Omics-Based Platforms: Current Status and Potential Use for Cholangiocarcinoma



1
Department of Biomedical Imaging and Radiological Sciences, National Yang-Ming University, Taipei 112, Taiwan
2
Genomics Research Center, Academia Sinica, 128 Academia Rd., Sec. 2, Nankang-Dist., Taipei 115, Taiwan
3
School of Medicine, National Yang-Ming University, Taipei 112, Taiwan
4
Center of Immuno-Oncology, Department of Oncology, Taipei Veterans General Hospital, Taipei 112, Taiwan
5
Department of General Surgery, Liver Research Center, Chang Gung Memorial Hospital, Linkou, Taoyuan 333, Taiwan
6
Cancer Genome Research Center, Chang Gung Memorial Hospital, Linkou, Taoyuan 333, Taiwan
*
Author to whom correspondence should be addressed.
Biomolecules 2020, 10(10), 1377; https://doi.org/10.3390/biom10101377
Submission received: 8 September 2020
/
Revised: 26 September 2020
/
Accepted: 26 September 2020
/
Published: 28 September 2020
(This article belongs to the Special Issue Cutting Edge Research for Exploration of Biomolecules for Gemcitabine-Based Chemo-Resistant Advanced Bile Duct Cancer: from Basic Study to Clinical Trial)
Abstract
:Cholangiocarcinoma (CCA) has been identified as a highly malignant cancer that can be transformed from epithelial cells of the bile duct, including intrahepatic, perihilar and extrahepatic. High-resolution imaging tools (abdominal ultrasound, computed tomography and percutaneous transhepatic cholangial drainage) are recruited for diagnosis. However, the lack of early diagnostic biomarkers and treatment evaluation can lead to serious outcomes and poor prognosis (i.e., CA19-9, MUC5AC). In recent years, scientists have established a large number of omics profiles to reveal underlying mechanisms and networks (i.e., IL-6/STAT3, NOTCH). With these results, we achieved several genomic alteration events (i.e., TP53mut, KRASmut) and epigenetic modifications (i.e., DNA methylation, histone modification) in CCA cells and clinical patients. Moreover, we reviewed candidate gene (such as NF-kB, YAP1) that drive gene transcription factors and canonical pathways through transcriptomics profiles (including microarrays and next-generation sequencing). In addition, the proteomics database also indicates which molecules and their directly binding status could trigger dysfunction signatures in tumorigenesis (carbohydrate antigen 19-9, mucins). Most importantly, we collected metabolomics datasets and pivotal metabolites. These results reflect the pharmacotherapeutic options and evaluate pharmacokinetic/pharmacodynamics in vitro and in vivo. We reversed the panels and selected many potentially small compounds from the connectivity map and L1000CDS2 system. In this paper, we summarize the prognostic value of each candidate gene and correlate this information with clinical events in CCA. This review can serve as a reference for further research to clearly investigate the complex characteristics of CCA, which may lead to better prognosis, drug repurposing and treatment strategies.
1. Introduction to CCA
Cholangiocarcinoma (CCA) is a malignant tumor transformed from bile duct epithelial cells [1,2]. According to the anatomic location, it is divided into three different subtypes—intrahepatic (iCCA, 10–20%), perihilar (pCCA, 50–60%) and extrahepatic (eCCA, 20–30%) [3]. iCCA is the second most common primary liver cancer after hepatocellular carcinoma. Recently, iCCA was divided into two subtypes (small- and large-duct types) based on the production of mucin and phenotype by immunohistochemically evaluated. The small-duct-type cancer usually has a history of chronic liver disease (five-year overall survival is longer ~60%, most of which are genetic changes of IDH1, BAP1 and FGFR2) and large-duct iCCA and pCCA have many clinicopathological characteristics and are accompanied by premalignant biliary intraepithelial neoplasia (BiIIN) in adjacent ducts (five-year overall survival is shorter, accounting for about 20%, most of which are genetic changes of SMAD4 and KRAS7) [4]. The incidence of CCA has been increasing worldwide, especially in Asia [5]. Analyzing the annual mortality rate of CCA, the average mortality rate for the period 2010–2014 was 0.3–2.88 (per 1 × 105 inhabitants) [6]. However, higher mortality rates (>4, per 1 × 105 inhabitants) including China [7], Japan [8], South Korea [9], Thailand [10], Taiwan [11] and Austria [12]. It has also been confirmed that men have a higher odds ratio than women [13]. Surgical resection is the only potentially curative clinical therapy and results in a five-year overall survival rate between 15–40% [14,15,16,17,18]. More than two-thirds of patients relapse after resection. The main reasons for the high recurrence rate are positive surgical margins, lymph node metastasis, vascular infiltration and multiple tumor regions.
Recent reports on CCA have identified several potential risk factors and carcinogens. Biliary tract infections caused by parasitic infections can cause liver flukes (Clonorchis sinensis and Opisthorchis viverrini) and they have the highest incidence of CCA in Northeastern Thailand [19]. Primary sclerosing cholangitis, hepatitis B/C virus (HBV/HCV) infection, biliary stone diseases, congenital biliary malformations, cirrhosis and exposure to aromatic toxins are all potential CCA disease etiologies [20]. In addition, smoking, alcohol consumption and diabetes also contribute to the risks of CCA. In particular, 1,2-dichloropropane (1,2-DCP) has been demonstrated as a carcinogen for CCA. The main role of 1,2-DCP is interference with CYP450-dependent proliferation and apoptosis [21]. For treatment, palliative chemotherapy with gemcitabine and cisplatin is the standard 1st line treatment for unresectable or relapse/recurrence CCA patients. However, the response to such therapy remains limited [22]. Most importantly, CCA can escape several known tolerance mechanisms induced by chemotherapy (including cisplatin, gemcitabine and other combinations) [23]. Drug development that relies on well-known signal transductions needs in-depth discussion.
Compared with known pathogenic factors and diagnosis protocols, the oncogenic mechanisms and treatment strategies still need to be improved and investigated [24]. Therefore, this review summarizes these omics-based datasets. These comprehensive approaches aim to solve the bottleneck of CCA study.
2. Genomics Alterations and Epigenetic Changes in CCA
In recent years, 38 CCA tumors have been added to The Cancer Genome Atlas (TCGA). Through these changes, we can achieve alterations of tumor suppressor genes and oncogenes. TCGA has established pipelines through the following platforms: whole exome sequencing, affymetrix SNP 6.0 copy number array, RNA sequencing (RNA-seq) containing microRNA/long noncoding RNA, DNA methylation and reverse-phase protein array (RPPA). Combining this evidence, scientists have revealed genetic alterations and epigenetic events in CCA [25,26].
2.1. Genomic Alterations
The genome sequencing study conducted by Nakamura et al. in CCA identified many driver gene changes. A total of 239 pairs of CCA and corresponding normal tissues (including 137 iCCA, 74 eCCA and 28 gallbladder cancers) were sequenced by whole exons [27]. The results show that FDFR2 was specifically fused with iCCA (6/109, 55%), and the two FGFR2 fusion proteins can induce ligand-independent FGFR2 autophosphorylation and downstream mitogen-activated protein kinase signaling (MAPK). After activation, anchorage-independent growth and tumor formation can be increased. On the other hand, in eCCA cases, cyclic AMP (cAMP)-dependent protein kinase (PKA) signaling components can form new fusions ATP1B-PRKACA and ATP1B-PRKACB (25/260, 9.6%). These fusions also increase the PKA activity and activate MAPK signaling [28]. Aside from somatic alterations, they also identified several driver gene landscapes in CCA. Sixteen putative tumor-suppressor genes have been recognized, including TP53, ARID1A, BAP1, ARID2, PBRM1, APC, EPHA2, ELF3, ATM, BRCA2, RPL22, ACVR2A, STK11, MLL3 (KMT2C), NF1 and MLL2 (KMT2D). After annotation and validation, they finally announced that the key factors in CCA include kinases (FGFR1, FGFR2, FGFR3, PIK3CA, ALK, EGFR, ERBB2, BRAF and AKT3), oncogenes (IDH1, IDH2, CCND1, CCND3 and MDM2) and tumor suppressor genes (BRCA1 and BRCA2) [27]. Tian et al. also identified a comprehensive genomic profile of CCA in China’s population. In this study, they inputted 44 iCCA and 22 eCCA cases. The results show: TP53 (62.1%), KRAS (36.4%), SMAD4 (24.2%), TERT (21.2%), ARID1A (19.7%), CDKN2A (19.7%), KMT2C (9.1%), RBM10 (9.1%), ERBB2 (7.6%) and BRAF (7.6%). Similarly, they observed specificities expressed in iCCA (STK11, CCND1, FGF19, FGF3, FGF4 and PBRM1). For FGFR2, they are expressed in both Western and Chinese cohorts [7]. Through the cBioPortal website [29,30], we merged six clinical cohorts to analyze their genetic alteration events, survival time and the proportion of several demographic factors (Figure 1). We also summarized the top frequency copy-number alteration (CNA)/mutation/fusion gene, as shown in Table 1. In addition, we performed a graph to indicate the multiple genetic alteration events in above studies (Figure 2).
2.2. Epigenetic Events
In an epigenetic study, there are often several cell cycle-related factors and genes involved in DNA repair functions that are frequently methylated in CCA. Therefore, dysfunction of these genes may suppress tumor suppression and unusual cell behavior. MLH1, DCLK1, CDO1, ZSCAN18, ZFG331, p14, p16, DAPK, CCND2, CDH13, GRIN2B, RUNX3, TWIST1, EGFR and LKB1 are most of all CpG sites that have exhibited hypermethylation and then loss of function in CCA tumorigenesis [31,32,33,34,35,36,37,38,39,40]. Moreover, RASSF1A, FHIT, 14-3-3, TMS1, CDH1, CHFR, GSTP, TIMP3, SEMA3B, RAR-β, MGMT, BLU, THBS1, RIZ1, OPCML and PTGS2 have also been mentioned in CCA subtypes or cell lines. In histone modification, CCA studies have shown that histone methylation, acetylation and chromatin remodeling are unbalanced. H3K9, H3K27 and H4K20 methylation leads to transcriptional repression, while H3K4 methylation leads to transcriptional activation [41]. In an in vivo CRISPR model, mutation of Arid1a leads to SWI/SNF chromatin complex remodeling. Moreover, both of the SWI/SNF family members, AT-rich interaction domain (ARID) 1A and polybromo 1 (PBRM1) showed loss of function in CCA tumorigenesis [42,43]. Non-synonymous mutations in chromatin remodelers are typically distributed throughout the gene. There are no definable hotspots, and the significance of many mutations is unknown. Therefore, histone modification and chromatin remodeling still require in-depth investigation and innovative research.
An important aspect of epigenomics is microRNAs (miRNAs). Deregulation of miRNAs can induce the initiation and progression of cancers by modifying target tumor suppressor genes or oncogenes. Recently, it has been reported that IL-6 regulates the activity of DNA methyltransferase 1 (DNMT1) by miRNAs in CCA cells [44]. DNMT1 can cause comprehensive changes in the degree of methylation. In CCA, various genes have been reported to have frequent methylation, including MLH1, DCLK1, CDO1, ZSCAN18, ZNF331, p14, p16, DAPK, CCND2, CDH13, GRIN2B, RUNX3, TWIST1, EGFR and LKB1 [45]. After verification, it was confirmed that the miR-148a/miR-152 family directly binds to the DNMT gene. Interestingly, DNMT1 regulates the hypermethylation of its CpG islands and then loses these tumor suppressor functions. Therefore, they form a negative feedback regulatory loop between DNMT1 and miR-148a/miR-152 family in tumorigenesis. Moreover, several tumor suppressor miRNAs have been shown in the CCA model. miR-370 was inhibited following global hypermethylation. Then, downstream target mitogen-activated protein kinase 8 (MAP3K8) was regulated by miR-370 [46]. Another study also showed that miR-376c was regulated by DNA methylation and associated with tumor suppression by targeting growth factor receptor-bound protein 2 (GRB2) [47]. Yang et al. observed that miR-144 was reduced in CCA tissues. They also verified platelet-activating factor acetylhydrolase isoform 1b (LIS1) as the direct target in CCA [48]. Although the target gene of direct action of some miRNAs has not yet been determined, through statistics and calculations, we list the most significant changes of miRNAs in CCA, especially between normal adjacent tissues and tumor groups. These miRNAs include oncomiRs (miR-183, miR-96, miR-182 and miR-181b) and tumor suppressor miRs (miR-99a, miR-125b-2, miR-621, miR-551b, miR-378c, miR-148a, miR-139, miR-378, miR-483, miR-885, miR-122, miR-490, miR-675 and miR-1258). Although many have not been examined and studied, these candidates have the potential for follow-up research. We drew a heat-map from the OncoMir Cancer Database (OMCD) to illustrate the expression of the most significantly different miRNAs between normal adjacent tissues and tumor sites according to appropriate criteria (Figure 3 and Table 2) [49]. Regarding the regulation of non-coding genes, not only mentioned the expression of microRNA in CCA, but also mentioned long-noncoding RNA (LncRNAs) and circular RNA (circRNAs) as contributors to the CCA model (Table 3). Zhang et al. revealed that LncRNA–colon cancer-associated transcript 1 (CCAT1) increased in tumor parts, and their result show that knockdown of CCAT1 suppressed the migration and invasion of iCCA cells. Then, they demonstrated the epithelial–mesenchymal transition (EMT) activation through the CCAT1-miR-152 axis in CCA [50]. Moreover, Xu et al. observed that CCAT2 had a similar function to CCAT1 in CCA. Overexpression of CCAT family members promotes CCA cell migration and invasion by EMT induction [51]. In addition, miR-590-3p was also revealed to suppress the EMT procedure through inhibition of the ZEB2 gene in iCCA [52]. On the other hand, LncRNA H19 (chromosome11p15.5) is also involved in cell proliferation, apoptosis and migration/invasion through EMT [53]. Li et al. also showed that lncRNA SOX2ot (SOX2 overlapping transcript) promotes cell proliferation and invasion in CCA cells [54]. Gao et al. confirmed that miR-10a-5p is one of the targets of PTEN and the miR-10a-5p inhibitor can suppress the PTEN expression and decrease inhibited Akt phosphorylation status in CCA cells. Therefore, miR-10a-5p regulates cholangiocarcinoma cell growth through the Akt pathway [55]. Similarly, more candidate miRNAs were reported over five years ago. Therefore, we do not mention them in detail in this review.
