Generation and Application of the Zebrafish heg1 Mutant as a Cardiovascular Disease Model


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Zebrafish Care and Maintenance
2.2. Purification of Hearts from Zebrafish Embryos
2.3. Generation of heg1 Mutant Zebrafish
2.4. Cardiac Phenotype Analysis
2.5. Blood Flow Rate and Heart Rate Statistics
2.6. Real-Time Quantitative PCR (qRT-PCR) Analysis
2.7. Whole-Mount in situ Hybridization and Red Blood Cell Staining Assay
2.8. Drug Preparation
2.9. Assessment Effects of Drugs on heg1 Mutant
2.10. Statistical Analysis
3. Results
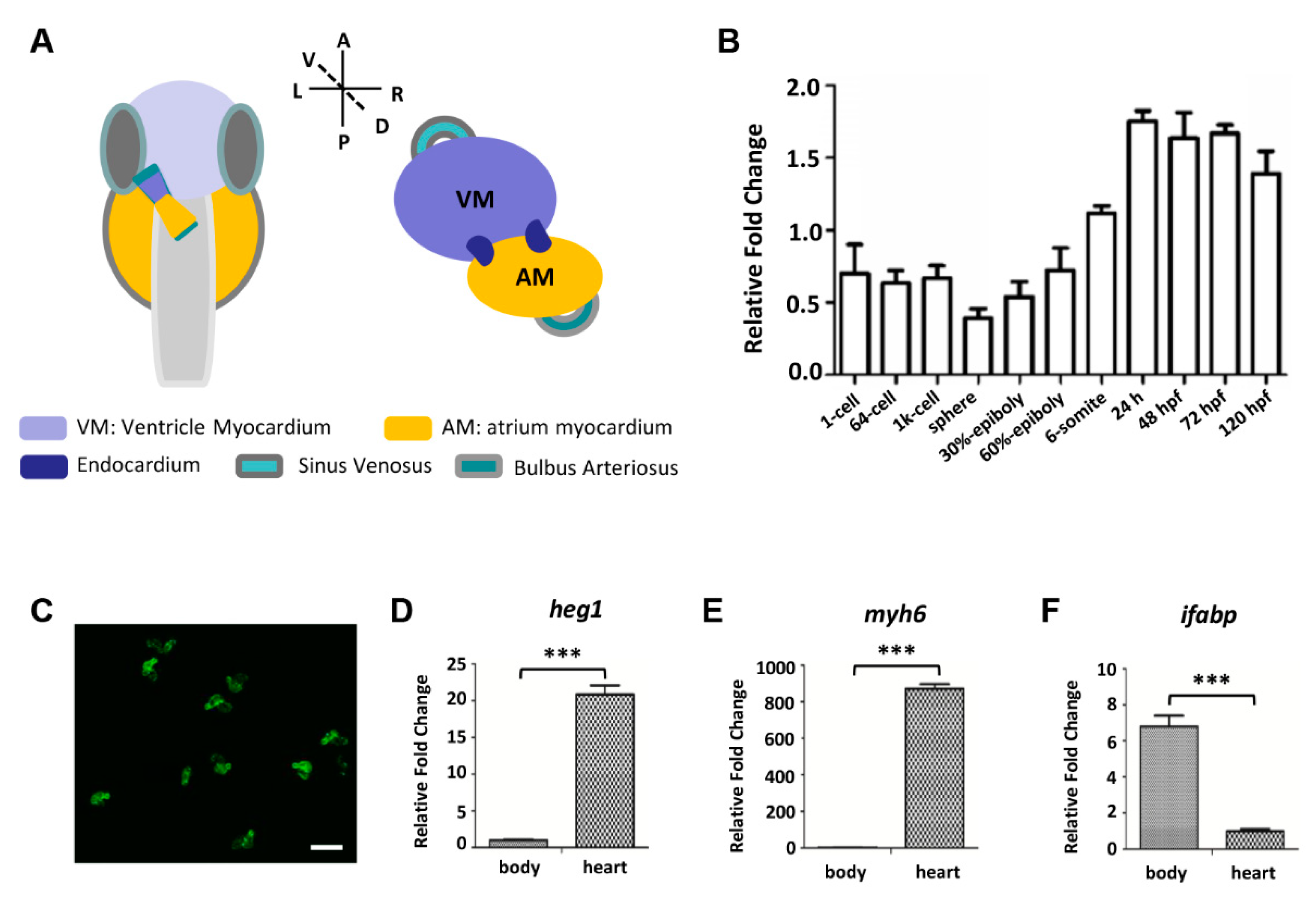
3.1. Zebrafish heg1 was Highly Expressed in Cardiac Tissue
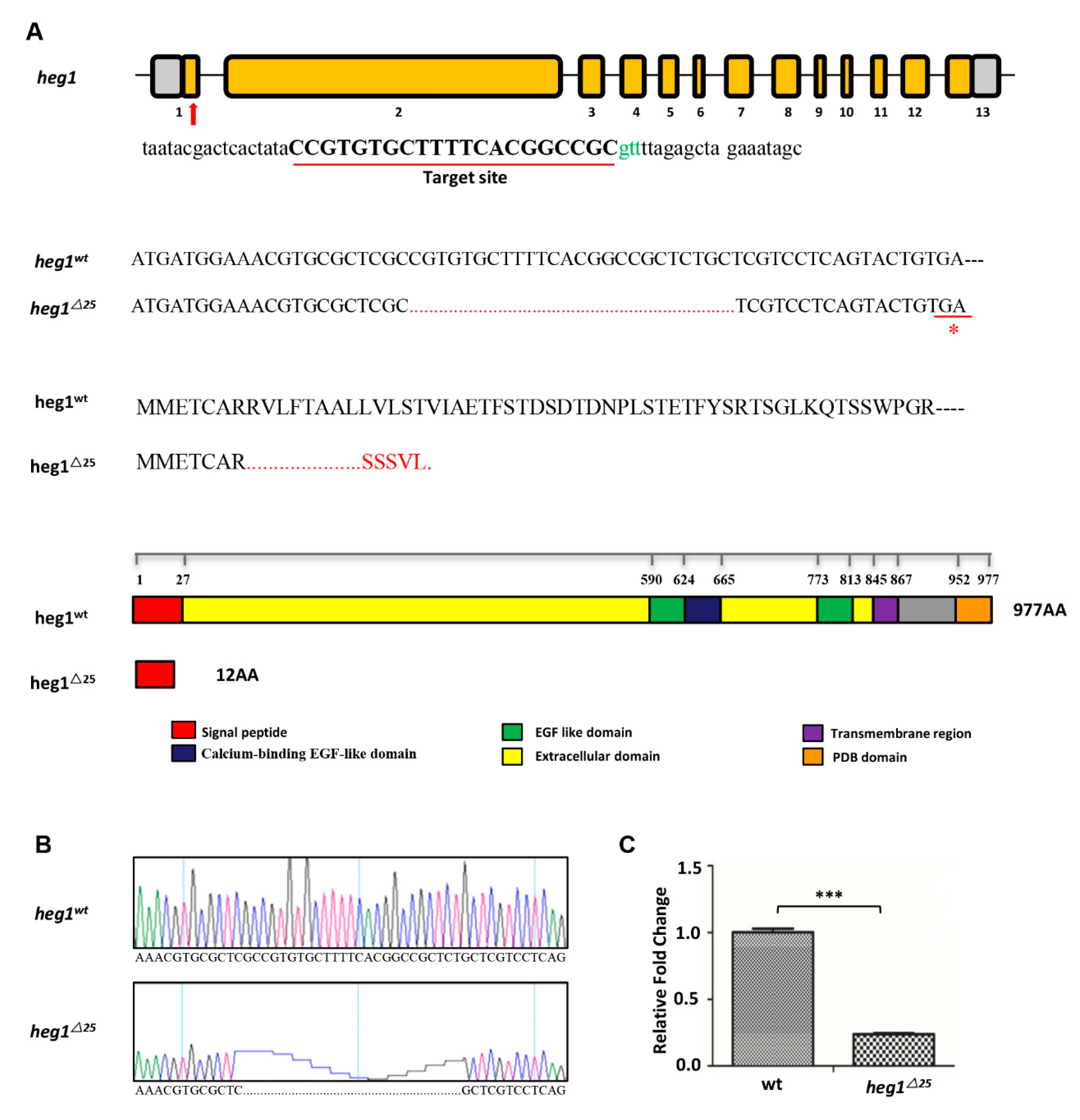
3.2. heg1 Deficient Zebrafish Was Generated Using CRISPR/Cas9 Technology
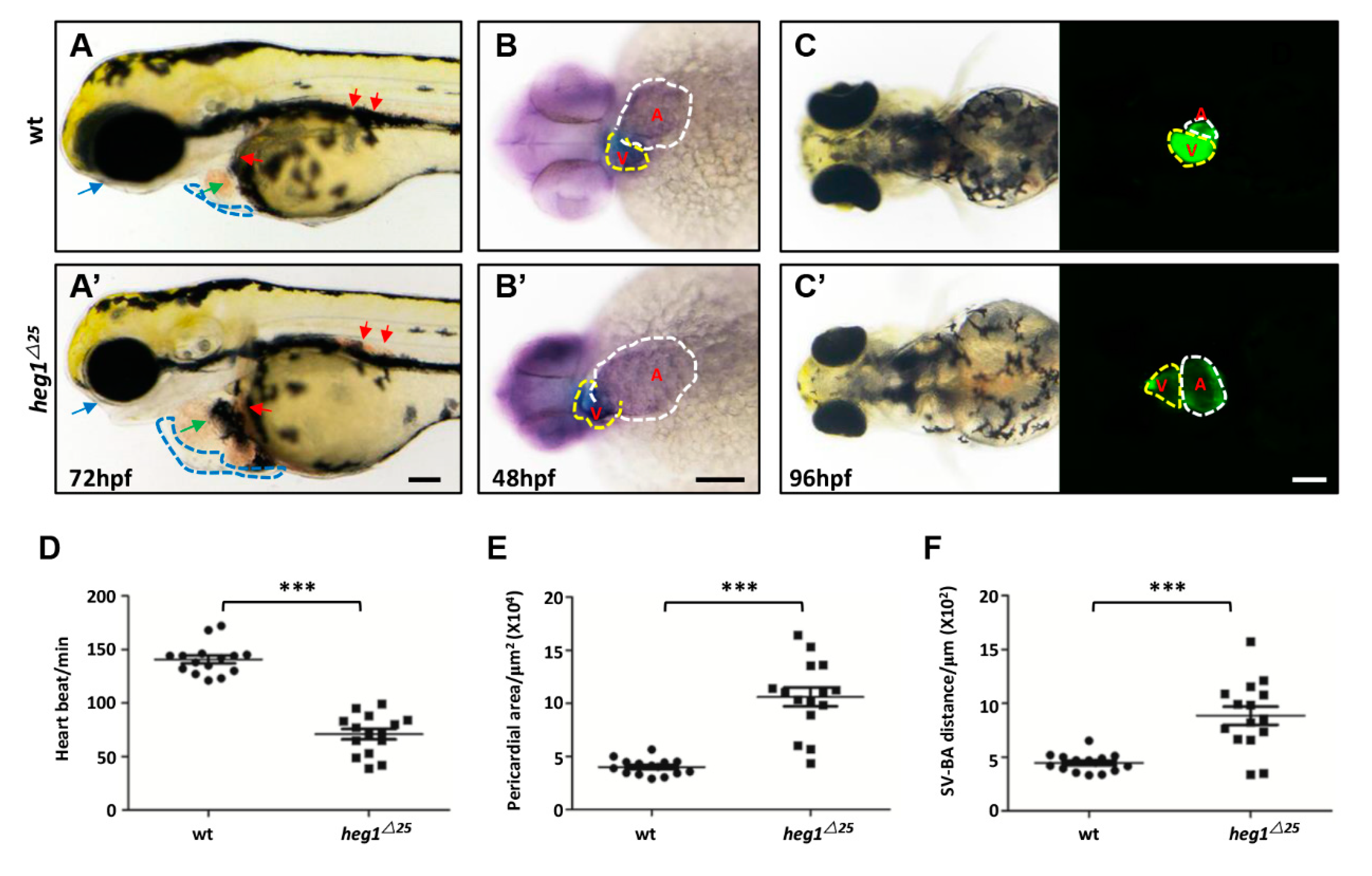
3.3. Abnormal Cardiac Formation in heg1∆25 Mutants
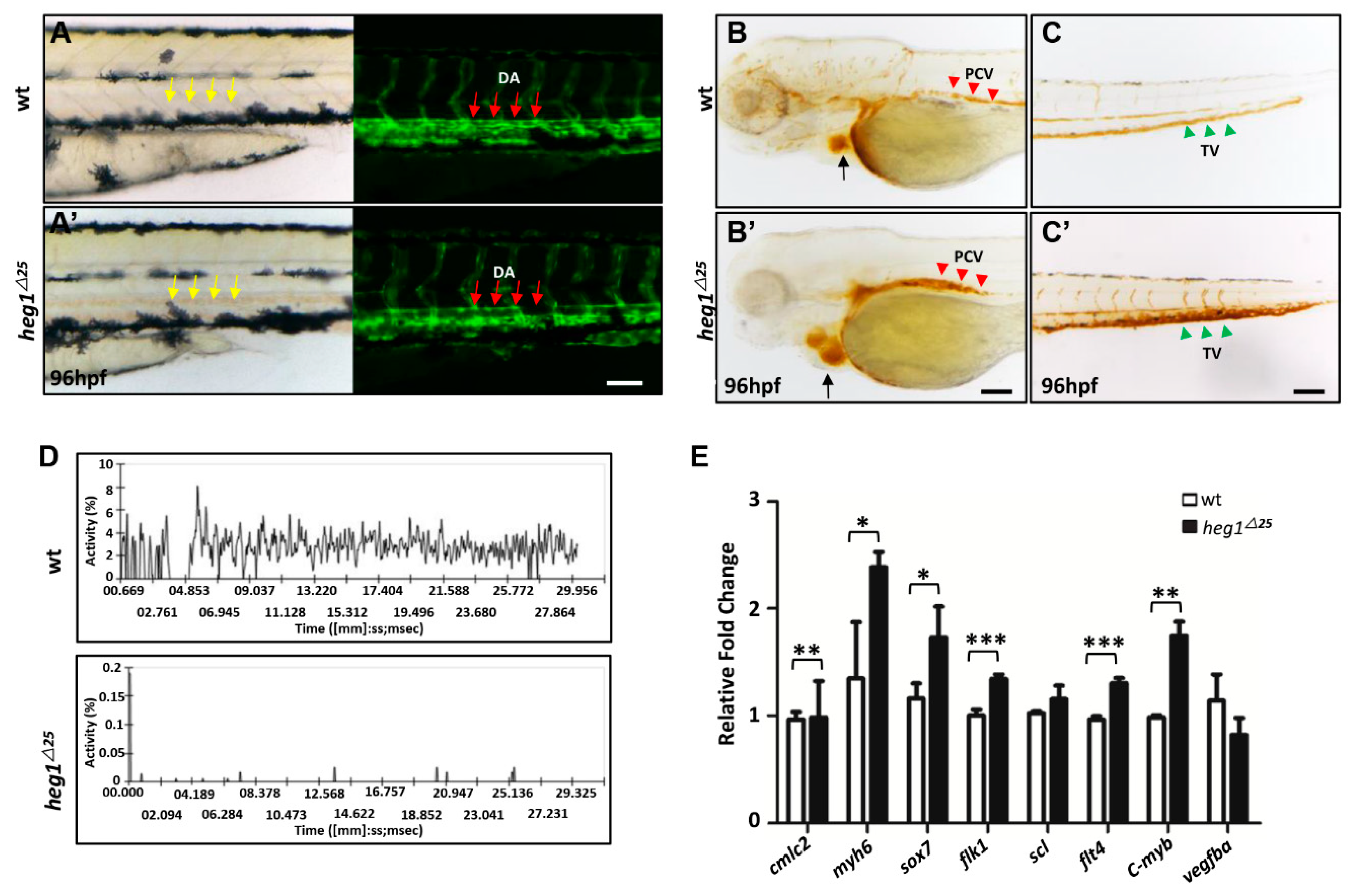
3.4. Abnormal Vascular Development and Blood Stagnation in heg1∆25 Mutants
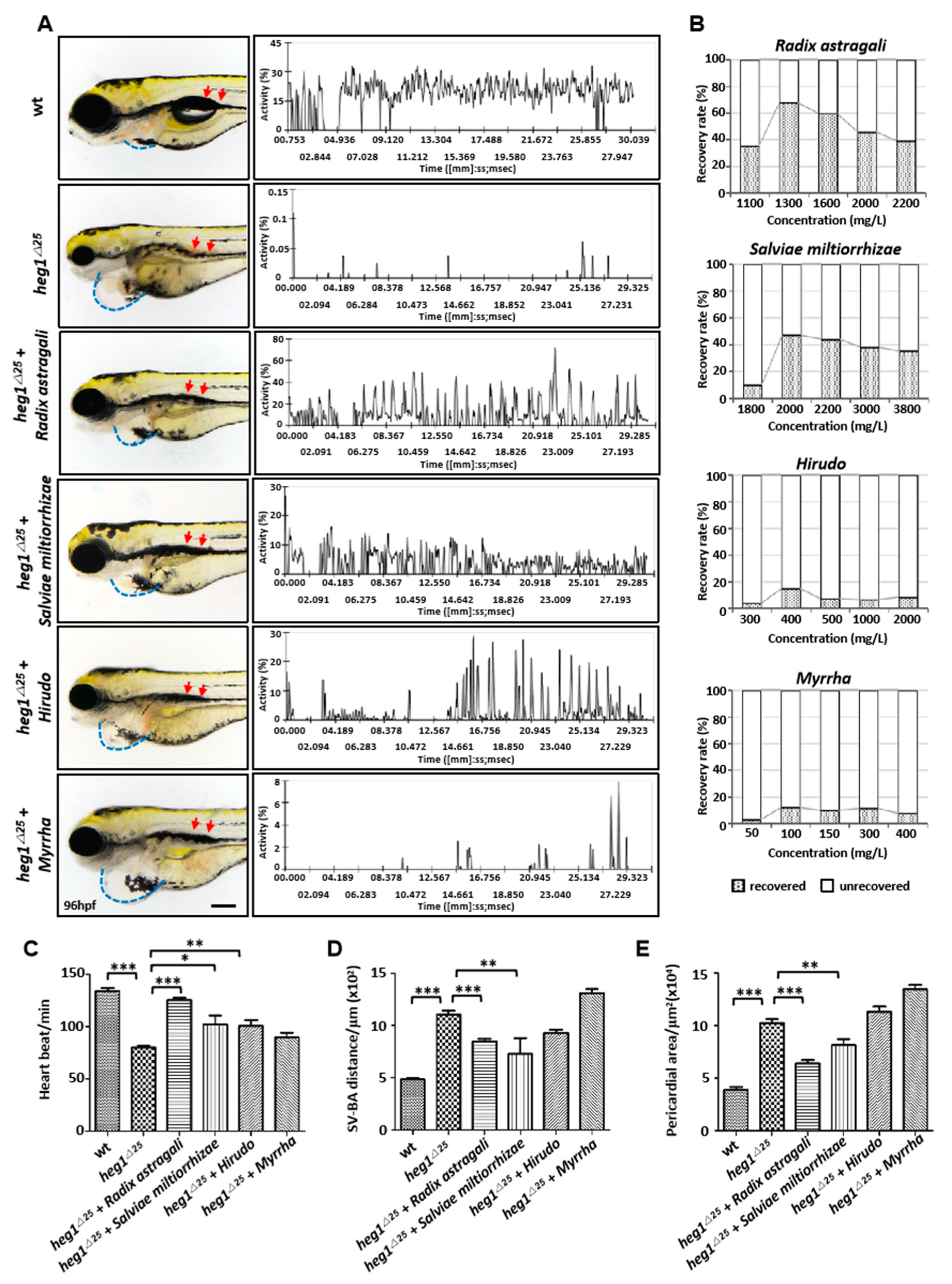
3.5. Application of heg1∆25 Mutants in Screening TCM Herbs for Cardiovascular Diseases Treatment
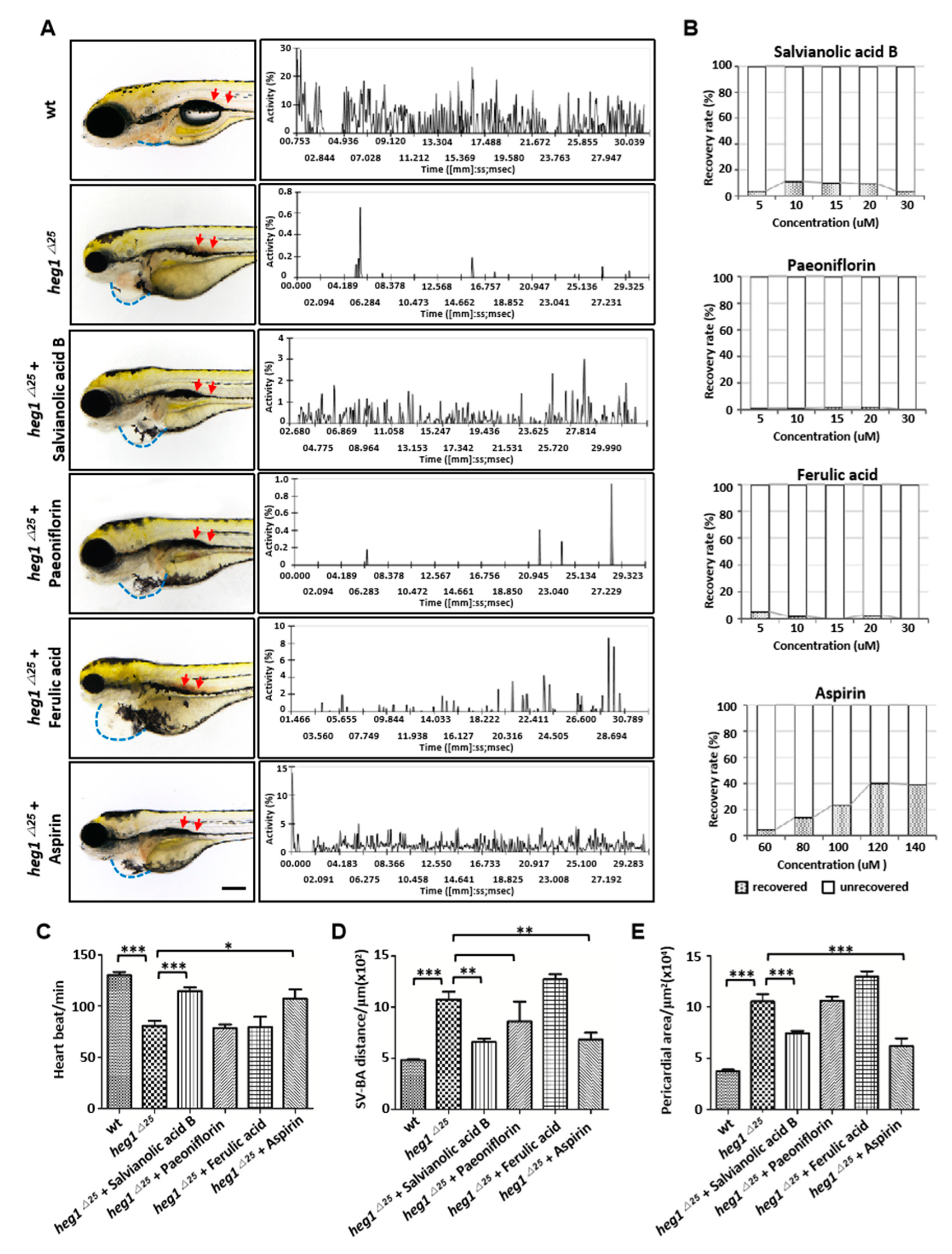
3.6. Application of heg1∆25 mutants in Screening monomers for Cardiovascular Diseases Treatment
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Fox, C.S.; Coady, S.F.; Sorlie, P.F.; D’Agostino, R.B.; Pencina, M.F.; Vasan, J.B.; Meigs, D.; Levy, P.J.; Savage, P.J. Increasing cardiovascular disease burden due to diabetes mellitus: The Framingham Heart Study. Circulation 2007, 12, 1544–1550. [Google Scholar]
- Dagenais, G.R.; Leong, D.P.; Rangarajan, S.; Lanas, F.; Lopez-Jaramillo, P.; Gupta, R.; Diaz, R.; Avezum, A.; Oliveira, G.B.F.; Wielgosz, A.; et al. Variations in common diseases, hospital admissions, and deaths in middle-aged adults in 21 countries from five continents (PURE): A prospective cohort study. Lancet 2020, 10226, 785–794. [Google Scholar]
