Identification of Feldin, an Antifungal Polyyne from the Beefsteak Fungus Fistulina hepatica

Abstract

1. Introduction
2. Materials and Methods
2.1. Fungal Material, Fungal Cultivation, and Media
2.2. Microscopy
2.3. Extraction
2.4. Liquid Chromatography/Mass Spectrometry
2.5. NMR Spectroscopy
2.6. Antifungal and Antibacterial Assays
3. Results
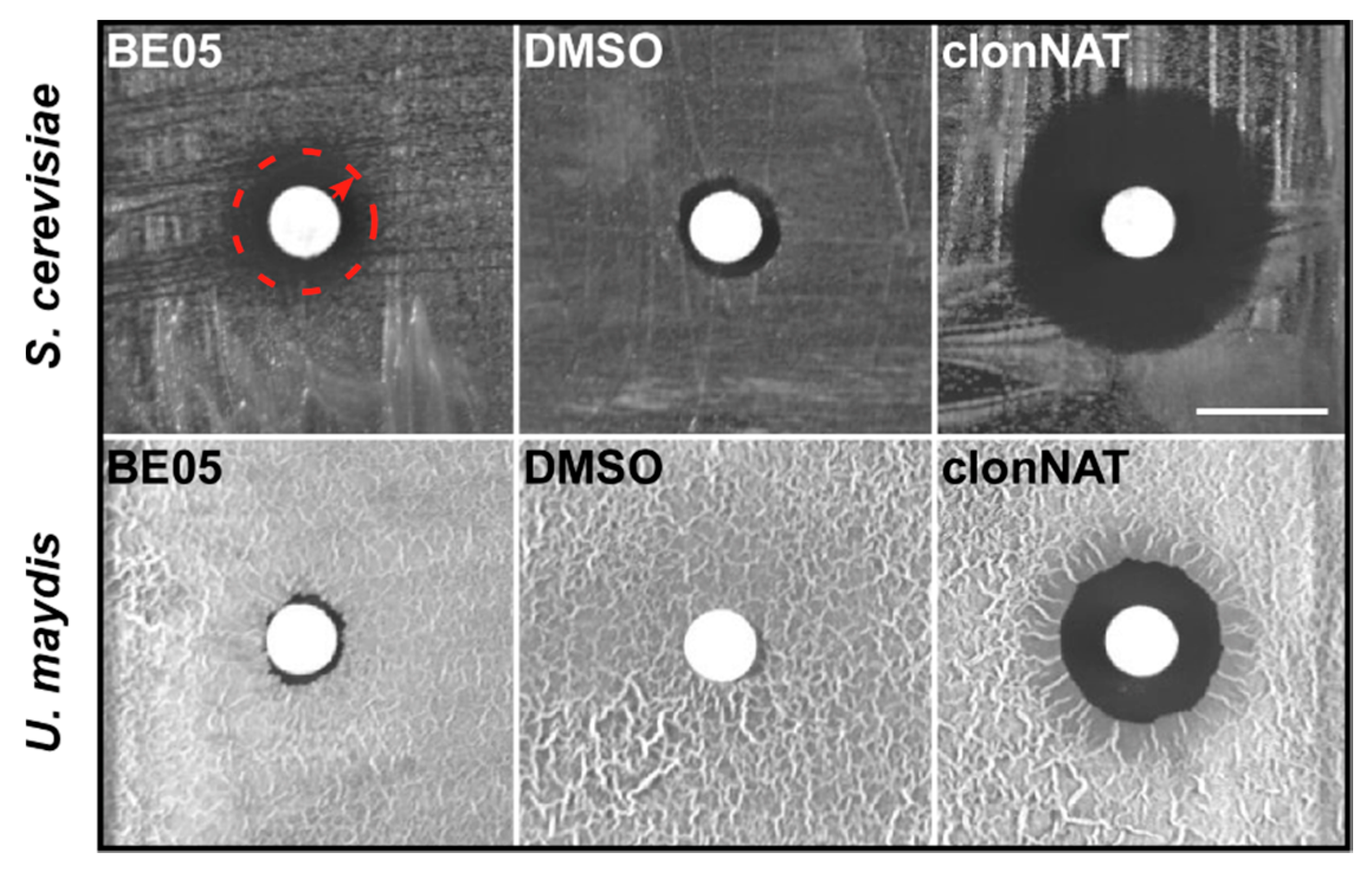
3.1. Bioactivity Tests with Ethyl Acetate Extracts from 41 Basidiomycete Mushroom Strains
3.2. Characteristics of Fistulina hepatica
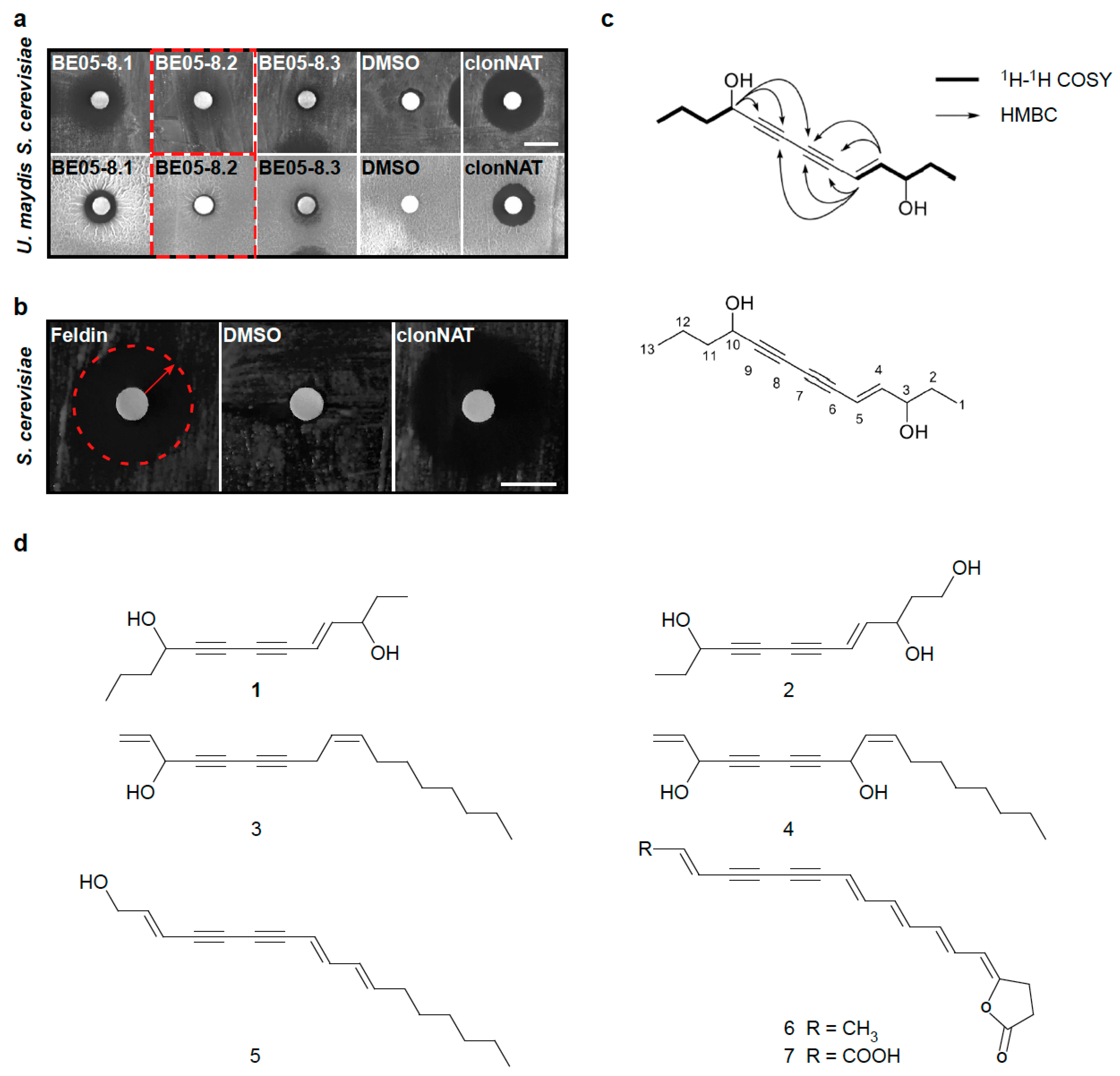
3.3. Identification of the Antifungal Polyyne Feldin from Basidiomycete Extract BE05
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- He, M.; Zhao, R.; Hyde, K.D.; Begerow, D.; Kemler, M.; Yurkov, A.; McKenzie, E.H.C.; Raspé, O.; Kakishima, M.; Sánchez-Ramírez, S.; et al. Notes, outline and divergence times of Basidiomycota. Fungal Divers. 2019, 99, 105–367. [Google Scholar] [CrossRef]
- Sandargo, B.; Chepkirui, C.; Cheng, T.; Chaverra-Munoz, L.; Thongbai, B.; Stadler, M.; Hüttel, S. Biological and chemical diversity go hand in hand: Basidiomycota as source of new pharmaceuticals and agrochemicals. Biotechnol. Adv. 2019, 37, 107344. [Google Scholar] [CrossRef] [PubMed]
- Zhao, R.-L.; Li, G.-J.; Sánchez-Ramírez, S.; Stata, M.; Yang, Z.L.; Wu, G.; Dai, Y.C.; He, S.H.; Cui, B.K.; Zhou, J.L.; et al. A six-gene phylogenetic overview of Basidiomycota and allied phyla with estimated divergence times of higher taxa and a phyloproteomics perspective. Fungal Divers. 2017, 84, 43–74. [Google Scholar] [CrossRef]
- Künzler, M. How fungi defend themselves against microbial competitors and animal predators. PLoS Pathog. 2018, 14, e1007184. [Google Scholar] [CrossRef] [PubMed]
- Hyde, K.D.; Xu, J.; Rapior, S.; Jeewon, R.; Lumyong, S.; Niego, A.G.T.; Abeywickrama, P.D.; Aluthmuhandiram, J.V.S.; Brahamanage, R.S.; Brooks, S.; et al. The amazing potential of fungi: 50 ways we can exploit fungi industrially. Fungal Divers. 2019, 97, 1–136. [Google Scholar] [CrossRef]
- Tayyrov, A.; Azevedo, S.; Herzog, R.; Vogt, E.; Arzt, S.; Lüthy, P.; Müller, P.; Rühl, M.; Hennicke, F.; Künzler, M. Heterologous production and functional characterization of ageritin, a novel type of ribotoxin highly expressed during fruiting of the edible mushroom Agrocybe aegerita. Appl. Environ. Microbiol. 2019, 85, e01549-19. [Google Scholar] [CrossRef]
- Alberti, F.; Khairudin, K.; Venegas, E.; Davies, J.A.; Hayes, P.M.; Willis, C.L.; Bailey, A.M.; Foster, G.D. Heterologous expression reveals the biosynthesis of the antibiotic pleuromutilin and generates bioactive semi-synthetic derivatives. Nat. Commun. 2017, 8, 1831. [Google Scholar] [CrossRef] [PubMed]