3. Canonical Pathways and Key Drivers of the Transcriptome
Several driver genes of CCA have been revealed through next-generation sequencing and their transcriptome datasets. Li et al. announced that Forkhead box M1 (FOXM1)/NF-kB has an inverse correlation between methionine adenosyltransferase 1(MAT1A) in both hepatocellular carcinoma and CCA (Pearson’s rho = −0.36, p = 1.24 × 10−190) [56]. MAT1A can mainly synthesize S- adenosylmethionine (SAMe) levels, which can prevent cholestatic liver injury. MAT1A has been identified that the expression level of MAT1A in primary and metastatic CCA is reduced compared with normal bile duct cells. Among them, MAT1A exerted a tumor suppressor effect and can also be used as an independent prognostic factor (Cox coefficient = −0.258, p = 6.8 × 10−3). In addition, prohibitin 1 (PHB1) plays a tumor-suppressive role and positively regulates MAT1A while suppressing c-Myc [57]. NAD(P)H-quinone oxidoreductase 1 (NQO1) plays an important role in chemo-resistance (5-fluorouracil and doxorubicin) and proliferation in several CCA cell lines. NQO1 enzyme activity can prevent potent cytotoxicity of chemotherapy drugs and eliminate cell apoptosis caused by caspase 3/7. [58]. Yeh et al. also claimed that TNNI3K has been noticed in array comparative genomic hybridization (aCGH) along with its overexpression in CCA cells and patients. TNNI3K is highly expressed and enhances cell growth and colony forming ability in vitro. [59]. Moreover, carbohydrate antigen 19-9 (CA19-9) is recognized as a well-known biomarker in pancreatic cancer and CCA [60]. In addition to internal genetic mutations and aberrant expression, external pathogens are also possible carcinogenic mechanisms. Opisthorchis viverrini is a specific pathogen of CCA. Aksorn et al. also recognized novel potential biomarkers that are associated with these events in CCA. These candidates include immunoglobulin heavy chain, translocated in liposarcoma (TLS) and visual system homeobox 2 (VSX2) [61]. There are many typical and classic markers in CCA, including KRAS, TP53, PROM1, CTGF, VIM, DKK1, SOX2, SOX17, MUC1, PTEN, PTPN14, c-Met, EGFR, VEGR, CD44 and CDH1 [62,63,64,65,66,67,68,69,70,71,72], but this review specifically focuses on novel evidence from the past five years.
4. Protein Levels and Post-Translational Modifications in CCA
Not only DNA status and RNA expression level change in CCA, but several protein molecules have also been raised in recent studies. For comprehensive and large-scale screening, scientists established proteomics-based approaches to address these unsolved issues. Liquid chromatography/mass spectrometry was used to examine the differential abundance of protein molecules in CCA. Myeloperoxidase (MPO), complement C3, inter-alpha-trypsin inhibitor heavy chain H4 (ITIH4), apolipoprotein B-100 and kininogen-1 isoform 2 (KNG1) increased in the tumor group compared to the benign group. On the other hand, trefoil factor 2, superoxide dismutase (SOD), kallikrein-1 (KLK1), carboxypeptidase B (CPDB) and trefoil factor 1 decreased in tumors [73]. Zhang et al. utilized two-dimensional difference in gel electrophoresis (2D-DIGE) combined with MALDI-TOF to identify mice exposed to 1,2-dichloropropane (1,2-DCP). The mice were exposed to 0, 50 and 250 ppm of 1,2-DCP through an inhalation exposure system (8 h/d, 7 d/week for four weeks) and the frozen livers was collected for further lysis and analysis. They captured several components include ferritin heavy chain (FTH1) (upregulated), ferritin light chain (FTL1) (upregulated), acyl-coenzyme a thioesterase 2 (ACOT2) (upregulated), GSTM1 (upregulated), aldehyde dehydrogenase X, mitochondrial (ALDH1B1), formimidoyltransferase cyclodeaminase (FTCD) (downregulated) and selenium-binding protein 2 (SELENBP2) (downregulated) [74]. Son et al. focused on eCCA specimens and collected bile samples from 18 patients with eCCA and the five patients with benign conditions for control group. By liquid chromatography–mass spectrometry (LC–MS) identified immunoglobulin kappa light chain, apolipoprotein E (APOE), albumin (ALB), apolipoprotein A-I (APOA1), antithrombin-III (SERPINC1), α1-antitrypsin (SERPINA1), serotransferrin (TF), immunoglobulin heavy constant mu (IGHM), immunoglobulin J chain (JCHAIN), complement C4-A (C4A) and complement C3 are all overexpressed compared with control groups [75].
There are several molecules with secretory ability, and it has recently been reported that various circulating RNAs and genes have been measured in serum. In the same situation, the establishment of secretome datasets also confirms that the extracellular molecules facilitate in CCA. These markers are also developing as prognostic/diagnostic factors in clinical study. Large et al. demonstrated the co-expression of plasma thrombospondin-2 (THBS2) with CA19-9 in CCA patients [76]. Furthermore, MMP7, IL-6, S100A6, DKK1 and SSP411 are also abnormally expressed proteins in CCA patients’ serum [70,77,78,79,80]. It is worth mentioning that Wang et al. declared that N-glycan status, including peak10 and NA3F2, presents a better diagnostic value than typical marker CA19-9 in eCCA patients’ serum [81].
5. Available Omics Datasets for CCA
Omics datasets have been applied in basic research studies. Da et al. merged the Gene Expression Omnibus (GEO) database GSE32225 with quantitative isobaric tags for relative and absolute quantification (iTRAQ) proteomics approaches to evaluate multiple candidate proteins associated with prognosis of CCA patients. In this study, they screened three candidate proteins APOF, ITGAV and CASK. They detected and scored the protein levels of all molecules on the CCA tissue microarray through IHC analysis. After comparison and statistical evaluation, they confirmed that peripheral plasma membrane protein CASK is considered the most significant molecule in this study [82]. The level of CASK expression has been determined to be correlated with overall survival (HR = 2.031, p = 0.027), vascular invasion (p = 0.009) and T classification (p = 0.017). Sulpice et al. established microarray chips (GSE45001) to predict that osteopontin is the independent predictor in iCCA and also performed gene expression profiling of tumor microenvironment in same cohort [83,84]. Peraldo Neia et al. revealed the gene and microRNA pattern of the iCCA patient-derived xenograft (PDX) model (GSE84918/GSE84938) [85]. Varamo analyzed the specific signatures to indicate the gemcitabine resistance CCA cell lines (GSE116118) [23]. Long-term exposure of gemcitabine-resistance cell lines (>1.5 uM, nine months) compared with sensible counterpart cells. They observed that several drug resistance- (ESR2 and FGF2) and EMT-related targets (COL5A2, RGS2, SNAI2, SPP1, TFPI2, TGFB1 and VIM) were regulated in the event. They further predicted several molecular mechanisms through 354 downregulated and 382 upregulated targets. These typical pathways include DNA replication, cell cycle and pyrimidine metabolism. In addition, they also found new molecules with differently expressed in their model (QPRT, RRM1, FOSB, PRSS1, etc.) [23]. Moreover, Gu et al. tried to find the linkage between microcystin–leucine–arginine (MC-LR) and iCCA (GSE140687). MC-LR has recognized that certain hepatotoxins can induce liver damage and liver cancer. MC-LR is also correlated with intrahepatic bile duct hyperplasia. Therefore, the group tried to explore the relationship between MC-LR, inflammation and intrahepatic bile duct epithelial cell proliferation. Although they have not yet come to the results of MC-LR in iCCA. However, they have established a microarray profile containing blank and MC-LR after 24 h of treatment in Huh28 cells. Meijer distinguished the circulating miR-16 and miR-877 as the diagnostic classifiers for eCCA (GSE117687) [86].
Recently, next-generation sequencing has been widely used. Yan et al. sequenced the single cell of iCCA and found intercellular crosstalk between iCCA cells and vascular cancer-associated fibroblasts (vCAFs) (GSE148773/GSE142784/GSE138709) [87]. They found that the IL-6/IL-6R axis was enriched in the interplay between vCAFs and iCCA cells. vCAFs contribute to the tumorigenesis and cancer stemness ability of iCCA through the axis. On the other hand, tumor exosomal miR-9-5p elicits IL-6 expression in vCAFs, which leads to iCCA epigenetic alterations. Saengboonmee et al. investigated the CCA cell lines in standard and high glucose medium (GSE137803). High glucose medium conditions promoted proliferation, metastatic and phosphorylation/ nuclear translocation of STAT3 [88]. Ma et al. claimed that HCC and iCCA have a varying degree of transcriptomics diversity through single-cell transcriptome profiling (GSE125449) [89]. Zhou et al. are performing ongoing non-coding RNA profiling to catalog the expression profiles of 1643 microRNAs in each sample (GSE93366). To determine the difference between serum and urine in CCA patients, Lapitz et al. established the array chips through a liquid biopsy approach (GSE144521) [90].
Many scientists have explored the aforementioned genetic alteration and epigenetic modification using omics-based technology. Bledea et al. dissected methylation data from IDH mutant CCA (GSE124617) [91], and their results support the suggestion that hypermethylation of enhancers associated with cell differentiation may mediate lack of differentiation in IDH1/2 mutant cancers. The profile of PTEN overexpression in CCA also has been checked by transcriptome array (GSE146196). A study classified their molecular panels and re-evaluated four classes for eCCA (KRAS: 36.7%, TP53:34.7%, ARID1A: 14% and SMAD4:10.7%) (GSE132305) [92]. All GSE datasets have been rationally designed, with appropriate criteria and comprehensively analyzed. Therefore, this information will become the foundation for future research.
6. Metabolomics Analysis of Oncometabolites and Metabolic Reprogramming in CCA
Isocitrate dehydrogenase 1 and 2 (IDH1 and IDH2) play a pivotal role in both DNA methylation and histone modification in CCA. IDH1 and IDH2 mutants occur only in ICC (10–28%). Wild-type IDH1 and IDH2 convert isocitrate to α-ketoglutarate (α-KG) in the cytoplasm and mitochondria, respectively. In contrast, mutant isoforms can produce anabolic 2-hydroxyglutarate (2-HG) [93,94]. As a result, many biologic processes that rely on α-KG, including ten-eleven translocase (TET) 2-mediated DNA demethylation, are disrupted. This leads to a wide range of epigenomic consequences, including increased 5-methylcytosine (5mC) content, decreased 5-hydroxymethylcytosine (5hmC) and increased histone H3 lysine 79 (H3K79) demethylation [95].
Liang et al. analyzed concentrated secretion ability through serum metabolomics establishment, and it was identified that the contents of LysoPC (14:0, C22H46NO7P) and LysoPC (15:0, C23H48NO7P) in the CCA group were reduced compared with the control groups [96]. Alsaleh et al. confirmed that lipid-derived compounds (phospholipids, bile acids and steroids) and amino acid products (phenylalanine) are associated with the risk of CCA in the United Kingdom [97]. The reduction of aberrant bile phospholipid secretion will cause the formation of “toxic bile” and exacerbate inflammatory bile duct damage. Chronic cholestasis and cholangitis may further lead to secondary cirrhosis of the liver and are susceptible to hepatobiliary cancer. In addition, this group also identified that purine metabolism and lipid metabolism have changed in the CCA urine metabolome in Opisthorchis viverrini-induced liver fluke-associated CCA [98]. Several comprehensive analyses merged with transcriptome, proteome and metabolome profiles of CCA have been published [99,100,101,102,103]. Combined with artificial intelligence, Urman et al. analyzed the metabolomics and proteomics analysis of bile from patients with benign and malignant tumors. Their results show that phosphatidylcholines (PCs), sphingomyelins (SMs) and ceramides (Cer) were found in serum and tissue through machine learning [104]. These metabolites and canonical metabolism signatures are all important indicators for detecting and evaluating diagnosis/prognosis/cytotoxicity.
7. Using Multiple-Omics Approaches to Determine Prognostic Factors
Zhou et al. detected that miR-378 is significantly associated with the TNM classification of malignant tumors (TNM) stage (p = 0.03) and lymph node metastasis (p = 0.018). Their results also reveal that miR-378 can serve as an independent prognostic predictor for CCA patients (Hazard Ratio (HR) = 1.735, 1.007–2.988, p = 0.041) [105]. In the CCA serum, Liu et al. also found that circulating miR-21 is a promising biomarker for diagnosis and a prognostic biomarker of CCA [106]. In serum of CCA patients, Silakit et al. also provided evidence that miR-192 expression level correlated with lymph node metastasis (p = 0.047) and poor survival (HR = 2.076, 1.004–4.291, p = 0.049) [107]. Wang et al. also described miR-26 in serum as a diagnostic and prognostic marker for CCA patients (overall survival p = 0.018 and progression-free survival p = 0.0381) [108].
There is an inverse correlation between suppressor of cytokine signaling 3 (SOC3) and tumor necrosis factor α-induced protein 3 (TNFAIP3) in CCA. Furthermore, patients with low SOC3 expression or high TNFAIP3 expression showed a dramatically lower overall survival rate. The SOC3 and TNFAIP3 expression levels had a significant prognostic power for CCA survival rate (SOC3: HR = 2.382, 1.253–4.722, p = 0.022, TNFAIP3: HR = 6.598, 2.584–14.261, p = 0.009) [109]. It has been reported that chitinase 3-like 1 (CHI3L1) glycoprotein is dysfunctional in CCA. Plasma CHI3L1 expression was particularly associated with poor survival in CCA patients (HR = 2.117, 1.027–4.363, p = 0.038, HR = 1.642, 0.780–3.455, p = 0.192) [110]. More important, we summarized the genes most related to survival in CCA (top 10) from the gene expression profiling interactive analysis (GEPIA) website [111]. Interestingly, we observed that no genes play a key role in overall survival and disease-free survival. These results may be divided into different systems due to proliferation/tumorigenesis and recurrence/metastasis in CCA (Table 4).
In our review, we have preliminarily sorted out the significant differences calculated on various analysis platforms in CCA based on statistical formulas, but these analyses are carried out in low-dimensional areas. The latest variable selection approach integrating multilevel omics data. Through horizontal/vertical integration model and various algorisms (penalized, Bayesian, etc.), we can obtain multidisciplinary data as the focus of future research [112,113]. These models will calculate and evaluate some of the above targets and combined with actual in vitro and in vivo studies to determine their authenticity. At current stage, we first collected significantly different genes in CCA through statistical models and predicted their participation in the interactions and networks by bioinformatics software (Figure 4).