- Staudt, D.; Stainier, D. Uncovering the molecular and cellular mechanisms of heart development using the zebrafish. Annu. Rev. Genet. 2012, 46, 397–418. [Google Scholar]
- Zakaria, Z.Z.; Benslimane, F.M.; Nasrallah, G.K.; Shurbaji, S.; Younes, N.N.; Mraiche, F.; Da’as, S.I.; Yalcin, H.C. Using Zebrafish for Investigating the Molecular Mechanisms of Drug-Induced Cardiotoxicity. Biomed. Res. Int. 2018, 2018, 1642684. [Google Scholar]
- Nguyen, C.T.; Lu, Q.; Wang, Y.; Chen, J.N. Zebrafish as a model for cardiovascular development and disease. Drug Discov. Today Dis. Models. 2008, 5, 135–140. [Google Scholar]
- Korzh, S.; Pan, X.M.; Garcia-Lecea, C.L.; Winata, C.X.; Pan, X.T.; Wohland, T.V.; Korzh, V.Z.; Gong, Z. Requirement of vasculogenesis and blood circulation in late stages of liver growth in zebrafish. BMC Dev. Biol. 2008, 16, 8–84. [Google Scholar]
- Brönnimann, D.; Annese, T.; Gorr, T.A.; Djonov, V. Splitting of circulating red blood cells as an in vivo mechanism of erythrocyte maturation in developing zebrafish, chick and mouse embryos. J. Exp. Biol. 2018, 221, jeb184564. [Google Scholar]
- Jagadeeswaran, P.; Sheehan, J.P.; Craig, F.E.; Troyer, D. Identification and characterization of zebrafish thrombocytes. Br. J. Haematol. 1999, 107, 731–738. [Google Scholar]
- Sheehan, J.; Templer, M.; Gregory, M.; Hanumanthaiah, R.; Troyer, D.T.; Phan, T.B.; Thankavel, B.P.; Jagadeeswaran, P. Demonstration of the extrinsic coagulation pathway in teleostei: Identification of zebrafish coagulation factor VII. Proc. Natl. Acad. Sci. USA 2001, 98, 8768–8773. [Google Scholar]
- Parng, C.; Seng, W.C.; Semino, C.P.; McGrath, P. Zebrafish: A preclinical model for drug screening. Assay Drug Dev. Technol. 2002, 1, 41–48. [Google Scholar]