- Nofiani, R.; de Mattos-Shipley, K.; Lebe, K.E.; Han, L.-C.; Iqbal, Z.; Bailey, A.M.; Willis, C.L.; Simpson, T.J.; Cox, R.J. Strobilurin biosynthesis in basidiomycete fungi. Nat. Commnun. 2018, 9, 3940. [Google Scholar] [CrossRef] [PubMed]
- Engels, B.; Heinig, U.; Grothe, T.; Stadler, M.; Jennewein, S. Cloning and characterization of an Armillaria gallica cDNA encoding protoilludene synthase, which catalyzes the first committed step in the synthesis of antimicrobial melleolides. J. Biol. 2011, 286, 6871–6878. [Google Scholar] [CrossRef]
- Croteau, R.; Ketchum, R.E.; Long, R.M.; Kaspera, R.; Wildung, M.R. Taxol biosynthesis and molecular genetics. Phytochem. Rev. 2006, 5, 75–97. [Google Scholar] [CrossRef]
- Dejong, J.M.; Liu, Y.; Bollon, A.P.; Long, R.M.; Jennewein, S.; Williams, D.; Croteau, R.B. Genetic engineering of taxol biosynthetic genes in Saccharomyces cerevisiae. Biotechnol. Bioeng. 2006, 93, 212–224. [Google Scholar] [CrossRef] [PubMed]
- Paddon, C.J.; Westfall, P.J.; Pitera, D.J.; Benjamin, K.; Fisher, K.; McPhee, D.; Leavell, M.D.; Tai, A.; Main, A.; Eng, D.; et al. High-level semi-synthetic production of the potent antimalarial artemisinin. Nature 2013, 496, 528–532. [Google Scholar] [CrossRef] [PubMed]
- Schlingmann, G.; Milne, L.; Pearce, C.J.; Borders, D.B.; Greenstein, M.; Maiese, W.M.; Carter, G.T. Isolation, characterization and structure of a new allenic polyine antibiotic produced by fungus LL-07F275. J. Antibiot. Res. 1995, 48, 375–379. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Costa-de-Oliveira, S.; Rodrigues, A.G. Candida albicans antifungal resistance and tolerance in bloodstream infections: The triad yeast-host-antifungal. Microorganisms 2020, 8, 154. [Google Scholar] [CrossRef] [PubMed]
- Matheny, P.B.; Curtis, J.M.; Hofstetter, V.; Aime, M.C.; Moncalvo, J.M.; Ge, Z.W.; Slot, J.C.; Ammirati, J.F.; Baroni, T.J.; Bougher, N.L.; et al. Major clades of Agaricales: A multilocus phylogenetic overview. Mycologia 2006, 98, 982–995. [Google Scholar] [CrossRef]
- Floudas, D.; Held, B.W.; Riley, R.; Nagy, L.G.; Koehler, G.; Ransdell, A.S.; Younus, H.; Chow, J.; Chiniquy, J.; Lipzen, A.; et al. Evolution of novel wood decay mechanisms in Agaricales revealed by the genome sequences of Fistulina hepatica and Cylindrobasidium torrendii. Fungal Genet. Biol. 2015, 76, 78–92. [Google Scholar] [CrossRef]
- Kaffenberger, J.T.; Schilling, J.S. Comparing lignocellulose physiochemistry after decomposition by brown rot fungi with distinct evolutionary origins. Environ. Microbiol. 2015, 17, 4885–4897. [Google Scholar] [CrossRef]
- Regué, A.; Bassié, L.; de-Miguel, S.; Colinas, C. Environmental and stand conditions related to Fistulina hepatica heart rot attack on Castanea sativa. For. Pathol. 2019, 49, e12517. [Google Scholar] [CrossRef]
- Jones, E.R.H.; Lowe, G.; Shannon, P.V.R. Natural acetylenes. Part XX. Tetra-acetylenic and other metabolites from Fistulina hepatica (Huds) Fr. J. Chem. Soc. C 1966, 139–144. [Google Scholar] [CrossRef]