8. Potential Small Molecule Compounds and Clinical Drugs from Omics Datasets
8.1. FGFR Family
Based on previous study, several specific inhibitors and small compounds were developed to combat aggressive features in CCA. Selective/non-selective small molecule kinase inhibitors were designed for FGFR gene fusion. Pemazyre (pemigatinib), which targets to fibroblast growth factor receptor 2 (FGFR2) rearrangement or fusion, was developed. This agent was proved to be effective by a multicenter, open-labeled single-arm trial in advanced cholangiocarcinoma patients and got FDA-approval recently [114]. Infigratinib passed Phase I and II studies of advanced CCA with FGFR2 gene fusion or other FGFR genetic alterations that failed platinum-based chemotherapy (NCT02150967) [115]. Erdafitinib (BALVERSA) is an oral pan-FGFR selective small molecule kinase inhibitor and the FDA granted accelerated approval for patients with locally advanced or metastatic urothelial carcinoma, with susceptible FGFR3 or FGFR2 genetic alterations. The initial reports of CCA pts with FGFR alterations treated with erdafitinib had encouraging efficacy and acceptable safety profile. (NCT02699606). ARQ-087 (derazantinib) is a small molecule kinase inhibitor with ATP competitiveness that can block the proliferative activity of cell lines with FGFR aberrations [116]. Its related Phase I/II trial also focused on iCCA patients with FGFR2 fusion (NCT01752920). Moreover, TAS-120 [117], AZD4547 [118] and CH5183284 (Debio-1347) [119] are undergoing early clinical trials for FGFR-driven tumors (NCT02052778, NCT01948297, respectively). Non-selective FGFR inhibitors ponatinib or pazopanib have also been used in specific populations (NCT02265341). In the case series study, ponatinib has been shown to reduce carbohydrate antigen 19-9 and decrease lymph node metastasis in chemotherapy-refractory CCA patients [120]. Finally, FGFR monoclonal antibodies have fewer off-target effects through competitive binding to FGFR2b and were evaluated in a Phase I trial (NCT02318329).
8.2. IDH Mutants
IDH1/IDH2-specific inhibitors are also important issues in the treatment of CCA. IDH mutations cause metabolic reprogramming and the production of oncometabolites. Epigenetic platforms are also affected by IDH mutation status. Therefore, AGI-5198 and AGI-6780 are divided into selective IDH1 and IDH2 inhibitors, respectively. In clinical trials (NCT02073994, NCT02273739), AG-120 (Ivosidenib) and AG-221 (enasidenib) were used as next-generation orally selective inhibitors (IDH1, IDH2, respectively). More important, it was determined that they can inhibit the production of oncometabolite 2-HG and impair the growth of IDH mutant cells.
8.3. Others
Through omics-based approaches, various genetic changes, oncogenic pathways and oncometabolites have been identified in CCA. Thus, more functional inhibitors have been designed for tumorigenesis: (1) The PKA inhibitor isoquinoline H89 can inhibit CCA cell proliferation by regulating the expression levels of PRKACA and PRKACB [121]; (2) S63845 and AZD1480 can regulate myeloid cell leukemia sequence 1 (Mcl-1), Janus kinase (JAK) and the signal transducer and activator of transcription (STAT) signaling cascade [122,123]; (3) KRAS is a typical mutant marker in CCA. However, there are no effective direct inhibitors of KRAS mutants and downstream pathways. In a clinical study, selumetinib (selective MEK 1/2 inhibitor) has a synergistic effect with gemcitabine and cisplatin in advanced CCA and ongoing clinical trials (NCT02042443, NCT01438554) [124]; (4) The combined treatment of CCA includes the PI3K/AKT/mTOR pathway (BKM120), the RAF–MEK–ERK axis with gemcitabine and platinum-based drugs [125]; (5) Tissue with abnormal expression of mesothelin in several malignant tumors has been observed. Amatuximab is a chimeric monoclonal antimesothelin antibody, while SS1P is a recombinant anti-mesothelin immunotoxin, and they are all involved in clinical trials of multiple cancer types [126]; (6) DNMT inhibitors azacitidine and decitabine and HDAC inhibitors vorinostat and romidepsin through rearrangement of chromatin remodeling form an epigenetic platform to rescue the function of tumor suppressor genes [127,128,129]; (7) The MET tyrosine kinase inhibitors tivantinib and cabozantinib were selected as chemotherapy strategies in clinical trials (NCT01954745) [130]; (8) reforafenib inhibited the Raf/Erk/Elk-1 pathway, and MALT1 has been identified as having a pivotal role in CCA tumorigenesis. Moreover, regorafenib has been recognized as the second standard treatment option in metastatic colon cancer. The clinical trial in CCA is currently at Phase II-III [131,132,133].
8.4. Immune Checkpoint Blockage
Immunotherapy was regarded as the next-generation strategy for cancer. Thus, immune checkpoint blockages are recruited and often used in combination therapy. The cytotoxic T lymphocyte associated protein 4 (CTLA-4) monoclonal antibody and the programmed death 1 (PD-1)/programed death ligand-1 (PD-L1) pathways are the spotlight of various cancers [134,135,136]. Investigation of iCCA specimens has shown that PD-L1 is highly expressed in tumor tissues and is inversely related to CD8+ T cell infiltration. The loss of HLA Class 1 antigen expression leads to CCA immune escape. On the other hand, the total mutation burden (TMB) and microsatellite instability (MSI) were reflected in the immune response in cancers. [137]. Therefore, patients with increased cases of high TMB or MSI-H may have a better immune response. Pembrolizumab, the PD-1 monoclonal antibody, was thawed for multiple treatment combinations (NCT02628067, NCT02268825). Merging all the information, we classified the clinical trial phase, targets and status of each potential compound (Table 5).
9. Future Prospects
CCA research still requires greater sample sizes and detailed clinical data as evidence and support. The omics-based dataset and techniques are suitable for CCA bottlenecks in basic research and clinical applications. Using the available information from RNA-sequencing, we can clearly classify subtypes and specific characteristics based on genetic/epigenetic background. Moreover, omics-based datasets also support a large number of expressions of each coding/non-coding gene/protein/metabolites for our analysis and quantification. Multi-omics platforms can merge with events in clinical part, including treatment options, drug response or clinicopathological factors. From these data, reliable prognostic and diagnostic factors can be obtained, and the results can also be judged by the blood, urine, bile as well as the primary tumor of CCA patients. On the other side, the combination of omics-based dataset and bioinformatics tools can reveal available/underlying signatures and mechanisms. Potential inhibitors/small compounds or drug repurposing can also be used to reverse their observations in precision medicine. Therefore, the omics-based approaches have the potential as useful tools to explore a series of multiplex events in CCA. More important, multi-omics approaches can integrate the latest artificial intelligence and machine-learning to improve the accuracy and remain competitive in future research (Figure 5).
Author Contributions
Y.-C.C., M.-H.C and M.H. participated in the conceptualization. C.-N.Y. and M.H. contributed to critical editing of manuscript. Funding is provided by C.-N.Y. and M.H.; Data acquisition was done by Y.-C.C. and M.-H.C. Supervision, M.H. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Acknowledgments
This study was supported by Academia Sinica and Ministry of Science and Technology: AS-SUMMIT-108, AS-SUMMIT-109, ASKPQ-109-BioMed, MOST-108-3114-Y-001-002 and MOST 107-2320-B-001 -016 -MY3 to Michael Hsiao. This study was also supported by the Chang Gung Memorial Hospital: CORPG3J0251~2, CMRPG3I0241~2, CRRPG3K0011, CRRPG3K0031 and CMRPG3K0711 to Chun-Nan Yeh.
Conflicts of Interest
The authors declare that they have no competing interests.
References
- Banales, J.M.; Cardinale, V.; Carpino, G.; Marzioni, M.; Andersen, J.B.; Invernizzi, P.; Lind, G.E.; Folseraas, T.; Forbes, S.J.; Fouassier, L.; et al. Expert consensus document: Cholangiocarcinoma: Current knowledge and future perspectives consensus statement from the European Network for the Study of Cholangiocarcinoma (ENS-CCA). Nat. Rev. Gastroenterol. Hepatol. 2016, 13, 261–280. [Google Scholar] [CrossRef]
- Rizvi, S.; Khan, S.A.; Hallemeier, C.L.; Kelley, R.K.; Gores, G.J. Cholangiocarcinoma—Evolving concepts and therapeutic strategies. Nat. Rev. Clin. Oncol. 2018, 15, 95–111. [Google Scholar] [CrossRef] [Green Version]
- Nakeeb, A.; Pitt, H.A.; Sohn, T.A.; Coleman, J.; Abrams, R.A.; Piantadosi, S.; Hruban, R.H.; Lillemoe, K.D.; Yeo, C.J.; Cameron, J.L. Cholangiocarcinoma. A spectrum of intrahepatic, perihilar, and distal tumors. Ann. Surg. 1996, 224, 463–473. [Google Scholar] [CrossRef] [PubMed]
- Akita, M.; Sofue, K.; Fujikura, K.; Otani, K.; Itoh, T.; Ajiki, T.; Fukumoto, T.; Zen, Y. Histological and molecular characterization of intrahepatic bile duct cancers suggests an expanded definition of perihilar cholangiocarcinoma. HPB 2019, 21, 226–234. [Google Scholar] [CrossRef] [PubMed]
- Banales, J.M.; Marin, J.J.G.; Lamarca, A.; Rodrigues, P.M.; Khan, S.A.; Roberts, L.R.; Cardinale, V.; Carpino, G.; Andersen, J.B.; Braconi, C.; et al. Cholangiocarcinoma 2020: The next horizon in mechanisms and management. Nat. Rev. Gastroenterol. Hepatol. 2020. [Google Scholar] [CrossRef] [PubMed]
- Bertuccio, P.; Malvezzi, M.; Carioli, G.; Hashim, D.; Boffetta, P.; El-Serag, H.B.; La Vecchia, C.; Negri, E. Global trends in mortality from intrahepatic and extrahepatic cholangiocarcinoma. J. Hepatol. 2019, 71, 104–114. [Google Scholar] [CrossRef] [PubMed]
- Tian, W.; Hu, W.; Shi, X.; Liu, P.; Ma, X.; Zhao, W.; Qu, L.; Zhang, S.; Shi, W.; Liu, A.; et al. Comprehensive genomic profile of cholangiocarcinomas in China. Oncol. Lett. 2020, 19, 3101–3110. [Google Scholar] [CrossRef] [Green Version]
- Utada, M.; Ohno, Y.; Tamaki, T.; Sobue, T.; Endo, G. Long-term trends in incidence and mortality of intrahepatic and extrahepatic bile duct cancer in Japan. J. Epidemiol. 2014, 24, 193–199. [Google Scholar] [CrossRef] [Green Version]
- Kim, B.W.; Oh, C.M.; Choi, H.Y.; Park, J.W.; Cho, H.; Ki, M. Incidence and Overall Survival of Biliary Tract Cancers in South Korea from 2006 to 2015: Using the National Health Information Database. Gut Liver 2019, 13, 104–113. [Google Scholar] [CrossRef]
- Chaiteerakij, R.; Pan-Ngum, W.; Poovorawan, K.; Soonthornworasiri, N.; Treeprasertsuk, S.; Phaosawasdi, K. Characteristics and outcomes of cholangiocarcinoma by region in Thailand: A nationwide study. World J. Gastroenterol. 2017, 23, 7160–7167. [Google Scholar] [CrossRef]
- Chen, M.F.; Jan, Y.Y.; Jeng, L.B.; Hwang, T.L.; Wang, C.S.; Chen, S.C.; Chao, T.C.; Chen, H.M.; Lee, W.C.; Yeh, T.S.; et al. Intrahepatic cholangiocarcinoma in Taiwan. J. Hepato-Biliary-Pancreat. Surg. 1999, 6, 136–141. [Google Scholar] [CrossRef] [PubMed]
- Pinter, M.; Hucke, F.; Zielonke, N.; Waldhör, T.; Trauner, M.; Peck-Radosavljevic, M.; Sieghart, W. Incidence and mortality trends for biliary tract cancers in Austria. Liver Int. Off. J. Int. Assoc. Study Liver 2014, 34, 1102–1108. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.J.; Wu, A.T.; Chiou, H.Y.; Chuang, M.T.; Meng, T.C.; Chien, L.N.; Yen, Y. Interactive role of diabetes mellitus and female sex in the risk of cholangiocarcinoma: A population-based nested case-control study. Oncotarget 2017, 8, 6642–6651. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alabraba, E.; Joshi, H.; Bird, N.; Griffin, R.; Sturgess, R.; Stern, N.; Sieberhagen, C.; Cross, T.; Camenzuli, A.; Davis, R.; et al. Increased multimodality treatment options has improved survival for Hepatocellular carcinoma but poor survival for biliary tract cancers remains unchanged. Eur. J. Surg. Oncol. J. Eur. Soc. Surg. Oncol. Br. Assoc. Surg. Oncol. 2019, 45, 1660–1667. [Google Scholar] [CrossRef] [PubMed]