- Zhu, X.Y.; Liu, H.C.; Guo, S.Y.; Xia, B.; Song, R.S.; Lao, Q.C.; Xuan, Y.X.; Li, C.Q. A Zebrafish Thrombosis Model for Assessing Antithrombotic Drugs. Zebrafish 2016, 13, 335–344. [Google Scholar] [PubMed]
- Lee, I.J.; Yang, Y.C.; Hsu, J.W.; Chang, W.T.; Chuang, Y.J.; Liau, I. Zebrafish model of photochemical thrombosis for translational research and thrombolytic screening in vivo. J. Biophotonics 2017, 10, 494–502. [Google Scholar] [PubMed]
- Qi, Y.; Zhao, X.; Liu, H.; Wang, Y.; Zhao, C.; Zhao, T.; Zhao, B.; Wang, Y. Identification of a Quality Marker (Q-Marker) of Danhong Injection by the Zebrafish Thrombosis Model. J. Biophotonics 2017, 10, 494–502. [Google Scholar]
- Zaidi, S.; Brueckner, M. Genetics and Genomics of Congenital Heart Disease. Circ. Res. 2017, 120, 923–940. [Google Scholar]
- Mably, J.D.; Mohideen, C.G.; Burns, J.N.; Chen, M.C.; Fishman, M.C. Heart of glass regulates the concentric growth of the heart in zebrafish. Curr. Biol. 2003, 13, 2138–2147. [Google Scholar]
- Donat, S.; Lourenço, M.; Paolini, A.; Otten, C.; Renz, M.; Abdelilah-Seyfried, S. Heg1 and Ccm1/2 proteins control endocardial mechanosensitivity during zebrafish valvulogenesis. Elife 2018, 7, e28939. [Google Scholar]
- Kleaveland, B.; Zheng, X.; Liu, J.J.; Blum, Y.; Tung, J.J.; Zou, Z.; Sweeney, S.M.; Chen, M.; Guo, L.; Lu, M.M.; et al. Regulation of cardiovascular development and integrity by the heart of glass-cerebral cavernous malformation protein pathway. Nat. Med. 2009, 15, 169–176. [Google Scholar]
- Monaco, A.P.; Neve, R.L.; Colletti-Feener, C.; Bertelson, C.J.; Kurnit, D.M.; Kunkel, L.M. Isolation of candidate cDNAs for portions of the Duchenne muscular dystrophy gene. Nature 1986, 323, 646–650. [Google Scholar]