- Ivanova, V.; Kolarova, M.; Aleksieva, K.; Schlegel, R.; Schumann, P.; Graefe, U. Octadeca-8,11-dienoic acid methylester, a new fatty acid metabolite from Fistulina hepatica. J. Mod. Med. 2013, 1, 43–48. [Google Scholar] [CrossRef]
- Tsuge, N.; Mori, T.; Hamano, T.; Tanaka, H.; Shin-ya, K.; Seto, H. Cinnatriacetins A and B, new antibacterial triacetylene derivatives from the fruiting bodies of Fistulina hepatica. J. Antibiot. Res. 1999, 52, 578–581. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Vaz, J.A.; Barros, L.; Martins, A.; Morais, J.S.; Vasconcelos, M.H.; Ferreira, I.C.F.R. Phenolic profile of seventeen Portuguese wild mushrooms. LWT Food Sci. Technol. 2011, 44, 343–346. [Google Scholar] [CrossRef]
- Wu, S.; Krings, U.; Zorn, H.; Berger, R.G. Volatile compounds from the fruiting bodies of beefsteak fungus Fistulina hepatica (Schaeffer: Fr.) Fr. Food Chem. 2005, 92, 221–226. [Google Scholar] [CrossRef]
- Wu, S.; Zorn, H.; Krings, U.; Berger, R.G. Volatiles from submerged and surface-cultured beefsteak fungus, Fistulina hepatica. Flavour Frag. J. 2007, 22, 53–60. [Google Scholar] [CrossRef]
- Garrod, B.; Lewis, B.G. Location of the antifungal compound falcarindiol in carrot root tissue. Trans. Brit. 1979, 72, 515–517. [Google Scholar] [CrossRef]
- Garrod, B.; Lea, E.J.A.; Lewis, B.G. Studies on the mechanism of action of the antifungal compound falcarindiol. New Phytol. 1979, 83, 463–471. [Google Scholar] [CrossRef]
- Minto, R.E.; Blacklock, B.J. Biosynthesis and function of polyacetylenes and allied natural products. Prog. Lipid Res. 2008, 47, 233–306. [Google Scholar] [CrossRef]
- Stevens, R. Mycology Guidebook; University of Washington Press: Seattle, WA, USA; London, UK, 1974; p. 719. [Google Scholar]
- Baumann, S.; Zander, S.; Weidtkamp-Peters, S.; Feldbrügge, M. Live cell imaging of septin dynamics in Ustilago maydis. Methods Cell Biol. 2016, 136, 143–159. [Google Scholar] [CrossRef]
- Jankowski, S.; Pohlmann, T.; Baumann, S.; Muntjes, K.; Devan, S.K.; Zander, S.; Feldbrügge, M. The multi PAM2 protein Upa2 functions as novel core component of endosomal mRNA transport. EMBO Rep. 2019, 20, e47381. [Google Scholar] [CrossRef]
- Harwoko, H.; Daletos, G.; Stuhldreier, F.; Lee, J.; Wesselborg, S.; Feldbrügge, M.; Müller, W.E.G.; Kalscheuer, R.; Ancheeva, E.; Proksch, P. Dithiodiketopiperazine derivatives from endophytic fungi Trichoderma harzianum and Epicoccum nigrum. Nat. Prod. Res. 2019, 18, 1–9. [Google Scholar] [CrossRef]
- Brachmann, A.; Weinzierl, G.; Kämper, J.; Kahmann, R. Identification of genes in the bW/bE regulatory cascade in Ustilago maydis. Mol. Microbiol. 2001, 42, 1047–1063. [Google Scholar] [CrossRef] [PubMed]