- Cambridge, W.A.; Fairfield, C.; Powell, J.J.; Harrison, E.M.; Søreide, K.; Wigmore, S.J.; Guest, R.V. Meta-analysis and Meta-regression of Survival After Liver Transplantation for Unresectable Perihilar Cholangiocarcinoma. Ann. Surg. 2020. [Google Scholar] [CrossRef] [PubMed]
- Kamsa-Ard, S.; Luvira, V.; Suwanrungruang, K.; Kamsa-Ard, S.; Luvira, V.; Santong, C.; Srisuk, T.; Pugkhem, A.; Bhudhisawasdi, V.; Pairojkul, C. Cholangiocarcinoma Trends, Incidence, and Relative Survival in Khon Kaen, Thailand from 1989 Through 2013: A Population-Based Cancer Registry Study. J. Epidemiol. 2019, 29, 197–204. [Google Scholar] [CrossRef] [Green Version]
- Komaya, K.; Ebata, T.; Yokoyama, Y.; Igami, T.; Sugawara, G.; Mizuno, T.; Yamaguchi, J.; Nagino, M. Recurrence after curative-intent resection of perihilar cholangiocarcinoma: Analysis of a large cohort with a close postoperative follow-up approach. Surgery 2018, 163, 732–738. [Google Scholar] [CrossRef]
- Strijker, M.; Belkouz, A.; van der Geest, L.G.; van Gulik, T.M.; van Hooft, J.E.; de Meijer, V.E.; Haj Mohammad, N.; de Reuver, P.R.; Verheij, J.; de Vos-Geelen, J.; et al. Treatment and survival of resected and unresected distal cholangiocarcinoma: A nationwide study. Acta Oncol. 2019, 58, 1048–1055. [Google Scholar] [CrossRef] [PubMed]
- Sripa, B.; Pairojkul, C. Cholangiocarcinoma: Lessons from Thailand. Curr. Opin. Gastroenterol. 2008, 24, 349–356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rigopoulou, E.I.; Zachou, K.; Gatselis, N.K.; Papadamou, G.; Koukoulis, G.K.; Dalekos, G.N. Primary biliary cirrhosis in HBV and HCV patients: Clinical characteristics and outcome. World J. Hepatol. 2013, 5, 577–583. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Zong, C.; Zhang, L.; Garner, E.; Sugie, S.; Huang, C.; Wu, W.; Chang, J.; Sakurai, T.; Kato, M.; et al. Exposure of Mice to 1,2-Dichloropropane Induces CYP450-Dependent Proliferation and Apoptosis of Cholangiocytes. Toxicol. Sci. Off. J. Soc. Toxicol. 2018, 162, 559–569. [Google Scholar] [CrossRef] [PubMed]
- Valle, J.; Wasan, H.; Palmer, D.H.; Cunningham, D.; Anthoney, A.; Maraveyas, A.; Madhusudan, S.; Iveson, T.; Hughes, S.; Pereira, S.P.; et al. Cisplatin plus gemcitabine versus gemcitabine for biliary tract cancer. N. Engl. J. Med. 2010, 362, 1273–1281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Varamo, C.; Peraldo-Neia, C.; Ostano, P.; Basiricò, M.; Raggi, C.; Bernabei, P.; Venesio, T.; Berrino, E.; Aglietta, M.; Leone, F.; et al. Establishment and Characterization of a New Intrahepatic Cholangiocarcinoma Cell Line Resistant to Gemcitabine. Cancers 2019, 11, 519. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, A.J.; Chun, Y.S. Intrahepatic cholangiocarcinoma: The AJCC/UICC 8th edition updates. Chin. Clin. Oncol. 2018, 7, 52. [Google Scholar] [CrossRef] [PubMed]
- Berger, A.C.; Korkut, A.; Kanchi, R.S.; Hegde, A.M.; Lenoir, W.; Liu, W.; Liu, Y.; Fan, H.; Shen, H.; Ravikumar, V.; et al. A Comprehensive Pan-Cancer Molecular Study of Gynecologic and Breast Cancers. Cancer Cell 2018, 33, 690–705. [Google Scholar] [CrossRef] [Green Version]
- Koplev, S.; Lin, K.; Dohlman, A.B.; Ma’ayan, A. Integration of pan-cancer transcriptomics with RPPA proteomics reveals mechanisms of epithelial-mesenchymal transition. PLoS Comput. Biol. 2018, 14, e1005911. [Google Scholar] [CrossRef] [Green Version]
- Nakamura, H.; Arai, Y.; Totoki, Y.; Shirota, T.; Elzawahry, A.; Kato, M.; Hama, N.; Hosoda, F.; Urushidate, T.; Ohashi, S.; et al. Genomic spectra of biliary tract cancer. Nat. Genet. 2015, 47, 1003–1010. [Google Scholar] [CrossRef]
- Arai, Y.; Totoki, Y.; Hosoda, F.; Shirota, T.; Hama, N.; Nakamura, H.; Ojima, H.; Furuta, K.; Shimada, K.; Okusaka, T.; et al. Fibroblast growth factor receptor 2 tyrosine kinase fusions define a unique molecular subtype of cholangiocarcinoma. Hepatology 2014, 59, 1427–1434. [Google Scholar] [CrossRef]
- Gao, J.; Aksoy, B.A.; Dogrusoz, U.; Dresdner, G.; Gross, B.; Sumer, S.O.; Sun, Y.; Jacobsen, A.; Sinha, R.; Larsson, E.; et al. Integrative analysis of complex cancer genomics and clinical profiles using the cBioPortal. Sci. Signal. 2013, 6, pl1. [Google Scholar] [CrossRef] [Green Version]
- Cerami, E.; Gao, J.; Dogrusoz, U.; Gross, B.E.; Sumer, S.O.; Aksoy, B.A.; Jacobsen, A.; Byrne, C.J.; Heuer, M.L.; Larsson, E.; et al. The cBio cancer genomics portal: An open platform for exploring multidimensional cancer genomics data. Cancer Discov. 2012, 2, 401–404. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.F.; Jiang, H.; Zhang, C.S.; Yu, S.P.; Wang, Z.Q.; Su, H.L. Targeted drug regulation on methylation of p53-BAX mitochondrial apoptosis pathway affects the growth of cholangiocarcinoma cells. J. Int. Med Res. 2012, 40, 67–75. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.F.; Kong, F.M.; Xu, Z.; Yu, S.P.; Sun, F.B.; Zhang, C.S.; Huang, Q.X.; Zhou, X.T.; Song, Z.W. Promoter hypermethylation of death-associated protein kinase gene in cholangiocarcinoma. Hepatobiliary Pancreat. Dis. Int. 2007, 6, 407–411. [Google Scholar] [PubMed]
- Shin, S.H.; Lee, K.; Kim, B.H.; Cho, N.Y.; Jang, J.Y.; Kim, Y.T.; Kim, D.; Jang, J.J.; Kang, G.H. Bile-based detection of extrahepatic cholangiocarcinoma with quantitative DNA methylation markers and its high sensitivity. J. Mol. Diagn. JMD 2012, 14, 256–263. [Google Scholar] [CrossRef] [PubMed]
- Taniai, M.; Higuchi, H.; Burgart, L.J.; Gores, G.J. p16INK4a promoter mutations are frequent in primary sclerosing cholangitis (PSC) and PSC-associated cholangiocarcinoma. Gastroenterology 2002, 123, 1090–1098. [Google Scholar] [CrossRef] [PubMed]
- Tannapfel, A.; Busse, C.; Geissler, F.; Witzigmann, H.; Hauss, J.; Wittekind, C. INK4a-ARF alterations in liver cell adenoma. Gut 2002, 51, 253–258. [Google Scholar] [CrossRef] [Green Version]
- Vedeld, H.M.; Andresen, K.; Eilertsen, I.A.; Nesbakken, A.; Seruca, R.; Gladhaug, I.P.; Thiis-Evensen, E.; Rognum, T.O.; Boberg, K.M.; Lind, G.E. The novel colorectal cancer biomarkers CDO1, ZSCAN18 and ZNF331 are frequently methylated across gastrointestinal cancers. Int. J. Cancer 2015, 136, 844–853. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Zhang, K.; Wang, J.; Wu, X.; Liu, X.; Li, B.; Zhu, Y.; Yu, Y.; Cheng, Q.; Hu, Z.; et al. Underexpression of LKB1 tumor suppressor is associated with enhanced Wnt signaling and malignant characteristics of human intrahepatic cholangiocarcinoma. Oncotarget 2015, 6, 18905–18920. [Google Scholar] [CrossRef] [Green Version]
- Wehbe, H.; Henson, R.; Meng, F.; Mize-Berge, J.; Patel, T. Interleukin-6 contributes to growth in cholangiocarcinoma cells by aberrant promoter methylation and gene expression. Cancer Res. 2006, 66, 10517–10524. [Google Scholar] [CrossRef] [Green Version]
- Xiang, J.; Luo, F.; Chen, Y.; Zhu, F.; Wang, J. si-DNMT1 restore tumor suppressor genes expression through the reversal of DNA hypermethylation in cholangiocarcinoma. Clin. Res. Hepatol. Gastroenterol. 2014, 38, 181–189. [Google Scholar] [CrossRef]
- Xiaofang, L.; Kun, T.; Shaoping, Y.; Zaiqiu, W.; Hailong, S. Correlation between promoter methylation of p14(ARF), TMS1/ASC, and DAPK, and p53 mutation with prognosis in cholangiocarcinoma. World J. Surg. Oncol. 2012, 10, 5. [Google Scholar] [CrossRef] [Green Version]
- Tian, X.; Zhang, S.; Liu, H.M.; Zhang, Y.B.; Blair, C.A.; Mercola, D.; Sassone-Corsi, P.; Zi, X. Histone lysine-specific methyltransferases and demethylases in carcinogenesis: New targets for cancer therapy and prevention. Curr. Cancer Drug Targets 2013, 13, 558–579. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Biegel, J.A.; Busse, T.M.; Weissman, B.E. SWI/SNF chromatin remodeling complexes and cancer. Am. J. Med. Genet. Part C Semin. Med. Genet. 2014, 166c, 350–366. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, R.C.; Wang, T.L.; Shih Ie, M. The emerging roles of ARID1A in tumor suppression. Cancer Biol. Ther. 2014, 15, 655–664. [Google Scholar] [CrossRef] [Green Version]
- Braconi, C.; Huang, N.; Patel, T. MicroRNA-dependent regulation of DNA methyltransferase-1 and tumor suppressor gene expression by interleukin-6 in human malignant cholangiocytes. Hepatology 2010, 51, 881–890. [Google Scholar] [CrossRef] [Green Version]
- Nakaoka, T.; Saito, Y.; Saito, H. Aberrant DNA Methylation as a Biomarker and a Therapeutic Target of Cholangiocarcinoma. Int. J. Mol. Sci. 2017, 18, 1111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meng, F.; Wehbe-Janek, H.; Henson, R.; Smith, H.; Patel, T. Epigenetic regulation of microRNA-370 by interleukin-6 in malignant human cholangiocytes. Oncogene 2008, 27, 378–386. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iwaki, J.; Kikuchi, K.; Mizuguchi, Y.; Kawahigashi, Y.; Yoshida, H.; Uchida, E.; Takizawa, T. MiR-376c down-regulation accelerates EGF-dependent migration by targeting GRB2 in the HuCCT1 human intrahepatic cholangiocarcinoma cell line. PLoS ONE 2013, 8, e69496. [Google Scholar] [CrossRef] [Green Version]
- Yang, R.; Chen, Y.; Tang, C.; Li, H.; Wang, B.; Yan, Q.; Hu, J.; Zou, S. MicroRNA-144 suppresses cholangiocarcinoma cell proliferation and invasion through targeting platelet activating factor acetylhydrolase isoform 1b. BMC Cancer 2014, 14, 917. [Google Scholar] [CrossRef] [Green Version]
- Sarver, A.L.; Sarver, A.E.; Yuan, C.; Subramanian, S. OMCD: OncomiR Cancer Database. BMC Cancer 2018, 18, 1223. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.; Xiao, J.; Chai, Y.; Du, Y.Y.; Liu, Z.; Huang, K.; Zhou, X.; Zhou, W. LncRNA-CCAT1 Promotes Migration, Invasion, and EMT in Intrahepatic Cholangiocarcinoma Through Suppressing miR-152. Dig. Dis. Sci. 2017, 62, 3050–3058. [Google Scholar] [CrossRef]
- Xu, Y.; Yao, Y.; Qin, W.; Zhong, X.; Jiang, X.; Cui, Y. Long non-coding RNA CCAT2 promotes cholangiocarcinoma cells migration and invasion by induction of epithelial-to-mesenchymal transition. Biomed. Pharmacother. 2018, 99, 121–127. [Google Scholar] [CrossRef] [PubMed]
- Zu, C.; Liu, S.; Cao, W.; Liu, Z.; Qiang, H.; Li, Y.; Cheng, C.; Ji, L.; Li, J.; Li, J. MiR-590-3p suppresses epithelial-mesenchymal transition in intrahepatic cholangiocarcinoma by inhibiting SIP1 expression. Oncotarget 2017, 8, 34698–34708. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, Y.; Wang, Z.; Jiang, X.; Cui, Y. Overexpression of long noncoding RNA H19 indicates a poor prognosis for cholangiocarcinoma and promotes cell migration and invasion by affecting epithelial-mesenchymal transition. Biomed. Pharmacother. 2017, 92, 17–23. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Li, J.; Ji, D.; Leng, K.; Xu, Y.; Huang, L.; Jiang, X.; Cui, Y. Overexpressed long noncoding RNA Sox2ot predicts poor prognosis for cholangiocarcinoma and promotes cell proliferation and invasion. Gene 2018, 645, 131–136. [Google Scholar] [CrossRef] [PubMed]
- Gao, L.; Yang, X.; Zhang, H.; Yu, M.; Long, J.; Yang, T. Inhibition of miR-10a-5p suppresses cholangiocarcinoma cell growth through downregulation of Akt pathway. Oncotargets Ther. 2018, 11, 6981–6994. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Lu, L.; Tu, J.; Zhang, J.; Xiong, T.; Fan, W.; Wang, J.; Li, M.; Chen, Y.; Steggerda, J.; et al. Reciprocal Regulation Between Forkhead Box M1/NF-kB and Methionine Adenosyltransferase 1A Drives Liver Cancer. Hepatology 2020. [Google Scholar] [CrossRef]
- Fan, W.; Yang, H.; Liu, T.; Wang, J.; Li, T.W.; Mavila, N.; Tang, Y.; Yang, J.; Peng, H.; Tu, J.; et al. Prohibitin 1 suppresses liver cancer tumorigenesis in mice and human hepatocellular and cholangiocarcinoma cells. Hepatology 2017, 65, 1249–1266. [Google Scholar] [CrossRef] [Green Version]
- Srijiwangsa, P.; Ponnikorn, S.; Na-Bangchang, K. Effect of β-Eudesmol on NQO1 suppression-enhanced sensitivity of cholangiocarcinoma cells to chemotherapeutic agents. BMC Pharmacol. Toxicol. 2018, 19, 32. [Google Scholar] [CrossRef]
- Yeh, C.N.; Chen, M.H.; Chang, Y.C.; Wu, R.C.; Tsao, L.C.; Wang, S.Y.; Cheng, C.T.; Chiang, K.C.; Chen, T.W.; Hsiao, M.; et al. Over-expression of TNNI3K is associated with early-stage carcinogenesis of cholangiocarcinoma. Mol. Carcinog. 2019, 58, 270–278. [Google Scholar] [CrossRef]