- Kawahara, G.; Karpf, J.A.; Myers, J.A.; Alexander, M.S.; Guyon, J.R.; Kunkel, L.M. Drug screening in a zebrafish model of Duchenne muscular dystrophy. Proc. Natl. Acad. Sci. USA 2011, 108, 5331–5336. [Google Scholar]
- Baraban, S.C.; Dinday, M.T.; Hortopan, G.A. Drug screening in Scn1a zebrafish mutant identifies clemizole as a potential Dravet syndrome treatment. Nat. Commun. 2013, 4, 2410. [Google Scholar]
- Westerfield, M. The Zebrafish Book: Guide for the Laboratory Use of Zebrafish (Danio Rerio); University of Oregon Press: Eugene, UO, USA, 2000. [Google Scholar]
- Kimmel, C.B.; Ballard, S.R.; Kimmel, S.B.; Ullmann, B.; Schilling, T.F. Stages of embryonic development of the zebrafish. Dev. Dyn. 1995, 203, 253–310. [Google Scholar] [CrossRef]
- Grimes, A.C.; Erwin, K.N.; Stadt, H.A.; Hunter, G.L.; Gefroh, H.A.; Tsai, H.J.; Kirby, M.L. PCB126 exposure disrupts zebrafish ventricular and branchial but not early neural crest development. Toxicol. Sci. 2008, 106, 193–205. [Google Scholar] [CrossRef]
- Gkatzis, K.; Thalgott, J.; Dos, D.; Martin, S.; Lamandé, N.; Carette, M.F.; Disch, F.; Snijder, R.J. Interaction Between ALK1 Signaling and Connexin40 in the Development of Arteriovenous Malformations. Arterioscler. Thromb. Vasc. Biol. 2016, 36, 707–717. [Google Scholar] [CrossRef]
- Burns, C.G.; MacRae, C.A. Purification of hearts from zebrafish embryos. Biotechniques 2006, 40, 274–278. [Google Scholar]
- Mali, P.; Yang, L.; Esvelt, K.M.; Aach, J.; Guell, M.; DiCarlo, J.E.; Norville, J.E.; Church, G.M. RNA-guided human genome engineering via Cas9. Science 2013, 339, 823–826. [Google Scholar] [CrossRef]
- Liu, C.X.; Li, C.Y.; Hu, C.C.; Wang, Y.; Lin, J.; Jiang, Y.H.; Li, Q.; Xu, X. CRISPR/Cas9-induced shank3b mutant zebrafish display autism-like behaviors. Mol. Autism. 2018, 9, 23. [Google Scholar] [CrossRef]