- Pereira, G.; Tanaka, T.U.; Nasmyth, K.; Schiebel, E. Modes of spindle pole body inheritance and segregation of the Bfa1p-Bub2p checkpoint protein complex. EMBO J. 2001, 20, 6359–6370. [Google Scholar] [CrossRef]
- Holliday, R. Molecular aspects of genetic exchange and gene conversion. Genetics 1974, 78, 273–287. [Google Scholar] [PubMed]
- Banuett, F.; Herskowitz, I. Different a alleles of Ustilago maydis are necessary for maintenance of filamentous growth but not for meiosis. Proc. Natl. Acad. Sci. USA 1989, 86, 5878–5882. [Google Scholar] [CrossRef] [PubMed]
- Abe, S.; Takayarna, K.; Kinoshita, S. Taxonomical studies on glutamic acid producing bacteria. J. Gen. Appl. Microbiol. 1967, 13, 279–301. [Google Scholar] [CrossRef]
- Knudsen, H.; Vesterholt, J. Funga Nordica; Nordsvamp: Copenhagen, Denmark, 2008; 966p. [Google Scholar]
- Krieglsteiner, G.J. Die Grosspilze Baden-Württembergs. Bd. 1: Allgemeiner Teil, Ständerpilze: Gallert-Rinden-, Stachel- und Porenpilze; Ulmer: Stuttgart, Germany, 2000; p. 629. (In German) [Google Scholar]
- Stalpers, J.A.; Vlug, I. Confistulina, the anamorph of Fistulina hepatica. Can. J. Bot. 1983, 61, 1660–1666. [Google Scholar] [CrossRef]
- Griffith, G.W.; Easton, G.L.; Detheridge, A.; Roderick, K.; Edwards, A.; Worgan, H.J.; Nicholson, J.; Perkins, W.T. Copper deficiency in potato dextrose agar causes reduced pigmentation in cultures of various fungi. FEMS Microbiol. Lett. 2007, 276, 165–171. [Google Scholar] [CrossRef]
- Bohlmann, F.; Knoll, K.-H. New acetylenic compounds from Emilia species. Phytochemistry 1978, 17, 557–558. [Google Scholar] [CrossRef]
- Christensen, L.P.; Brandt, K. Bioactive polyacetylenes in food plants of the Apiaceae family: Occurrence, bioactivity and analysis. J. Pharm. Biomed. Anal. 2006, 41, 683–693. [Google Scholar] [CrossRef]
- Kuhnt, D.; Anke, T.; Besl, H.; Bross, M.; Herrmann, R.; Mocek, U.; Steffan, B.; Steglich, W. Antibiotics from basidiomycetes. XXXVII. New inhibitors of cholesterol biosynthesis from cultures of Xerula melanotricha Dörfelt. J. Antibiot. Res. 1990, 43, 1413–1420. [Google Scholar] [CrossRef]
- Suay, I.; Arenal, F.; Asensio, F.J.; Basilio, A.; Cabello, M.A.; Díez, M.T.; García, J.B.; del Val, A.G.; Gorrochategui, J.; Hernández, P.; et al. Screening of basidiomycetes for antimicrobial activities. Antonie Van Leeuwenhoek 2000, 78, 129–139. [Google Scholar] [CrossRef] [PubMed]
- Kjer, J.; Debbab, A.; Aly, A.H.; Proksch, P. Methods for isolation of marine-derived endophytic fungi and their bioactive secondary products. Nat. Protoc. 2010, 3, 479–490. [Google Scholar] [CrossRef]
- Pan, F.; El-Kashef, D.H.; Kalscheuer, R.; Müller, W.E.G.; Lee, J.; Feldbrügge, M.; Mándi, A.; Kurtán, T.; Liu, Z.; Wu, W.; et al. New hybrid polyketides from the endophytic fungus Cladosporium sphaerospermum WBS017. Eur. J. Med. Chem. 2020, 191, 112159. [Google Scholar] [CrossRef] [PubMed]
