol. 2017, 8, 681. [Google Scholar] [CrossRef]
- Ariey-Bonnet, J.; Carrasco, K.; Le Grand, M.; Hoffer, L.; Betzi, S.; Feracci, M.; Tsvetkov, P.; Devred, F.; Collette, Y.; Morelli, X.; et al. In silico molecular target prediction unveils mebendazole as a potent MAPK14 inhibitor. Mol. Oncol. 2020. [Google Scholar] [CrossRef]
- Peón, A.; Li, H.; Ghislat, G.; Leung, K.S.; Wong, M.H.; Lu, G.; Ballester, P.J. MolTarPred: A web tool for comprehensive target prediction with reliability estimation. Chem. Biol. Drug Des. 2019, 94, 1390–1401. [Google Scholar] [CrossRef]
- Bento, A.P.; Gaulton, A.; Hersey, A.; Bellis, L.J.; Chambers, J.; Davies, M.; Krüger, F.A.; Light, Y.; Mak, L.; McGlinchey, S.; et al. The ChEMBL bioactivity database: An update. Nucleic Acids Res. 2014, 42, D1083–D1090. [Google Scholar] [CrossRef]
- Anderson, N.G.; Wilbur, K.M. Electrometric and colorimetric determination of carbonic anhydrase. Anat. Rec. 1948, 101, 685. [Google Scholar]
- Aso, M.; Tanaka, Y.; Saito, K.; Fujii, K.; Takazawa, A.; Ota, T.; Eto, S. Additive combination of actarit and methotrexate in the treatment of refractory rheumatoid arthritis. Mod. Rheumatol. 2000, 10, 103–109. [Google Scholar] [CrossRef]
- Benjamin, O.; Bansal, P.; Goyal, A.; Lappin, S. Disease Modifying Anti-Rheumatic Drugs (DMARD); StatPearls Publishing: Treasure Island, FL, USA, 2019. [Google Scholar]
- Nagaoka, S.; Ohno, M.; Ishigatsubo, Y. Usefulness of actarit in elderly rheumatoid arthritis. Mod. Rheumatol. 2002, 12, 118–127. [Google Scholar] [CrossRef]
- Zhang, J.; Gao, Y.; Qian, S.; Liu, X.; Zu, H. Physicochemical and pharmacokinetic characterization of a spray-dried malotilate emulsion. Int. J. Pharm. 2011, 414, 186–192. [Google Scholar] [CrossRef]
- Nakamura, H.; Ueki, Y.; Sakito, S.; Matsumoto, K.; Yano, M.; Miyake, S.; Tominaga, Y.; Eguchi, K. Clinical effects of actarit in rheumatoid arthritis: Improvement of early disease activity mediated by reduction of serum concentrations of nitric oxide. Clin. Exp. Rheumatol. 2000, 18, 445–450. [Google Scholar]
- Li, Y.; Pan, H.; Duan, H.; Chen, J.; Zhu, Z.; Fan, J.; Li, P.; Yang, X.; Pan, W. Double-layered osmotic pump controlled release tablets of actarit: In vitro and in vivo evaluation. Asian J. Pharm. Sci. 2019, 14, 340–348. [Google Scholar] [CrossRef]
- Ye, J.; Wang, Q.; Zhou, X.; Zhang, N. Injectable actarit-loaded solid lipid nanoparticles as passive targeting therapeutic agents for rheumatoid arthritis. Int. J. Pharm. 2008, 352, 273–279. [Google Scholar] [CrossRef]
- Abiramasundari, A.; Joshi, R.P.; Jalani, H.B.; Sharma, J.A.; Pandya, D.H.; Pandya, A.N.; Sudarsanam, V.; Vasu, K.K. Stability-indicating assay method for determination of actarit, its process related impurities and degradation products: Insight into stability profile and degradation pathways. J. Pharm. Anal. 2014, 4, 374–383. [Google Scholar] [CrossRef][Green Version]
- Canudas, N.; Figueroa, L.; Zamora, D.; Villamizar, J.E.; Guedez, A.; Fuentes, J.M.; Castelli, C.; Taddei, A. Photosensitizing properties of actarit, an antirheumatic drug. Arzneim. Forsch. Drug Res. 2008, 58, 182–187. [Google Scholar] [CrossRef]
- Bosc, N.; Atkinson, F.; Felix, E.; Gaulton, A.; Hersey, A.; Leach, A.R. Large scale comparison of QSAR and conformal prediction methods and their applications in drug discovery. J. Cheminform. 2019, 11, 4. [Google Scholar] [CrossRef] [PubMed]
- Bua, S.; Di Cesare Mannelli, L.; Vullo, D.; Ghelardini, C.; Bartolucci, G.; Scozzafava, A.; Supuran, C.T.; Carta, F. Design and Synthesis of Novel Nonsteroidal Anti-Inflammatory Drugs and Carbonic Anhydrase Inhibitors Hybrids (NSAIDs-CAIs) for the Treatment of Rheumatoid Arthritis. J. Med. Chem. 2017, 60, 1159–1170. [Google Scholar] [CrossRef] [PubMed]
- Berrino, E.; Milazzo, L.; Micheli, L.; Vullo, D.; Angeli, A.; Bozdag, M.; Nocentini, A.; Menicatti, M.; Bartolucci, G.; Di Cesare Mannelli, L.; et al. Synthesis and evaluation of carbonic anhydrase inhibitors with carbon monoxide releasing properties for the management of rheumatoid arthritis. J. Med. Chem. 2019, 62, 7233–7249. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Wei, Y.; Wang, J.; Pi, L.; Huang, J.; Wang, P. Carbonic anhydrases III and IV autoantibodies in rheumatoid arthritis, systemic lupus erythematosus, diabetes, hypertensive renal disease, and heart failure. Clin. Dev. Immunol. 2012, 2012, 354594. [Google Scholar] [CrossRef] [PubMed]