- Moreno-Mateos, M.A.; Vejnar, C.E.; Beaudoin, J.D.; Fernandez, J.P.; Mis, E.K.; Khokha, M.K.; Giraldez, A.J. CRISPRscan: Designing highly efficient sgRNAs for CRISPR-Cas9 targeting in vivo. Nat. Methods 2015, 12, 982–988. [Google Scholar] [CrossRef]
- Tian, J.; Shao, J.; Liu, C.; Hou, H.Y.; Chou, C.W.; Shboul, M.; Li, G.Q.; El-Khateeb, M.; Samarah, O.Q.; Kou, Y.; et al. Deficiency of lrp4 in zebrafish and human LRP4 mutation induce aberrant activation of Jagged-Notch signaling in fin and limb development. Cell. Mol. Life Sci. 2019, 76, 163–178. [Google Scholar] [CrossRef]
- Tian, J.; Ling, L.; Shboul, M.; Lee, H.; O’Connor, B.; Merriman, B.; Nelson, S.F.; Cool, S.; Ababneh, O.H.; Al-Hadidy, A.; et al. Loss of CHSY1, a secreted FRINGE enzyme, causes syndromic brachydactyly in humans via increased NOTCH signaling. Am. J. Hum. Genet. 2010, 87, 768–778. [Google Scholar] [CrossRef]
- Bhakta, M.; Padanad, M.S.; Harris, J.P.; Lubczyk, C.; Amatruda, J.F.; Munshi, N.A.-O.X. pouC Regulates Expression of bmp4 During Atrioventricular Canal Formation in Zebrafish. Dev. Dyn. 2019, 248, 173–188. [Google Scholar] [CrossRef]
- Korzh, S.; Winata, C.L.; Zheng, W.; Yang, S.; Yin, A.; Ingham, P.; Korzh, V.; Gong, Z. The interaction of epithelial Ihha and mesenchymal Fgf10 in zebrafish esophageal and swimbladder development. Dev. Biol. 2011, 359, 262–276. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.C.; Lawson, N.D.; Weinstein, B.M.; Johnson, S.L. reg6 is required for branching morphogenesis during blood vessel regeneration in zebrafish caudal fins. Dev. Biol. 2003, 264, 263–274. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Du, L.; Xu, J.; Li, X.; Ma, N.; Liu, Y.; Peng, J.; Osato, M.; Zhang, W.; Wen, Z. Rumba and Haus3 are essential factors for the maintenance of hematopoietic stem/progenitor cells during zebrafish hematopoiesis. Development 2011, 138, 619–629. [Google Scholar] [CrossRef]