- Vandermolen, K.M.; Raja, H.A.; El-Elimat, T.; Oberlies, N.H. Evaluation of culture media for the production of secondary metabolites in a natural products screening program. AMB Express 2013, 3, 71. [Google Scholar] [CrossRef] [PubMed]
- Fritsche, K.; Berg, M.V.D.; Boer, W.D.; Beek, T.A.V.; Raaijmakers, J.M.; Veen, J.A.V.; Leveau, J.H.J. Biosynthetic genes and activity spectrum of antifungal polyynes from Collimonas fungivorans Ter331. Environ. Microbiol. 2014, 16, 1334–1345. [Google Scholar] [CrossRef]
- Kobaisy, M.; Abramowski, Z.; Lermer, L.; Saxena, G.; Hancock, R.E.W.; Towers, G.H.N.; Doxsee, D.; Stokes, R.W. Antimycobacterial polyynes of Devil’s Club (Oplopanax horridus), a North American native medicinal plant. J. Nat. Prod. 1997, 60, 1210–1213. [Google Scholar] [CrossRef]
- Lechner, D.; Stavri, M.; Oluwatuyi, M.; Pereda-Miranda, R.; Gibbons, S. The anti-staphylococcal activity of Angelica dahurica (Bai Zhi). Phytochemistry 2004, 65, 331–335. [Google Scholar] [CrossRef]
- Ondeyka, J.G.; Zink, D.L.; Young, K.; Painter, R.; Kodali, S.; Galgoci, A.; Collado, J.; Tormo, J.R.; Basilio, A.; Vicente, F.; et al. Discovery of bacterial fatty acid synthase inhibitors from a Phoma species as antimicrobial agents using a new antisense-based strategy. J. Nat. Prod. 2006, 69, 377–380. [Google Scholar] [CrossRef]
- Ahmed, M.; Barley, G.C.; Hearn, M.T.W.; Jones, E.R.H.; Thaller, V.; Yates, J.A. Natural acetylenes. Part XLIII. Polyacetylenes from cultures of the fungus Fistulina pallida (berk. and rev.). J. Chem. Soc. Perkin Trans. 1974, 1974, 1981–1987. [Google Scholar] [CrossRef]
- Frey-Klett, P.; Burlinson, P.; Deveau, A.; Barret, M.; Tarkka, M.; Sarniguet, A. Bacterial-fungal interactions: Hyphens between agricultural, clinical, environmental, and food microbiologists. Microbiol. Mol. Biol. Rev. 2011, 75, 583–609. [Google Scholar] [CrossRef]
- Berendsen, R.L.; Baars, J.J.; Kalkhove, S.I.; Lugones, L.G.; Wösten, H.A.; Bakker, P.A. Lecanicillium fungicola: Causal agent of dry bubble disease in white-button mushroom. Mol. Plant Pathol. 2010, 11, 585–595. [Google Scholar] [CrossRef] [PubMed]
- Kettering, M.; Sterner, O.; Anke, T. Antibiotics in the chemical communication of fungi. Z. Naturforsch. 2004, 59, 816–823. [Google Scholar] [CrossRef] [PubMed]
- Bäuerle, J.; Anke, T.; Jente, R.; Bosold, F. Antibiotics from Basidiomycetes. XVI. Antimicrobial and cytotoxic polyines from Mycena viridimarginata Karst. Arch. Microbiol. 1982, 132, 194–196. [Google Scholar] [CrossRef]
- Celmer, W.D.; Solomons, I.A. The structure of the antibiotic mycomycin. J. Am. Chem. Soc. 1952, 74, 1870–1871. [Google Scholar] [CrossRef]
- Parish, C.A.; Huber, J.; Baxter, J.; González, A.; Collado, J.; Platas, G.; Diez, M.T.; Vicente, F.; Dorso, K.; Abruzzo, G.; et al. A new ene-triyne antibiotic from the fungus Baeospora myosura. J. Nat. Prod. 2004, 67, 1900–1902. [Google Scholar] [CrossRef] [PubMed]