- Araujo, G.R.; Vaz, E.R.; Fujimura, P.T.; Fonseca, J.E.; de Lima, L.M.; Canhão, H.; Venturini, G.; Cardozo, K.H.M.; Carvalho, V.M.; Napimoga, M.H.; et al. Improved serological detection of rheumatoid arthritis: A highly antigenic mimotope of carbonic anhydrase III selected in a murine model by phage display. Arthritis Res. Ther. 2015, 17, 168. [Google Scholar] [CrossRef]
- Alver, A.; Şentürk, A.; Çakirbay, H.; Menteşe, A.; Gökmen, F.; Keha, E.E.; Uçar, F. Carbonic anhydrase II autoantibody and oxidative stress in rheumatoid arthritis. Clin. Biochem. 2011, 44, 1385–1389. [Google Scholar] [CrossRef] [PubMed]
- Supuran, C.T. Structure and function of carbonic anhydrases. Biochem. J. 2016, 473, 2023–2032. [Google Scholar] [CrossRef]
- Kuo, W.H.; Yang, S.F.; Hsieh, Y.S.; Tsai, C.S.; Hwang, W.L.; Chu, S.C. Differential expression of carbonic anhydrase isoenzymes in various types of anemia. Clin. Chim. Acta 2005, 351, 79–86. [Google Scholar] [CrossRef]
- McMahon, C.; Will, A.; Hu, P.; Shah, G.N.; Sly, W.S.; Smith, O.P. Bone marrow transplantation corrects osteopetrosis in the carbonic anhydrase II deficiency syndrome. Blood 2001, 97, 1947–1950. [Google Scholar] [CrossRef]
- Scozzafava, A.; Supuran, C.T. Glaucoma and the applications of carbonic anhydrase inhibitors. Subcell. Biochem. 2014, 75, 349–359. [Google Scholar]
- Innocenti, A.; Hilvo, M.; Scozzafava, A.; Parkkila, S.; Supuran, C.T. Carbonic anhydrase inhibitors: Inhibition of the new membrane-associated isoform XV with phenols. Bioorganic Med. Chem. Lett. 2008, 18, 3593–3596. [Google Scholar] [CrossRef] [PubMed]
- Zijlstra, F.J.; Paul Wilson, J.H.; Vermeer, M.A.; Ouwendijk, R.J.T.; Eric Vincent, J. Differential effects of malotilate on 5-, 12- and 15-lipoxygenase in human ascites cells. Eur. J. Pharmacol. 1989, 159, 291–295. [Google Scholar] [CrossRef]
- Willett, P. The Calculation of Molecular Structural Similarity: Principles and Practice. Mol. Inform. 2014, 33, 403–413. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Liu, Y.; Alvarez, B.V.; Casey, J.R.; Fliegel, L. A novel carbonic anhydrase II binding site regulates NHE1 activity. Biochemistry 2006, 45, 2414–2424. [Google Scholar] [CrossRef] [PubMed]
- Bronner, S.M.; Zbieg, J.R.; Crawford, J.J. RORγ antagonists and inverse agonists: A patent review. Expert Opin. Ther. Pat. 2017, 27, 101–112. [Google Scholar] [CrossRef]
- Capone, A.; Volpe, E. Transcriptional Regulators of T Helper 17 Cell Differentiation in Health and Autoimmune Diseases. Front. Immunol. 2020, 11, 348. [Google Scholar] [CrossRef]
- Maddur, M.S.; Miossec, P.; Kaveri, S.V.; Bayry, J. Th17 cells: Biology, pathogenesis of autoimmune and inflammatory diseases, and therapeutic strategies. Am. J. Pathol. 2012, 181, 8–18. [Google Scholar] [CrossRef]
- Zhang, Y.; Wallace, D.L.; De Lara, C.M.; Ghattas, H.; Asquith, B.; Worth, A.; Griffin, G.E.; Taylor, G.P.; Tough, D.F.; Beverley, P.C.L.; et al. In vivo kinetics of human natural killer cells: The effects of ageing and acute and chronic viral infection. Immunology 2007, 121, 258–265. [Google Scholar] [CrossRef]
- Kumar, N.; Lyda, B.; Chang, M.R.; Lauer, J.L.; Solt, L.A.; Burris, T.P.; Kamenecka, T.M.; Griffin, P.R. Identification of SR2211: A potent synthetic RORγ-selective modulator. ACS Chem. Biol. 2012, 7, 672–677. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ghislat, G.; Rahman, T.; Ballester, P.J. Identification and Validation of Carbonic Anhydrase II as the First Target of the Anti-Inflammatory Drug Actarit. Biomolecules 2020, 10, 1570. https://doi.org/10.3390/biom10111570
Ghislat G, Rahman T, Ballester PJ. Identification and Validation of Carbonic Anhydrase II as the First Target of the Anti-Inflammatory Drug Actarit. Biomolecules. 2020; 10(11):1570. https://doi.org/10.3390/biom10111570
Chicago/Turabian StyleGhislat, Ghita, Taufiq Rahman, and Pedro J. Ballester. 2020. "Identification and Validation of Carbonic Anhydrase II as the First Target of the Anti-Inflammatory Drug Actarit" Biomolecules 10, no. 11: 1570. https://doi.org/10.3390/biom10111570
APA StyleGhislat, G., Rahman, T., & Ballester, P. J. (2020). Identification and Validation of Carbonic Anhydrase II as the First Target of the Anti-Inflammatory Drug Actarit. Biomolecules, 10(11), 1570. https://doi.org/10.3390/biom10111570