- Hu, G.; Mahady, G.B.; Li, S.; Hoi, M.P.; Wang, Y.H.; Lee, S.M. Polysaccharides from astragali radix restore chemical-induced blood vessel loss in zebrafish. Vasc. Cell 2012, 4, 2. [Google Scholar] [CrossRef]
- Tse, H.Y.; Hui, M.N.; Li, L.; Lee, S.M.; Leung, A.Y.; Cheng, S.H. Angiogenic efficacy of simplified 2-herb formula (NF3) in zebrafish embryos in vivo and rat aortic ring in vitro. J. Ethnopharmacol. 2012, 139, 447–453. [Google Scholar] [CrossRef]
- Adams, J.D.; Wang, R.; Yang, J.; Lien, E.J. Preclinical and clinical examinations of Salvia miltiorrhiza and its tanshinones in ischemic conditions. Chin. Med. 2006, 23, 3. [Google Scholar] [CrossRef]
- Buchwald, A.B.; Hammerschmidt, S.; Stevens, J.; Göring, J.; Nebendahl, K.; Unterberg, C. Inhibition of neointimal proliferation after coronary angioplasty by low-molecular-weight heparin (clivarine) and polyethyleneglycol-hirudin. J. Cardiovasc. Pharmacol. 1996, 28, 481–487. [Google Scholar] [CrossRef]
- Shedoeva, A.; Leavesley, D.; Upton, Z.; Fan, C. Wound Healing and the Use of Medicinal Plants. Evid. Based Complement. Alternat. Med. 2019, 2019, 2684108. [Google Scholar] [CrossRef]
- Wang, J.; Xiong, X.; Feng, B. Cardiovascular effects of salvianolic Acid B. Evid. Based Complement. Alternat. Med. 2013, 2013, 247948. [Google Scholar] [CrossRef]
- Li, W.; Zhi, W.; Liu, F.; Zhao, J.; Yao, Q.; Niu, X. Paeoniflorin inhibits VSMCs proliferation and migration by arresting cell cycle and activating HO-1 through MAPKs and NF-κB pathway. Int. Immunopharmacol. 2018, 54, 103–111. [Google Scholar] [CrossRef]
- Chen, H.; Dong, Y.; He, X.; Li, J.; Wang, J. Paeoniflorin improves cardiac function and decreases adverse postinfarction left ventricular remodeling in a rat model of acute myocardial infarction. Drug Des. Devel. Ther. 2018, 12, 823–836. [Google Scholar] [CrossRef] [PubMed]