- Kai, K.; Sogame, M.; Sakurai, F.; Nasu, N.; Fujita, M. Collimonins A-D, unstable polyynes with antifungal or pigmentation activities from the fungus-feeding bacterium Collimonas fungivorans Ter331. Org. Lett. 2018, 20, 3536–3540. [Google Scholar] [CrossRef]
- Pan, Y.; Lowary, T.D.; Tykwinski, R.R. Naturally occurring and synthetic polyyne glycosides. Can. J. Chem. 2009, 87, 1565–1582. [Google Scholar] [CrossRef]
- He, J.; Shen, Y.; Jiang, J.S.; Yang, Y.-N.; Feng, Z.-M.; Zhang, P.-C.; Yuan, S.-P.; Hou, Q. New polyacetylene glucosides from the florets of Carthamus tinctorius and their weak anti-inflammatory activities. Carbohydr. Res. 2011, 346, 1903–1908. [Google Scholar] [CrossRef]
- Konovalov, D.A. Polyacetylene compounds of plants of the Asteraceae family (review). Pharm. Chem. J. 2014, 48, 615–633. [Google Scholar] [CrossRef]
- Walsh, C.T.; Schonbrunn, A.; Lockridge, O.; Massey, V.; Abeles, R.H. Inactivation of a flavoprotein, lactate oxidase, by an acetylenic substance. J. Biol. 1972, 247, 6004–6006. [Google Scholar]
- Citores, L.; Ragucci, S.; Ferreras, J.M.; Di Maro, A.; Iglesias, R. Ageritin, a ribotoxin from poplar mushroom (Agrocybe aegerita) with defensive and antiproliferative activities. ACS Chem. Biol. 2019, 14, 1319–1327. [Google Scholar] [CrossRef] [PubMed]
- Dresch, P.; D’Aguanno, M.N.; Rosam, K.; Grienke, U.; Rollinger, J.M.; Peintner, U. Fungal strain matters: Colony growth and bioactivity of the European medicinal polypores Fomes fomentarius, Fomitopsis pinicola and Piptoporus betulinus. AMB Express 2015, 5, 4. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Hilgers, F.; Loeschke, A.; Jaeger, K.-E.; Feldbrügge, M. Ustilago maydis serves as a novel production host for the synthesis of plant and fungal sesquiterpenoids. Front. Microbiol. 2020, 11, 1655. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Extract | Species | Strain | Geographic 1 and Ecological Data |
|---|---|---|---|
| BE01 | Armillaria ostoyae | P089 | Göttingen (Gö), Kerstlingeröder Feld, under Pinus nigra, tc 2, leg., det. M.G. (2009-10-07) |
| BE02 | Armillaria cf. ostoyae | P145 | 1.2 km S of Grobsdorf (Gd), Nordhalde (Nh), lc 3, Tilia sp. stump, leg., det. M.G. (2010-09-27) |
| BE03 | Coprinopsis atramentaria | P125 | Jena (J), Beutenberg, half buried softwood, lc, leg., det. M.G. (2013-11-18) |
| BE04 | Coprinopsis picacea | P106 | 1 km W of Gö-Herberhausen (GöHe), Fagus sylvatica forest (Fsf), tc, leg., det. M.G. (2013-11-25) |
| BE05 | Fistulina hepatica | P052 | J, 1 km SW of “Steinkreuz Ziegenhain”, Fsf, lc, Quercus sp. stump, leg., det. F.H. (2013-08-18) |
| BE06 | Flammulina velutipes | P111 | J, Cospedaer Grund, lc, Fraxinus excelsior stump, leg., det. M.G. (2009-12-01) |