- Hatzaras, I.; Schmidt, C.; Muscarella, P.; Melvin, W.S.; Ellison, E.C.; Bloomston, M. Elevated CA 19-9 portends poor prognosis in patients undergoing resection of biliary malignancies. HPB Off. J. Int. Hepato Pancreato Biliary Assoc. 2010, 12, 134–138. [Google Scholar] [CrossRef] [Green Version]
- Aksorn, N.; Roytrakul, S.; Kittisenachai, S.; Leelawat, K.; Chanvorachote, P.; Topanurak, S.; Hamano, S.; Lek-Uthai, U. Novel Potential Biomarkers for Opisthorchis viverrini Infection and Associated Cholangiocarcinoma. In Vivo 2018, 32, 871–878. [Google Scholar] [CrossRef] [PubMed]
- Boonla, C.; Sripa, B.; Thuwajit, P.; Cha-On, U.; Puapairoj, A.; Miwa, M.; Wongkham, S. MUC1 and MUC5AC mucin expression in liver fluke-associated intrahepatic cholangiocarcinoma. World J. Gastroenterol. 2005, 11, 4939–4946. [Google Scholar] [CrossRef] [PubMed]
- Gardini, A.; Corti, B.; Fiorentino, M.; Altimari, A.; Ercolani, G.; Grazi, G.L.; Pinna, A.D.; Grigioni, W.F.; D’Errico Grigioni, A. Expression of connective tissue growth factor is a prognostic marker for patients with intrahepatic cholangiocarcinoma. Dig. Liver Dis. Off. J. Ital. Soc. Gastroenterol. Ital. Assoc. Study Liver 2005, 37, 269–274. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.Y.; Ke, A.W.; Shi, G.M.; Ding, Z.B.; Devbhandari, R.P.; Gu, F.M.; Li, Q.L.; Dai, Z.; Zhou, J.; Fan, J. Overexpression of CD151 as an adverse marker for intrahepatic cholangiocarcinoma patients. Cancer 2010, 116, 5440–5451. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.F.; Zhao, R.; Guo, S.; Wang, X.Q.; Lian, P.L.; Chen, Y.G.; Xu, K.S. Expression and clinical significance of hepatoma-derived growth factor as a prognostic factor in human hilar cholangiocarcinoma. Ann. Surg. Oncol. 2011, 18, 872–879. [Google Scholar] [CrossRef]
- Mao, X.; Chen, D.; Wu, J.; Li, J.; Zhou, H.; Wu, Y.; Duan, X. Differential expression of fascin, E-cadherin and vimentin: Proteins associated with survival of cholangiocarcinoma patients. Am. J. Med. Sci. 2013, 346, 261–268. [Google Scholar] [CrossRef]
- Merino-Azpitarte, M.; Lozano, E.; Perugorria, M.J.; Esparza-Baquer, A.; Erice, O.; Santos-Laso, Á.; O’Rourke, C.J.; Andersen, J.B.; Jiménez-Agüero, R.; Lacasta, A.; et al. SOX17 regulates cholangiocyte differentiation and acts as a tumor suppressor in cholangiocarcinoma. J. Hepatol. 2017, 67, 72–83. [Google Scholar] [CrossRef]
- Morine, Y.; Imura, S.; Ikemoto, T.; Iwahashi, S.; Saito, Y.U.; Shimada, M. CD44 Expression Is a Prognostic Factor in Patients with Intrahepatic Cholangiocarcinoma after Surgical Resection. Anticancer Res. 2017, 37, 5701–5705. [Google Scholar] [CrossRef]
- Nepal, C.; O’Rourke, C.J.; Oliveira, D.; Taranta, A.; Shema, S.; Gautam, P.; Calderaro, J.; Barbour, A.; Raggi, C.; Wennerberg, K.; et al. Genomic perturbations reveal distinct regulatory networks in intrahepatic cholangiocarcinoma. Hepatology 2018, 68, 949–963. [Google Scholar] [CrossRef] [Green Version]
- Shi, R.Y.; Yang, X.R.; Shen, Q.J.; Yang, L.X.; Xu, Y.; Qiu, S.J.; Sun, Y.F.; Zhang, X.; Wang, Z.; Zhu, K.; et al. High expression of Dickkopf-related protein 1 is related to lymphatic metastasis and indicates poor prognosis in intrahepatic cholangiocarcinoma patients after surgery. Cancer 2013, 119, 993–1003. [Google Scholar] [CrossRef]
- Shimada, M.; Sugimoto, K.; Iwahashi, S.; Utsunomiya, T.; Morine, Y.; Imura, S.; Ikemoto, T. CD133 expression is a potential prognostic indicator in intrahepatic cholangiocarcinoma. J. Gastroenterol. 2010, 45, 896–902. [Google Scholar] [CrossRef]
- Yoshikawa, D.; Ojima, H.; Iwasaki, M.; Hiraoka, N.; Kosuge, T.; Kasai, S.; Hirohashi, S.; Shibata, T. Clinicopathological and prognostic significance of EGFR, VEGF, and HER2 expression in cholangiocarcinoma. Br. J. Cancer 2008, 98, 418–425. [Google Scholar] [CrossRef]
- Navaneethan, U.; Lourdusamy, V.; Gk Venkatesh, P.; Willard, B.; Sanaka, M.R.; Parsi, M.A. Bile proteomics for differentiation of malignant from benign biliary strictures: A pilot study. Gastroenterol. Rep. 2015, 3, 136–143. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Morikawa, K.; Mori, Y.; Zong, C.; Zhang, L.; Garner, E.; Huang, C.; Wu, W.; Chang, J.; Nagashima, D.; et al. Proteomic analysis of liver proteins of mice exposed to 1,2-dichloropropane. Arch. Toxicol. 2020. [Google Scholar] [CrossRef]
- Son, K.H.; Ahn, C.B.; Kim, H.J.; Kim, J.S. Quantitative proteomic analysis of bile in extrahepatic cholangiocarcinoma patients. J. Cancer 2020, 11, 4073–4080. [Google Scholar] [CrossRef]
- Le Large, T.Y.S.; Meijer, L.L.; Paleckyte, R.; Boyd, L.N.C.; Kok, B.; Wurdinger, T.; Schelfhorst, T.; Piersma, S.R.; Pham, T.V.; van Grieken, N.C.T.; et al. Combined Expression of Plasma Thrombospondin-2 and CA19-9 for Diagnosis of Pancreatic Cancer and Distal Cholangiocarcinoma: A Proteome Approach. Oncologist 2020, 25, 634–643. [Google Scholar] [CrossRef] [Green Version]
- Cheon, Y.K.; Cho, Y.D.; Moon, J.H.; Jang, J.Y.; Kim, Y.S.; Kim, Y.S.; Lee, M.S.; Lee, J.S.; Shim, C.S. Diagnostic utility of interleukin-6 (IL-6) for primary bile duct cancer and changes in serum IL-6 levels following photodynamic therapy. Am. J. Gastroenterol. 2007, 102, 2164–2170. [Google Scholar] [CrossRef]
- Leelawat, K.; Sakchinabut, S.; Narong, S.; Wannaprasert, J. Detection of serum MMP-7 and MMP-9 in cholangiocarcinoma patients: Evaluation of diagnostic accuracy. BMC Gastroenterol. 2009, 9, 30. [Google Scholar] [CrossRef] [Green Version]
- Onsurathum, S.; Haonon, O.; Pinlaor, P.; Pairojkul, C.; Khuntikeo, N.; Thanan, R.; Roytrakul, S.; Pinlaor, S. Proteomics detection of S100A6 in tumor tissue interstitial fluid and evaluation of its potential as a biomarker of cholangiocarcinoma. Tumour Biol. J. Int. Soc. Oncodev. Biol. Med. 2018, 40. [Google Scholar] [CrossRef] [Green Version]
- Shen, J.; Wang, W.; Wu, J.; Feng, B.; Chen, W.; Wang, M.; Tang, J.; Wang, F.; Cheng, F.; Pu, L.; et al. Comparative proteomic profiling of human bile reveals SSP411 as a novel biomarker of cholangiocarcinoma. PLoS ONE 2012, 7, e47476. [Google Scholar] [CrossRef]
- Wang, M.; Fang, M.; Zhu, J.; Feng, H.; Warner, E.; Yi, C.; Ji, J.; Gu, X.; Gao, C. Serum N-glycans outperform CA19-9 in diagnosis of extrahepatic cholangiocarcinoma. Electrophoresis 2017, 38, 2749–2756. [Google Scholar] [CrossRef]
- Da, Z.; Gao, L.; Su, G.; Yao, J.; Fu, W.; Zhang, J.; Zhang, X.; Pei, Z.; Yue, P.; Bai, B.; et al. Bioinformatics combined with quantitative proteomics analyses and identification of potential biomarkers in cholangiocarcinoma. Cancer Cell Int. 2020, 20, 130. [Google Scholar] [CrossRef]
- Sulpice, L.; Desille, M.; Turlin, B.; Fautrel, A.; Boudjema, K.; Clément, B.; Coulouarn, C. Gene expression profiling of the tumor microenvironment in human intrahepatic cholangiocarcinoma. Genom. Data 2016, 7, 229–232. [Google Scholar] [CrossRef]
- Sulpice, L.; Rayar, M.; Desille, M.; Turlin, B.; Fautrel, A.; Boucher, E.; Llamas-Gutierrez, F.; Meunier, B.; Boudjema, K.; Clément, B.; et al. Molecular profiling of stroma identifies osteopontin as an independent predictor of poor prognosis in intrahepatic cholangiocarcinoma. Hepatology 2013, 58, 1992–2000. [Google Scholar] [CrossRef] [PubMed]
- Peraldo Neia, C.; Cavalloni, G.; Chiorino, G.; Ostano, P.; Aglietta, M.; Leone, F. Gene and microRNA modulation upon trabectedin treatment in a human intrahepatic cholangiocarcinoma paired patient derived xenograft and cell line. Oncotarget 2016, 7, 86766–86780. [Google Scholar] [CrossRef] [Green Version]
- Meijer, L.L.; Puik, J.R.; Le Large, T.Y.S.; Heger, M.; Dijk, F.; Funel, N.; Wurdinger, T.; Garajová, I.; van Grieken, N.C.T.; van de Wiel, M.A.; et al. Unravelling the Diagnostic Dilemma: A MicroRNA Panel of Circulating MiR-16 and MiR-877 as A Diagnostic Classifier for Distal Bile Duct Tumors. Cancers 2019, 11, 1181. [Google Scholar] [CrossRef] [Green Version]
- Zhang, M.; Yang, H.; Wan, L.; Wang, Z.; Wang, H.; Ge, C.; Liu, Y.; Hao, Y.; Zhang, D.; Shi, G.; et al. Single cell transcriptomic architecture and intercellular crosstalk of human intrahepatic cholangiocarcinoma. J. Hepatol. 2020. [Google Scholar] [CrossRef]
- Saengboonmee, C.; Seubwai, W.; Pairojkul, C.; Wongkham, S. High glucose enhances progression of cholangiocarcinoma cells via STAT3 activation. Sci. Rep. 2016, 6, 18995. [Google Scholar] [CrossRef]
- Ma, L.; Hernandez, M.O.; Zhao, Y.; Mehta, M.; Tran, B.; Kelly, M.; Rae, Z.; Hernandez, J.M.; Davis, J.L.; Martin, S.P.; et al. Tumor Cell Biodiversity Drives Microenvironmental Reprogramming in Liver Cancer. Cancer Cell 2019, 36, 418–430. [Google Scholar] [CrossRef] [Green Version]
- Lapitz, A.; Arbelaiz, A.; O’Rourke, C.J.; Lavin, J.L.; Casta, A.; Ibarra, C.; Jimeno, J.P.; Santos-Laso, A.; Izquierdo-Sanchez, L.; Krawczyk, M.; et al. Patients with Cholangiocarcinoma Present Specific RNA Profiles in Serum and Urine Extracellular Vesicles Mirroring the Tumor Expression: Novel Liquid Biopsy Biomarkers for Disease Diagnosis. Cells 2020, 9, 721. [Google Scholar] [CrossRef] [Green Version]
- Bledea, R.; Vasudevaraja, V.; Patel, S.; Stafford, J.; Serrano, J.; Esposito, G.; Tredwin, L.M.; Goodman, N.; Kloetgen, A.; Golfinos, J.G.; et al. Functional and topographic effects on DNA methylation in IDH1/2 mutant cancers. Sci. Rep. 2019, 9, 16830. [Google Scholar] [CrossRef] [PubMed]
- Montal, R.; Sia, D.; Montironi, C.; Leow, W.Q.; Esteban-Fabró, R.; Pinyol, R.; Torres-Martin, M.; Bassaganyas, L.; Moeini, A.; Peix, J.; et al. Molecular classification and therapeutic targets in extrahepatic cholangiocarcinoma. J. Hepatol. 2020. [Google Scholar] [CrossRef]
- Mondesir, J.; Willekens, C.; Touat, M.; de Botton, S. IDH1 and IDH2 mutations as novel therapeutic targets: Current perspectives. J. Blood Med. 2016, 7, 171–180. [Google Scholar] [CrossRef] [Green Version]
- Raineri, S.; Mellor, J. IDH1: Linking Metabolism and Epigenetics. Front. Genet. 2018, 9, 493. [Google Scholar] [CrossRef] [Green Version]
- Rasmussen, K.D.; Helin, K. Role of TET enzymes in DNA methylation, development, and cancer. Genes Dev. 2016, 30, 733–750. [Google Scholar] [CrossRef]
- Liang, Q.; Liu, H.; Zhang, T.; Jiang, Y.; Xing, H.; Zhang, H. Serum metabolomics uncovering specific metabolite signatures of intra- and extrahepatic cholangiocarcinoma. Mol. Biosyst. 2016, 12, 334–340. [Google Scholar] [CrossRef]
- Alsaleh, M.; Leftley, Z.; Barbera, T.A.; Koomson, L.K.; Zabron, A.; Crossey, M.M.E.; Reeves, H.L.; Cramp, M.; Ryder, S.; Greer, S.; et al. Characterisation of the Serum Metabolic Signature of Cholangiocarcinoma in a United Kingdom Cohort. J. Clin. Exp. Hepatol. 2020, 10, 17–29. [Google Scholar] [CrossRef]
- Alsaleh, M.; Sithithaworn, P.; Khuntikeo, N.; Loilome, W.; Yongvanit, P.; Chamadol, N.; Hughes, T.; O’Connor, T.; Andrews, R.H.; Holmes, E.; et al. Characterisation of the Urinary Metabolic Profile of Liver Fluke-Associated Cholangiocarcinoma. J. Clin. Exp. Hepatol. 2019, 9, 657–675. [Google Scholar] [CrossRef] [Green Version]
- Prasopdee, S.; Thitapakorn, V.; Sathavornmanee, T.; Tesana, S. A comprehensive review of omics and host-parasite interplays studies, towards control of Opisthorchis viverrini infection for prevention of cholangiocarcinoma. Acta Trop. 2019, 196, 76–82. [Google Scholar] [CrossRef]
- Winter, H.; Kaisaki, P.J.; Harvey, J.; Giacopuzzi, E.; Ferla, M.P.; Pentony, M.M.; Knight, S.J.L.; Sharma, R.A.; Taylor, J.C.; McCullagh, J.S.O. Identification of Circulating Genomic and Metabolic Biomarkers in Intrahepatic Cholangiocarcinoma. Cancers 2019, 11, 1895. [Google Scholar] [CrossRef] [Green Version]
- Banales, J.M.; Iñarrairaegui, M.; Arbelaiz, A.; Milkiewicz, P.; Muntané, J.; Muñoz-Bellvis, L.; La Casta, A.; Gonzalez, L.M.; Arretxe, E.; Alonso, C.; et al. Serum Metabolites as Diagnostic Biomarkers for Cholangiocarcinoma, Hepatocellular Carcinoma, and Primary Sclerosing Cholangitis. Hepatology 2019, 70, 547–562. [Google Scholar] [CrossRef] [Green Version]
- Kotawong, K.; Chaijaroenkul, W.; Roytrakul, S.; Phaonakrop, N.; Na-Bangchang, K. Screening of Molecular Targets of Action of Atractylodin in Cholangiocarcinoma by Applying Proteomic and Metabolomic Approaches. Metabolites 2019, 9, 260. [Google Scholar] [CrossRef] [Green Version]