- Alam, M.A.; Sernia, C.; Brown, L. Ferulic acid improves cardiovascular and kidney structure and function in hypertensive rats. J. Cardiovasc. Pharmacol. 2013, 61, 240–249. [Google Scholar] [CrossRef] [PubMed]
- Chowdhury, S.; Ghosh, S.; Rashid, K.; Sil, P.C. Deciphering the role of ferulic acid against streptozotocin-induced cellular stress in the cardiac tissue of diabetic rats. Food Chem. Toxicol. 2016, 97, 187–198. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Huang, Y. Chinese Herbal Medicine on Cardiovascular Diseases and the Mechanisms of Action. Front. Pharmacol. 2016, 7, 469. [Google Scholar] [CrossRef] [PubMed]
- Fu, S.; Zhang, J.; Menniti-Ippolito, F.; Gao, X.; Galeotti, F.; Massari, M.; Hu, L.; Zhang, B.; Ferrelli, R.; Fauci, A.; et al. Huangqi injection (a traditional Chinese patent medicine) for chronic heart failure: A systematic review. PLoS ONE 2011, 6, e19604. [Google Scholar] [CrossRef]
- Ma, X.; Zhang, K.; Li, H.; Han, S.; Ma, Z.; Tu, P. Extracts from Astragalus membranaceus limit myocardial cell death and improve cardiac function in a rat model of myocardial ischemia. J. Ethnopharmacol. 2013, 149, 720–728. [Google Scholar] [CrossRef]
- Ding, M.; Ye, T.X.; Zhao, G.R.; Yuan, Y.J.; Guo, Z.X. Aqueous extract of Salvia miltiorrhiza attenuates increased endothelial permeability induced by tumor necrosis factor-alpha. Int. Immunopharmacol. 2005, 5, 1641–1651. [Google Scholar] [CrossRef]
- Wang, X.; Wang, Y.; Jiang, M.; Zhu, Y.; Hu, L.; Fan, G.; Wang, Y.; Li, X.; Gao, X. Differential cardioprotective effects of salvianolic acid and tanshinone on acute myocardial infarction are mediated by unique signaling pathways. J. Ethnopharmacol. 2011, 135, 662–671. [Google Scholar] [CrossRef]