| BE07 | Fomes fomentarius | P116 | Jonsdorf, Weißer Stein, lc, Betula pendula wood, leg., det. M.G. (2012-03-05) |
| BE08 | Pleurotus dryinus | P090 | Dederstedt, lc, living stem of F. excelsior, leg., det. Dr. habil. H. Dörfelt (2009-11-18) |
| BE09 | Pleurotus cf. pulmonarius | P046 | Oberursel-Hohemark (OHm), mixed Fsf, lc, F. sylvatica wood, leg., det. F.H. (2013-09-13) |
| BE10 | Pleurotus ostreatus | P118 | Hannoversche Klippen 1.5 km NW of Bad Karlshafen, lc, F. sylvatica wood, leg., det. M.G. (2012-07-30) |
| BE11 | Agaricus arvensis | P151 | Kauern, Nh, 1 km S of Gd, mixed forest, tc 2, leg., det. M. G. (2009-09-23) |
| BE12 | Agaricus augustus | P096 | 2.5 km NE of Maria Laach (ML), mixed Fsf, tc, leg., det. M. G. (2010-08-12) |
| BE13 | Agaricus augustus | P148 | Gö., Brüder-Grimm-Allee, tc, under Tilia sp., leg., det. M. G. (2013-10-25) |
| BE14 | Auricularia auricula-judae | P093 | J, Mühltal, lc 3, Sambucus nigra wood, leg., det. M. G. (2009-12-07) |
| BE15 | Bovista nigrescens | P123 | Rothesütte, about 500 m SE of Rothesütte, tc, pasture, leg., det. M. G. (2013-09-23) |
| BE16 | Clitocybe geotropa | P149 | Gö, close to GöHe, Fsf, tc, leg., det. M. G. (2013-12-02) |
| BE17 | Clitocybe odora | P053 | Morgenröthe, mixed Picea abies forest, tc, needle litter, leg., det. F. H. (2013-09-07) |
| BE18 | Galerina marginata | P057 | OHm, mixed Fsf, lc, P. abies wood, leg., det. F. H. (2013-09-13) |
| BE19 | Ganoderma lucidum | P095 | Königsfeld/SW, 500 m N of Königsfeld, lc, Quercus sp. stump, leg., det. M. G. (2010-07-17) |
| BE20 | Gloeophyllum odoratum | P124 | J, Mühltal, lc, P. abies stump, leg., det. M. G. (2013-11-04) |
| BE21 | Hypholoma fasciculare | P099 | 750 m SE of Gd, mixed forest, lc, Quercus sp. stump, leg., det. M. G. (2010-09-07) |
| BE22 | Hypholoma sublateritium | P169 | 200 m S of Closewitz, Quercus spp. forest, lc, Quercus sp. stump, leg., det. M. G. (2014-02-17) |
| BE23 | Inonotus obliquus | P150 | Oybin, Töpfer/Brandhöhe, lc, B. pendula wood, leg., det. M. G. (2014-01-06) |
| BE24 | Kuehneromyces mutabilis | P050 | OHm, mixed Fsf, lc, on F. sylvatica wood, leg., det. F. H. (2013-09-13) |
| BE25 | Langermannia gigantea | P051 | Bad Berka, Trebestrasse, tc, garden lawn, leg., det. F. H. (2013-08-17) |
| BE26 | Lycoperdon excipuliforme | P122 | 800 m S of Gd, tc, under scattered B. pendula trees, leg., det. M. G. (2013-09-12) |
| BE27 | Lycoperdon molle | P154 | ML, Laacher-See-Haus, 1.3 km SE of ML, on a path in a Fsf, tc, leg., det. M. G. (2010-08-12) |
| BE28 | Macrolepiota procera | P097 | ML, 2.5 km NE of ML, mixed Fsf, tc, leg., det. M. G. (2010-08-16) |
| BE29 | Macrolepiota procera | P130 | Kauern, Nh, 1.2 km S of Gd, mixed forest, tc, leg., det. M. G. (2009-09-25) |
| BE30 | Marasmius oreades | P144 | 1 km S of Gd, tc, grassy forest margin, leg., det. M. G. (2010-09-08) |