- Murakami, Y.; Kubo, S.; Tamori, A.; Itami, S.; Kawamura, E.; Iwaisako, K.; Ikeda, K.; Kawada, N.; Ochiya, T.; Taguchi, Y.H. Comprehensive analysis of transcriptome and metabolome analysis in Intrahepatic Cholangiocarcinoma and Hepatocellular Carcinoma. Sci. Rep. 2015, 5, 16294. [Google Scholar] [CrossRef] [Green Version]
- Urman, J.M.; Herranz, J.M.; Uriarte, I.; Rullán, M.; Oyón, D.; González, B.; Fernandez-Urién, I.; Carrascosa, J.; Bolado, F.; Zabalza, L.; et al. Pilot Multi-Omic Analysis of Human Bile from Benign and Malignant Biliary Strictures: A Machine-Learning Approach. Cancers 2020, 12, 1644. [Google Scholar] [CrossRef]
- Zhou, Z.; Ma, J. miR-378 serves as a prognostic biomarker in cholangiocarcinoma and promotes tumor proliferation, migration, and invasion. Cancer Biomark. Sect. A Dis. Markers 2019, 24, 173–181. [Google Scholar] [CrossRef]
- Liu, C.H.; Huang, Q.; Jin, Z.Y.; Xie, F.; Zhu, C.L.; Liu, Z.; Wang, C. Circulating microRNA-21 as a prognostic, biological marker in cholangiocarcinoma. J. Cancer Res. Ther. 2018, 14, 220–225. [Google Scholar] [CrossRef]
- Silakit, R.; Loilome, W.; Yongvanit, P.; Chusorn, P.; Techasen, A.; Boonmars, T.; Khuntikeo, N.; Chamadol, N.; Pairojkul, C.; Namwat, N. Circulating miR-192 in liver fluke-associated cholangiocarcinoma patients: A prospective prognostic indicator. J. Hepato-Biliary-Pancreat. Sci. 2014, 21, 864–872. [Google Scholar] [CrossRef]
- Wang, L.J.; Zhang, K.L.; Zhang, N.; Ma, X.W.; Yan, S.W.; Cao, D.H.; Shi, S.J. Serum miR-26a as a diagnostic and prognostic biomarker in cholangiocarcinoma. Oncotarget 2015, 6, 18631–18640. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Wan, M.; Zhou, Q.; Wang, H.; Wang, Z.; Zhong, X.; Zhang, L.; Tai, S.; Cui, Y. The Prognostic Role of SOCS3 and A20 in Human Cholangiocarcinoma. PLoS ONE 2015, 10, e0141165. [Google Scholar] [CrossRef]
- Thongsom, S.; Chaocharoen, W.; Silsirivanit, A.; Wongkham, S.; Sripa, B.; Choe, H.; Suginta, W.; Talabnin, C. YKL-40/chitinase-3-like protein 1 is associated with poor prognosis and promotes cell growth and migration of cholangiocarcinoma. Tumour Biol. J. Int. Soc. Oncodev. Biol. Med. 2016, 37, 9451–9463. [Google Scholar] [CrossRef]
- Tang, Z.; Li, C.; Kang, B.; Gao, G.; Li, C.; Zhang, Z. GEPIA: A web server for cancer and normal gene expression profiling and interactive analyses. Nucleic Acids Res. 2017, 45, 98–102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, C.; Ma, S. A selective review of robust variable selection with applications in bioinformatics. Brief. Bioinform. 2015, 16, 873–883. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, C.; Zhou, F.; Ren, J.; Li, X.; Jiang, Y.; Ma, S. A Selective Review of Multi-Level Omics Data Integration Using Variable Selection. High-Throughput 2019, 8, 4. [Google Scholar] [CrossRef] [Green Version]
- Abou-Alfa, G.K.; Sahai, V.; Hollebecque, A.; Vaccaro, G.; Melisi, D.; Al-Rajabi, R.; Paulson, A.S.; Borad, M.J.; Gallinson, D.; Murphy, A.G.; et al. Pemigatinib for previously treated, locally advanced or metastatic cholangiocarcinoma: A multicentre, open-label, phase 2 study. Lancet Oncol. 2020, 21, 671–684. [Google Scholar] [CrossRef]
- Javle, M.; Lowery, M.; Shroff, R.T.; Weiss, K.H.; Springfeld, C.; Borad, M.J.; Ramanathan, R.K.; Goyal, L.; Sadeghi, S.; Macarulla, T.; et al. Phase II Study of BGJ398 in Patients With FGFR-Altered Advanced Cholangiocarcinoma. J. Clin. Oncol. Off. J. Am. Soc. Clin. Oncol. 2018, 36, 276–282. [Google Scholar] [CrossRef] [PubMed]
- Hall, T.G.; Yu, Y.; Eathiraj, S.; Wang, Y.; Savage, R.E.; Lapierre, J.M.; Schwartz, B.; Abbadessa, G. Preclinical Activity of ARQ 087, a Novel Inhibitor Targeting FGFR Dysregulation. PLoS ONE 2016, 11, e0162594. [Google Scholar] [CrossRef] [Green Version]
- Goyal, L.; Shi, L.; Liu, L.Y.; Fece de la Cruz, F.; Lennerz, J.K.; Raghavan, S.; Leschiner, I.; Elagina, L.; Siravegna, G.; Ng, R.W.S.; et al. TAS-120 Overcomes Resistance to ATP-Competitive FGFR Inhibitors in Patients with FGFR2 Fusion-Positive Intrahepatic Cholangiocarcinoma. Cancer Discov. 2019, 9, 1064–1079. [Google Scholar] [CrossRef] [Green Version]
- Chae, Y.K.; Hong, F.; Vaklavas, C.; Cheng, H.H.; Hammerman, P.; Mitchell, E.P.; Zwiebel, J.A.; Ivy, S.P.; Gray, R.J.; Li, S.; et al. Phase II Study of AZD4547 in Patients With Tumors Harboring Aberrations in the FGFR Pathway: Results From the NCI-MATCH Trial (EAY131) Subprotocol W. J. Clin. Oncol. Off. J. Am. Soc. Clin. Oncol. 2020, 38, 2407–2417. [Google Scholar] [CrossRef]
- Voss, M.H.; Hierro, C.; Heist, R.S.; Cleary, J.M.; Meric-Bernstam, F.; Tabernero, J.; Janku, F.; Gandhi, L.; Iafrate, A.J.; Borger, D.R.; et al. A Phase I, Open-Label, Multicenter, Dose-escalation Study of the Oral Selective FGFR Inhibitor Debio 1347 in Patients with Advanced Solid Tumors Harboring FGFR Gene Alterations. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2019, 25, 2699–2707. [Google Scholar] [CrossRef] [Green Version]
- Borad, M.J.; Champion, M.D.; Egan, J.B.; Liang, W.S.; Fonseca, R.; Bryce, A.H.; McCullough, A.E.; Barrett, M.T.; Hunt, K.; Patel, M.D.; et al. Integrated genomic characterization reveals novel, therapeutically relevant drug targets in FGFR and EGFR pathways in sporadic intrahepatic cholangiocarcinoma. PLoS Genet. 2014, 10, e1004135. [Google Scholar] [CrossRef]
- Chong, D.Q.; Zhu, A.X. The landscape of targeted therapies for cholangiocarcinoma: Current status and emerging targets. Oncotarget 2016, 7, 46750–46767. [Google Scholar] [CrossRef] [Green Version]
- Hedvat, M.; Huszar, D.; Herrmann, A.; Gozgit, J.M.; Schroeder, A.; Sheehy, A.; Buettner, R.; Proia, D.; Kowolik, C.M.; Xin, H.; et al. The JAK2 inhibitor AZD1480 potently blocks Stat3 signaling and oncogenesis in solid tumors. Cancer Cell 2009, 16, 487–497. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kotschy, A.; Szlavik, Z.; Murray, J.; Davidson, J.; Maragno, A.L.; Le Toumelin-Braizat, G.; Chanrion, M.; Kelly, G.L.; Gong, J.N.; Moujalled, D.M.; et al. The MCL1 inhibitor S63845 is tolerable and effective in diverse cancer models. Nature 2016, 538, 477–482. [Google Scholar] [CrossRef] [PubMed]
- Bekaii-Saab, T.; Phelps, M.A.; Li, X.; Saji, M.; Goff, L.; Kauh, J.S.; O’Neil, B.H.; Balsom, S.; Balint, C.; Liersemann, R.; et al. Multi-institutional phase II study of selumetinib in patients with metastatic biliary cancers. J. Clin. Oncol. Off. J. Am. Soc. Clin. Oncol. 2011, 29, 2357–2363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Padthaisong, S.; Dokduang, H.; Yothaisong, S.; Techasen, A.; Namwat, N.; Yongvanit, P.; Khuntikeo, N.; Titapun, A.; Sangkhamanon, S.; Loilome, W. Inhibitory effect of NVP-BKM120 on cholangiocarcinoma cell growth. Oncol. Lett. 2018, 16, 1627–1633. [Google Scholar] [CrossRef]
- Fujisaka, Y.; Kurata, T.; Tanaka, K.; Kudo, T.; Okamoto, K.; Tsurutani, J.; Kaneda, H.; Okamoto, I.; Namiki, M.; Kitamura, C.; et al. Phase I study of amatuximab, a novel monoclonal antibody to mesothelin, in Japanese patients with advanced solid tumors. Investig. New Drugs 2015, 33, 380–388. [Google Scholar] [CrossRef] [Green Version]
- Kwak, T.W.; Kim, D.H.; Jeong, Y.I.; Kang, D.H. Antitumor activity of vorinostat-incorporated nanoparticles against human cholangiocarcinoma cells. J. Nanobiotechnol. 2015, 13, 60. [Google Scholar] [CrossRef] [Green Version]
- Wang, B.; Li, H.; Yang, R.; Zhou, S.; Zou, S. Decitabine inhibits the cell growth of cholangiocarcinoma in cultured cell lines and mouse xenografts. Oncol. Lett. 2014, 8, 1919–1924. [Google Scholar] [CrossRef]
- Zhang, M.; Pan, Y.; Tang, D.; Dorfman, R.G.; Xu, L.; Zhou, Q.; Zhou, L.; Wang, Y.; Li, Y.; Yin, Y.; et al. Low levels of pyruvate induced by a positive feedback loop protects cholangiocarcinoma cells from apoptosis. Cell Commun. Signal. CCS 2019, 17, 23. [Google Scholar] [CrossRef] [Green Version]
- Pant, S.; Saleh, M.; Bendell, J.; Infante, J.R.; Jones, S.; Kurkjian, C.D.; Moore, K.M.; Kazakin, J.; Abbadessa, G.; Wang, Y.; et al. A phase I dose escalation study of oral c-MET inhibitor tivantinib (ARQ 197) in combination with gemcitabine in patients with solid tumors. Ann. Oncol. Off. J. Eur. Soc. Med. Oncol. 2014, 25, 1416–1421. [Google Scholar] [CrossRef]
- Demols, A.; Borbath, I.; Van den Eynde, M.; Houbiers, G.; Peeters, M.; Marechal, R.; Delaunoit, T.; Goemine, J.C.; Laurent, S.; Holbrechts, S.; et al. Regorafenib after failure of gemcitabine and platinum-based chemotherapy for locally advanced/metastatic biliary tumors: REACHIN, a randomized, double-blind, phase II trial. Ann. Oncol. Off. J. Eur. Soc. Med. Oncol. 2020. [Google Scholar] [CrossRef]
- Sun, W.; Patel, A.; Normolle, D.; Patel, K.; Ohr, J.; Lee, J.J.; Bahary, N.; Chu, E.; Streeter, N.; Drummond, S. A phase 2 trial of regorafenib as a single agent in patients with chemotherapy-refractory, advanced, and metastatic biliary tract adenocarcinoma. Cancer 2019, 125, 902–909. [Google Scholar] [CrossRef] [PubMed]
- Yeh, C.N.; Chang, Y.C.; Su, Y.; Shin-Shian Hsu, D.; Cheng, C.T.; Wu, R.C.; Chung, Y.H.; Chiang, K.C.; Yeh, T.S.; Lu, M.L.; et al. Identification of MALT1 as both a prognostic factor and a potential therapeutic target of regorafenib in cholangiocarcinoma patients. Oncotarget 2017, 8, 113444–113459. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fontugne, J.; Augustin, J.; Pujals, A.; Compagnon, P.; Rousseau, B.; Luciani, A.; Tournigand, C.; Cherqui, D.; Azoulay, D.; Pawlotsky, J.M.; et al. PD-L1 expression in perihilar and intrahepatic cholangiocarcinoma. Oncotarget 2017, 8, 24644–24651. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, G.; Sun, L.; Li, Y.; Xie, F.; Zhou, X.; Yang, H.; Du, S.; Xu, H.; Mao, Y. The Clinicopathological and Prognostic Value of PD-L1 Expression in Cholangiocarcinoma: A Meta-Analysis. Front. Oncol. 2019, 9, 897. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Wang, X.Y.; Zhang, Y.; Xu, D.; Dong, J.; Zhang, Z.; Yi, C.H.; Jia, H.L.; Yang, X. Programmed death ligand 1 expression in human intrahepatic cholangiocarcinoma and its association with prognosis and CD8(+) T-cell immune responses. Cancer Manag. Res. 2018, 10, 4113–4123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tan, K.T.; Yeh, C.N.; Chang, Y.C.; Cheng, J.H.; Fang, W.L.; Yeh, Y.C.; Wang, Y.C.; Hsu, D.S.; Wu, C.E.; Lai, J.I.; et al. PRKDC: New biomarker and drug target for checkpoint blockade immunotherapy. J. Immunother. Cancer 2020, 8. [Google Scholar] [CrossRef] [Green Version]
Figure 1.
Meta-analysis of cholangiocarcinoma (CCA) demographics and clinical events. Study recruited six cohorts, including MSK, National Cancer Center of Singapore, National University of Singapore, TCGA, JHU and Shanghai) with a total of 409 patients (412 samples). We summarized the clinical information through the cBioPortal website. (A) Average overall survival probability in the study; (B) average disease-free survival rate in the study; (C) ratio of mutation counts and the range of changes in the number of genome alters, Y-axis = samples; (D) proportion graph indicates the ratio of mutation counts and the range of changes in the number of genome alters; (E) range of microsatellite instability (MSI) scores in the study, Y-axis = samples; (F) circle graphs show percentage of various clinical parameters in the study.
Figure 1.
Meta-analysis of cholangiocarcinoma (CCA) demographics and clinical events. Study recruited six cohorts, including MSK, National Cancer Center of Singapore, National University of Singapore, TCGA, JHU and Shanghai) with a total of 409 patients (412 samples). We summarized the clinical information through the cBioPortal website. (A) Average overall survival probability in the study; (B) average disease-free survival rate in the study; (C) ratio of mutation counts and the range of changes in the number of genome alters, Y-axis = samples; (D) proportion graph indicates the ratio of mutation counts and the range of changes in the number of genome alters; (E) range of microsatellite instability (MSI) scores in the study, Y-axis = samples; (F) circle graphs show percentage of various clinical parameters in the study.