- Ren, J.; Fu, L.; Nile, S.H.; Zhang, J.; Kai, G. Salvia miltiorrhiza in Treating Cardiovascular Diseases: A Review on Its Pharmacological and Clinical Applications. Front. Pharmacol. 2019, 10, 753. [Google Scholar] [CrossRef]
- Liu, J.F.; Hu, A.N.; Zan, J.F.; Wang, P.; You, Q.Y.; Tan, A.H. Network Pharmacology Deciphering Mechanisms of Volatiles of Wendan Granule for the Treatment of Alzheimer’s Disease. Evid. Based Complement. Alternat. Med. 2019, 2018, 7826769. [Google Scholar] [CrossRef]
- Li, L.; Zhou, X.; Li, N.; Sun, M.; Lv, J.; Xu, Z. Herbal drugs against cardiovascular disease: Traditional medicine and modern development. Drug Discov. Today 2015, 20, 1074–1086. [Google Scholar] [CrossRef] [PubMed]
- Gu, G.; Na, Y.; Chung, H.; Seok, S.H.; Lee, H.Y. Zebrafish Larvae Model of Dilated Cardiomyopathy Induced by Terfenadine. Korean Circ. J. 2017, 47, 960–969. [Google Scholar] [CrossRef] [PubMed]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lu, S.; Hu, M.; Wang, Z.; Liu, H.; Kou, Y.; Lyu, Z.; Tian, J. Generation and Application of the Zebrafish heg1 Mutant as a Cardiovascular Disease Model. Biomolecules 2020, 10, 1542. https://doi.org/10.3390/biom10111542
Lu S, Hu M, Wang Z, Liu H, Kou Y, Lyu Z, Tian J. Generation and Application of the Zebrafish heg1 Mutant as a Cardiovascular Disease Model. Biomolecules. 2020; 10(11):1542. https://doi.org/10.3390/biom10111542
Chicago/Turabian StyleLu, Shuxian, Mengyan Hu, Zhihao Wang, Hongkai Liu, Yao Kou, Zhaojie Lyu, and Jing Tian. 2020. "Generation and Application of the Zebrafish heg1 Mutant as a Cardiovascular Disease Model" Biomolecules 10, no. 11: 1542. https://doi.org/10.3390/biom10111542
APA StyleLu, S., Hu, M., Wang, Z., Liu, H., Kou, Y., Lyu, Z., & Tian, J. (2020). Generation and Application of the Zebrafish heg1 Mutant as a Cardiovascular Disease Model. Biomolecules, 10(11), 1542. https://doi.org/10.3390/biom10111542