| BE31 | Marasmius scorodonius | P055 | OHm, mixed Fsf, tc, F. sylvatica leaf litter, leg., det. F. H. (2013-09-13) |
| BE32 | Megacollybia platyphylla | P026 | 2.5 km N of Rambach, 2 km NW of Naurod, mixed Fsf, tc, leg., det. F. H. (2013-05-27) |
| BE33 | Pleurotus ostreatus | P114 | ML, Laacher-See-Haus, 1.3 km SE of ML, lc, on F. sylvatica wood, leg., det. M. G. (2011-08-02) |
| BE34 | Pleurotus ostreatus | P119 | Rosdorf, Kiessee, lc, on wood of Salix sp., leg., det. M. G. (2013-08-23) |
| BE35 | Pleurotus ostreatus | P128 | Hainewalde, near graveyard, lc, Populus x canadensis stump, leg., det. M. G. (2014-01-06) |
| BE36 | Polyporus brumalis | P117 | 1.5 km SW of GöHe, mixed Fsf, lc, Tilia sp. wood, leg., det. M. G. (2012-03-05) |
| BE37 | Polyporus tuberaster | P036 | J, 300 m S of “Steinkreuz Ziegenhain”, Fsf, lc, on decayed wood, leg., det. F. H. (2013-08-18) |
| BE38 | Sarcomyxa serotina | P126 | 800 m W of GöHe, mixed Fsf, lc, F. sylvatica stump, leg., det. M. G. (2013-11-25) |
| BE39 | Stropharia aeruginosa | P102 | Jenaprießnitz, Tännicht, Fsf, tc, leg., det. M. G. (2010-10-06) |
| BE40 | Trametes gibbosa | P129 | J, Mühltal, 1.3 km SW of Cospeda, F, sylvatica wood, leg., det. M. G. (2013-03-17) |
| BE41 | Tricholomopsis rutilans | P056 | OHm, mixed Fsf, lc, F. sylvatica wood, leg., det. F. H. (2013-09-13) |
| No. | 1H (mult., J) | 13C, mult. |
|---|---|---|
| 1 | 0.95 (t, 6.8) | 8.6, CH3 |
| 2 | 1.53 (m) | 29.3, CH2 |
| 3 | 4.06 (td, 7.1, 1.5) | 72.4, CH |
| 4 | 6.32 (15.9, 5.6) | 149.8, CH |
| 5 | 5.78 (ddd, 15.9, 1.7, 0.7) | 107.3, CH |
| 6 | - | 76.0, C |
| 7 | - | 72.8, C |
| 8 | - | 68.0, C |
| 9 | - | 83.2, C |
| 10 | 4.41 (t, 6.7) | 61.5, CH |
| 11 | 1.68 (m) | 39.5, CH2 |
| 12 | 1.48 (m) | 18.1, CH2 |
| 13 | 0.98 (t, 6.7) | 12.6, CH3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, J.; Shi, Y.-M.; Grün, P.; Gube, M.; Feldbrügge, M.; Bode, H.; Hennicke, F. Identification of Feldin, an Antifungal Polyyne from the Beefsteak Fungus Fistulina hepatica. Biomolecules 2020, 10, 1502. https://doi.org/10.3390/biom10111502
Lee J, Shi Y-M, Grün P, Gube M, Feldbrügge M, Bode H, Hennicke F. Identification of Feldin, an Antifungal Polyyne from the Beefsteak Fungus Fistulina hepatica. Biomolecules. 2020; 10(11):1502. https://doi.org/10.3390/biom10111502
Chicago/Turabian StyleLee, Jungho, Yi-Ming Shi, Peter Grün, Matthias Gube, Michael Feldbrügge, Helge Bode, and Florian Hennicke. 2020. "Identification of Feldin, an Antifungal Polyyne from the Beefsteak Fungus Fistulina hepatica" Biomolecules 10, no. 11: 1502. https://doi.org/10.3390/biom10111502
APA StyleLee, J., Shi, Y.-M., Grün, P., Gube, M., Feldbrügge, M., Bode, H., & Hennicke, F. (2020). Identification of Feldin, an Antifungal Polyyne from the Beefsteak Fungus Fistulina hepatica. Biomolecules, 10(11), 1502. https://doi.org/10.3390/biom10111502