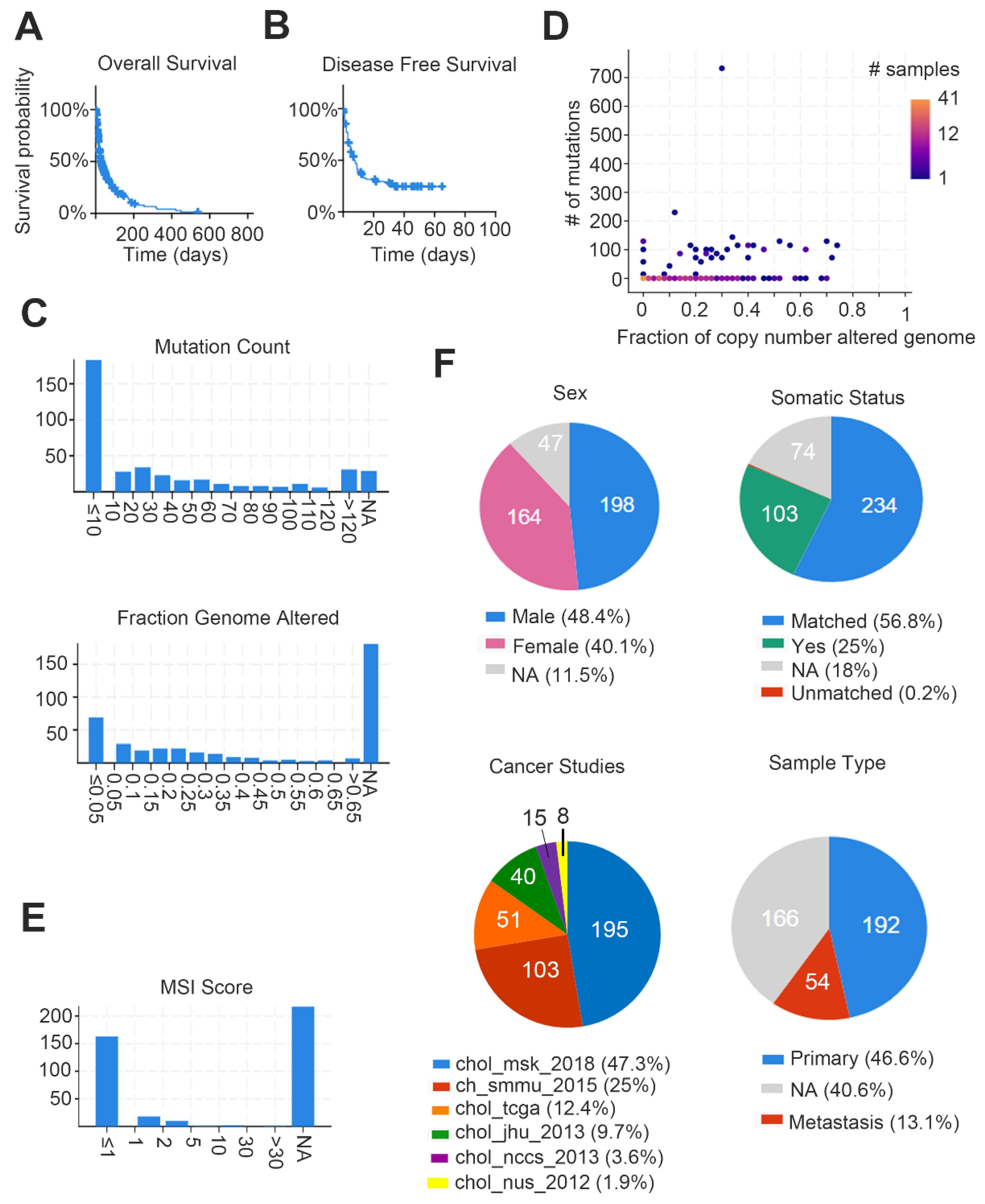
Figure 2.
Meta-analysis of genetic alterations in various CCA cohorts. Study recruited six cohorts, including MSK, National Cancer Center of Singapore, National University of Singapore, TCGA, JHU and Shanghai) with a total of 409 patients (412 samples). We summarized the plot by RNA-sequencing of CCA patients on the cBioPortal website. Graph collects the types and patterns of genetic alteration of multiple targets.
Figure 2.
Meta-analysis of genetic alterations in various CCA cohorts. Study recruited six cohorts, including MSK, National Cancer Center of Singapore, National University of Singapore, TCGA, JHU and Shanghai) with a total of 409 patients (412 samples). We summarized the plot by RNA-sequencing of CCA patients on the cBioPortal website. Graph collects the types and patterns of genetic alteration of multiple targets.

Figure 3.
Most significant differences of miRNAs in CCA. List selected from OncoMir Cancer Database (OMCD) website. After statistical analysis (t-test), the top four upregulated miRNAs and the top 15 downregulated miRNAs in CCA were selected (ratio > 5 or < 0.2, p-value < 0.001).
Figure 3.
Most significant differences of miRNAs in CCA. List selected from OncoMir Cancer Database (OMCD) website. After statistical analysis (t-test), the top four upregulated miRNAs and the top 15 downregulated miRNAs in CCA were selected (ratio > 5 or < 0.2, p-value < 0.001).
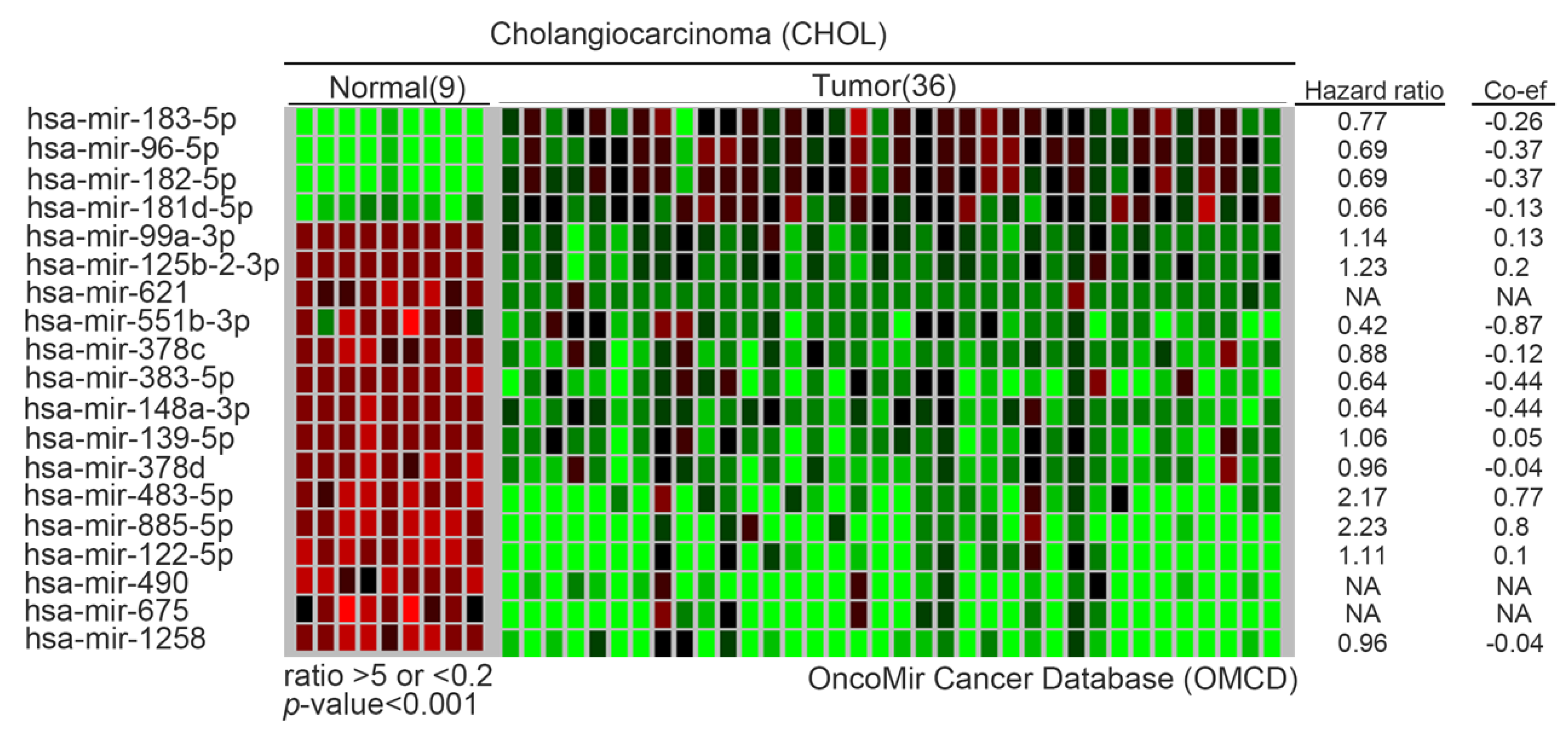
Figure 4.
Network of most differential genes in CCA. All candidates were determined that overexpress in CCA tumors compared with normal parts from gene expression profiling interactive analysis (GEPIA) website. (A) String and (B) ingenuity pathway analysis websites predict potential interactions, transcription factors and networks (p < 0.01).
Figure 4.
Network of most differential genes in CCA. All candidates were determined that overexpress in CCA tumors compared with normal parts from gene expression profiling interactive analysis (GEPIA) website. (A) String and (B) ingenuity pathway analysis websites predict potential interactions, transcription factors and networks (p < 0.01).
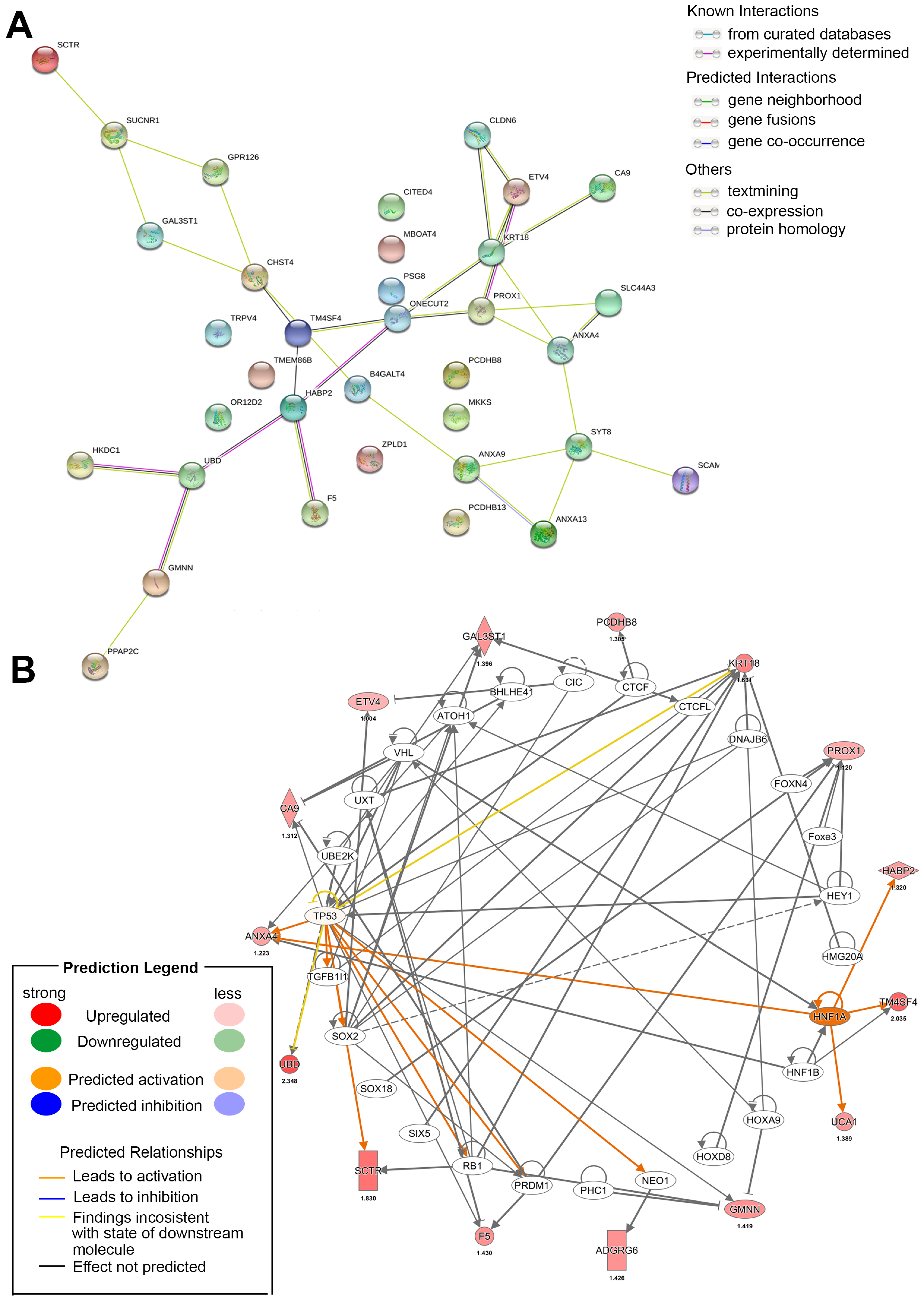
Figure 5.
Schematic model of CCA approaches and applications. In this overview, we show several CCA subtypes according to the location of CCA, including intrahepatic (iCCA), perihepatic (pCCA) and extrahepatic (eCCA). We collected previous studies to indicate various mechanisms and experiments in patients’ blood, urine and primary tissue specimens. This schematic model illustrates that omics-based approaches can be used and integrated for further CCA basic and clinical research. In this topic, we input genomics, epigenomics, transcriptomics, proteomics and metabolomics observations for readers.
Figure 5.
Schematic model of CCA approaches and applications. In this overview, we show several CCA subtypes according to the location of CCA, including intrahepatic (iCCA), perihepatic (pCCA) and extrahepatic (eCCA). We collected previous studies to indicate various mechanisms and experiments in patients’ blood, urine and primary tissue specimens. This schematic model illustrates that omics-based approaches can be used and integrated for further CCA basic and clinical research. In this topic, we input genomics, epigenomics, transcriptomics, proteomics and metabolomics observations for readers.
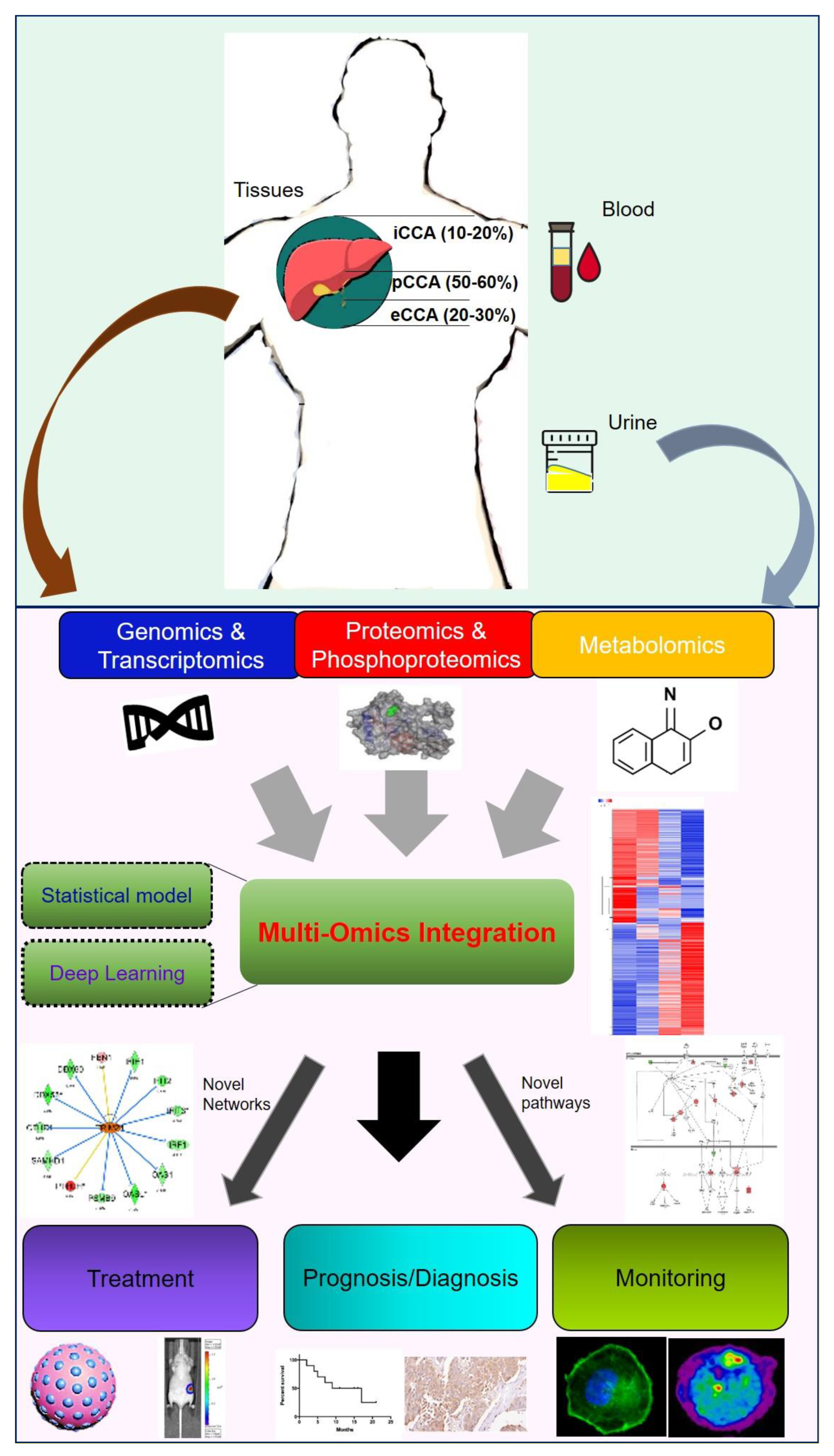

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Genetic events of cholangiocarcinoma (CCA).
| Copy-Number Alteration (CNA) | ||||
| Gene | Cytoband | CAN | # | Frequency (%) |
| CDKN2A | 9p21.3 | HOMDEL | 21 | 9.1 |
| CDKN2B | 9p21.3 | HOMDEL | 16 | 6.9 |
| MCL1 | 1q21.2 | AMP | 11 | 4.8 |
| CCND1 | 11q13.3 | AMP | 8 | 3.5 |
| FDF4 | 11q13.3 | AMP | 8 | 3.5 |
| MDM2 | 12q15 | AMP | 8 | 3.5 |
| RIT1 | 1q22 | AMP | 8 | 3.5 |
| FDF19 | 11q13.3 | AMP | 8 | 3.5 |
| HIST2H3C | 1q21.2 | AMP | 6 | 3.4 |
| HIST2H3D | 1q21.2 | AMP | 6 | 3.4 |
| ERBB2 | 17q12 | AMP | 7 | 3 |
| FGF3 | 11q13.3 | AMP | 7 | 3 |
| MUC1 | 1q22 | AMP | 7 | 3 |
| MYC | 8q24.21 | AMP | 7 | 3 |
| NOTCH2 | 1p12 | AMP | 7 | 3 |
| Mutated Genes | ||||
| Gene | #Mut | Frequency (%) | ||
| TP53 | 114 | 27.3 | ||
| IDH1 | 64 | 16.2 | ||
| ARID1A | 69 | 16.2 | ||
| KRAS | 49 | 12.4 | ||
| BAP1 | 49 | 11.9 | ||
| PBRM1 | 37 | 9.3 | ||
| KMT2C | 28 | 7.1 | ||
| HLA-A | 37 | 6.7 | ||
| SMAD4 | 27 | 6.3 | ||
| ATM | 27 | 6.1 | ||
| Fusion Genes | ||||
| Gene | #Mut | Frequency (%) | ||
| FDFR2 | 19 | 4.8 | ||
| RAD21 | 1 | 0.3 | ||
| BRCA1 | 1 | 0.3 | ||
| BRAF | 1 | 0.3 | ||
| ESR1 | 1 | 0.3 | ||
| MAP3K1 | 1 | 0.3 | ||
| NOTCH2 | 1 | 0.3 | ||
| PIK3C2G | 1 | 0.3 | ||
| MAP2K1 | 1 | 0.3 | ||
| RB1 | 1 | 0.3 | ||
Table 2.
Most significant miRNAs in CCA is related to survival rate or tumorigenesis. All candidates from WashU Pan-Cancer miRNome Atlas (OncomiR) website. Significance in tumor development was determined through paired Student’s t-test, for survival, unpaired Student’s t-test and univariate Cox proportional hazards analysis between living and decreased patients.
Table 2.
Most significant miRNAs in CCA is related to survival rate or tumorigenesis. All candidates from WashU Pan-Cancer miRNome Atlas (OncomiR) website. Significance in tumor development was determined through paired Student’s t-test, for survival, unpaired Student’s t-test and univariate Cox proportional hazards analysis between living and decreased patients.
| miRNAs Significantly Associated with Survival in CHOL | |||
| miRNA Name | Z-Score | Log Rank p-Value | Upregulated in: |
| hsa-miR-802 | 0.000 | 3.02 × 10−3 | Living |
| hsa-miR-500b-5p | 2.718 | 3.57 × 10−3 | Living |
| hsa-miR-500a-5p | 2.674 | 4.27 × 10−3 | Living |
| hsa-miR-202-5p | 0.000 | 4.91 × 10−3 | Living |
| hsa-miR-551b-3p | 1.746 | 5.57 × 10−3 | Living |
| hsa-miR-129-5p | 2.166 | 5.79 × 10−3 | Living |
| hsa-miR-3161 | 3.025 | 9.51 × 10−3 | Decreased |
| hsa-miR-3199 | 0.173 | 1.50 × 10−2 | Living |
| hsa-miR-1228-5p | 0.000 | 2.03 × 10−2 | Living |
| hsa-miR-10b-3p | 1.970 | 2.40 × 10−2 | Living |
| miRNAs Associated with Tumorigenesis of CHOL | |||
| miRNA Name | t-Test p-Value | t-Test FDR | Upregulated in: |
| hsa-miR-183-5p | 3.84 × 10−9 | 2.16 × 10−6 | Tumor |
| hsa-miR-182-5p | 1.15 × 10−8 | 3.25 × 10−6 | Tumor |
| hsa-miR-194-3p | 2.88 × 10−8 | 5.41 × 10−6 | Normal |
| hsa-miR-125b-2-3p | 1.35 × 10−7 | 1.85 × 10−5 | Normal |
| hsa-let-7c-5p | 1.71 × 10−7 | 1.85 × 10−5 | Normal |
| hsa-miR-378a-3p | 1.97 × 10−7 | 1.85 × 10−5 | Normal |
| hsa-miR-92b-3p | 2.60 × 10−7 | 2.09 × 10−5 | Tumor |
| hsa-miR-1258 | 3.43 × 10−7 | 2.41 × 10−5 | Normal |
| hsa-miR-378c | 4.13 × 10−7 | 2.59 × 10−5 | Normal |
| hsa-miR-23c | 8.50 × 10−7 | 4.36 × 10−5 | Normal |
Table 3.
Most significant LncRNAs/CircRNAs are related to the tumorigenesis of CCA. All candidates from LncRNADisease website (version 2.0).
Table 3.
Most significant LncRNAs/CircRNAs are related to the tumorigenesis of CCA. All candidates from LncRNADisease website (version 2.0).
| LncRNAs/CircRNAs Dependency of CCA | |||
|---|---|---|---|
| LncRNAs Symbol | Disease Name | Confidence Score | Database ID |
| SPRY4-IT1 | CCA | 0.9820 | LDA0003741 |
| CPS1-IT1 | iCCA | 0.9820 | LDA0004726 |
| MALAT1 | CCA | 0.9526 | LDA0003742 |
| NEAT1 | CCA | 0.9526 | LDA0003743 |
| SOX2-OT | CCA | 0.9462 | LDA0003746 |
| AFAP1-AS1 | CCA | 0.8808 | LDA0003736 |
| UCA1 | CCA | 0.8808 | LDA0003749 |
| CCAT1 | iCCA | 0.8808 | LDA0004724 |
| H19 | CCA | 0.8785 | LDA0003740 |
| HULC | CCA | 0.8785 | LDA0003741 |
| PCAT1 | CCA | 0.8785 | LDA0003745 |
| CCAT2 | CCA | 0.7311 | LDA0003737 |
| PANDAR | CCA | 0.7311 | LDA0003744 |
| TUG1 | CCA | 0.7311 | LDA0003748 |
| PCAT1 | eCCA | 0.7311 | LDA0004199 |
| TLINC | iCCA | 0.7311 | LDA0004727 |
| TUG1 | iCCA | 0.7311 | LDA0004728 |
| CDR1-AS | CCA | 0.7311 | LDA0178708 |
| hsa-circ_0001649 | CCA | 0.7311 | LDA0178709 |
| CCAT2 | iCCA | 0.6606 | LDA0004725 |
| ENST00000517758.1 | CCA | 0.5483 | LDA0003738 |
| ENST00000588480.1 | CCA | 0.5483 | LDA0003739 |
The confidence scores can be calculated as follows: CS = 1 − (1 − ).
Table 4.
Most significant survival genes in CCA. All candidates from gene expression profiling interactive analysis (GEPIA) website. Log-rank test (Mantel-Cox test), group cutoff: median.
Table 4.
Most significant survival genes in CCA. All candidates from gene expression profiling interactive analysis (GEPIA) website. Log-rank test (Mantel-Cox test), group cutoff: median.
| Overall Survival | ||
| Gene Symbol | Gene ID | pValue |
| CTD-2033C11.1 | ENSG00000269961.1 | 4.36 × 10−5 |
| AD001527.7 | ENSG00000270760.1 | 1.46 × 10−4 |
| FAM86KP | ENSG00000163612.10 | 4.47 × 10−4 |
| RP11-209M4.1 | ENSG00000267253.1 | 9.00 × 10−4 |
| PTGER3 | ENSG00000050628.20 | 9.41 × 10−4 |
| AP003774.6 | ENSG00000231680.1 | 9.95 × 10−4 |
| FAM177A1 | ENSG00000151327.12 | 1.00 × 10−3 |
| RP11-574F21.2 | ENSG00000228606.1 | 1.05 × 10−3 |
| CXCL17 | ENSG00000189377.8 | 1.14 × 10−3 |
| FUT4 | ENSG00000196371.3 | 1.27 × 10−3 |
| Disease-Free Survival | ||
| Gene Symbol | Gene ID | pValue |
| AC104654.2 | ENSG00000234362.5 | 1.31 × 10−6 |
| AP003774.6 | ENSG00000231680.1 | 5.59 × 10−6 |
| NADK | ENSG00000008130.15 | 3.05 × 10−5 |
| AGAP2 | ENSG00000135439.11 | 3.59 × 10−5 |
| NRXN2 | ENSG00000110076.18 | 4.80 × 10−5 |
| RP1-28O10.1 | ENSG00000227591.5 | 4.81 × 10−5 |
| CPSF3L | ENSG00000127054.18 | 7.45 × 10−5 |
| TPRG1L | ENSG00000158109.14 | 8.32 × 10−5 |
| CDC40 | ENSG00000168438.14 | 8.54 × 10−5 |
| KLHL34 | ENSG00000185915.5 | 8.86 × 10−5 |
Table 5.
Inhibitors and their related clinical trials for CCA.
| Compound | Target | Clinical Trial Number | Phase | Status |
|---|---|---|---|---|
| NVP-BGJ398 (infigratinib) | FGFR | NCT02150967 | II | Recruiting |
| JNJ-42756493 (Erdafitinib) | FGFR | NCT02699606 | II | Recruiting |
| ARQ 087 (derazantinib) | FGFR | NCT017529520 | I + II | Active |
| TAS-120 | FGFR | NCT02052778 | I + II | Active |
| CH5183284 (debio 1347) | FGFR | NCT03834220 | II | Active |
| Ponatinib | FGFR2 | NCT02265341 | II | Completed |
| FPA144 (bemarituzumab) | FGFR2 | N/A | ||
| Pemazyre (pemigatinib) | FGFR2 | NCT03656536 | III | Recruiting |
| AG-120 (Ivosidenib) | IDH1 | NCT02073994 | I | Active |
| Trametinib | MEK | NCT02070549/NCT02042443 | I/II | Recruiting/Completed |
| Pazopanib | MEK/VEGFR/PDGFR/REF | NCT01855724 | II | Terminated |
| Pembrolizumab | PD-1 | NCT03111732 | II | Recruiting |
| Regorafenib | MEK/VEGFR/PDGFR/REF | NCT02115542 | II | Active |
| AGI-5198 | IDH1 | N/A | ||
| AGI-6780 | IDH2 | N/A | ||
| AG-221 (enasidenib) | IDH2 | NCT02273739 | I + II | Completed |
| Isoquinoline | PKA | N/A | ||
| S63845 | MCL1 | N/A | ||
| AZD1480 | JAK | N/A | ||
| AZD6244 (selumetinib) | MEK1/2 | NCT00553332 | II | Completed |
| BKM120 (buparlisib) | PI3K | NCT01501604 | II | Withdrawn |
| Amatuximab | Mesothelin | NCT01766219 | I + II | Completed |
| Azacitidine | DNMT | N/A | ||
| Decitabine | DNMT | N/A | ||
| Vorinostat | HDAC | N/A | ||
| Romidepsin | HDAC | N/A | ||
| Tivantinib | MET | N/A | ||
| Cabozantinib | MET | NCT01954745 | II | Completed |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Chang, Y.-C.; Chen, M.-H.; Yeh, C.-N.; Hsiao, M. Omics-Based Platforms: Current Status and Potential Use for Cholangiocarcinoma. Biomolecules 2020, 10, 1377. https://doi.org/10.3390/biom10101377
AMA Style
Chang Y-C, Chen M-H, Yeh C-N, Hsiao M. Omics-Based Platforms: Current Status and Potential Use for Cholangiocarcinoma. Biomolecules. 2020; 10(10):1377. https://doi.org/10.3390/biom10101377
Chicago/Turabian StyleChang, Yu-Chan, Ming-Huang Chen, Chun-Nan Yeh, and Michael Hsiao. 2020. "Omics-Based Platforms: Current Status and Potential Use for Cholangiocarcinoma" Biomolecules 10, no. 10: 1377. https://doi.org/10.3390/biom10101377
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.