Distinct Peculiarities of In Planta Synthesis of Isoprenoid and Aromatic Cytokinins

 , , and
, , and
Abstract
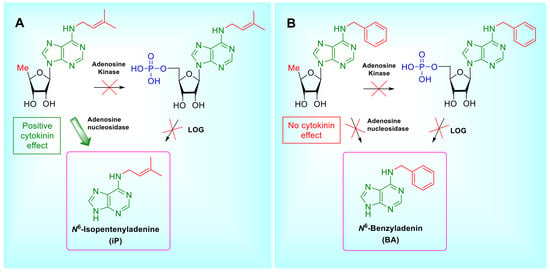
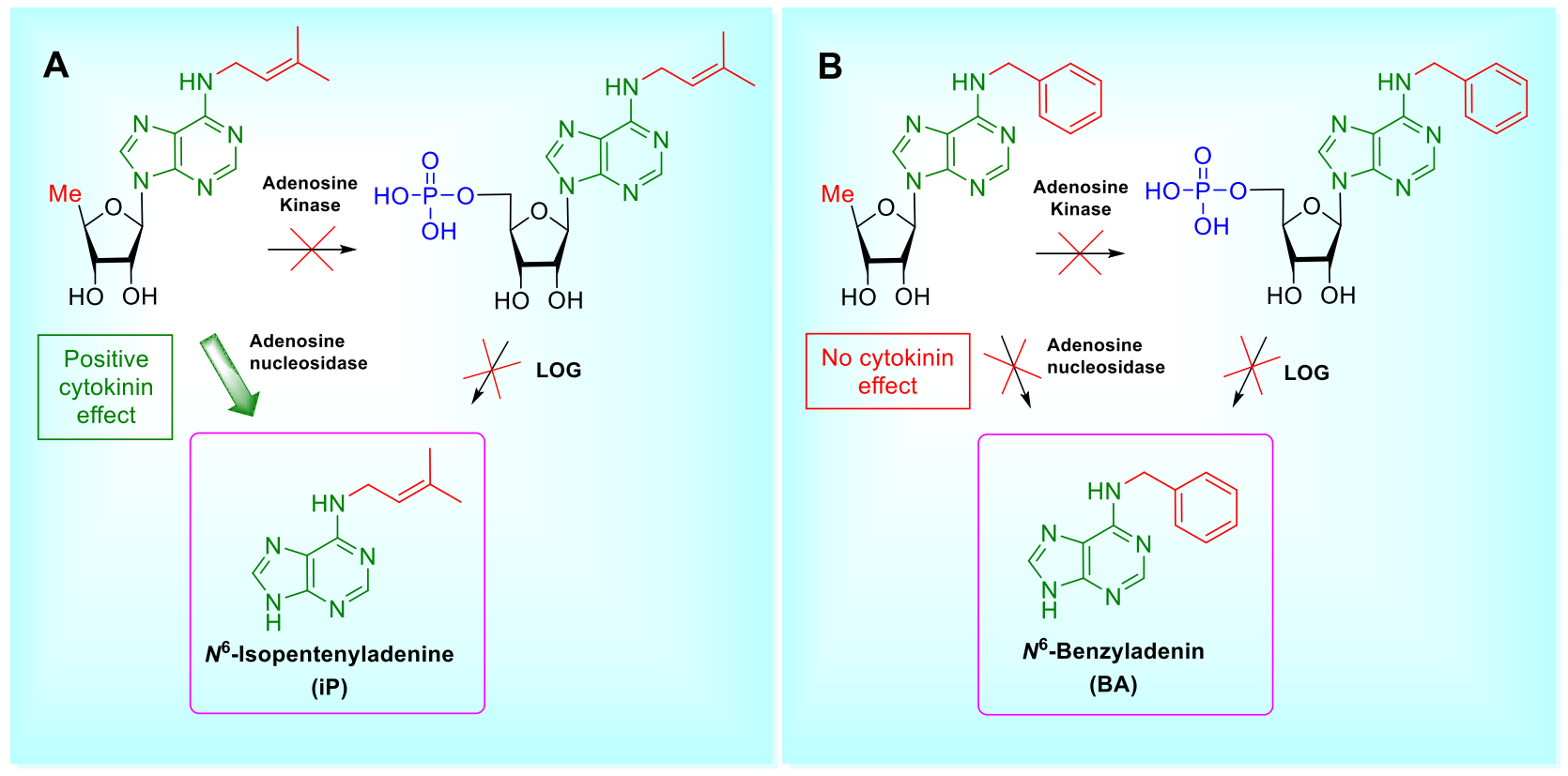
1. Introduction
2. Materials and Methods
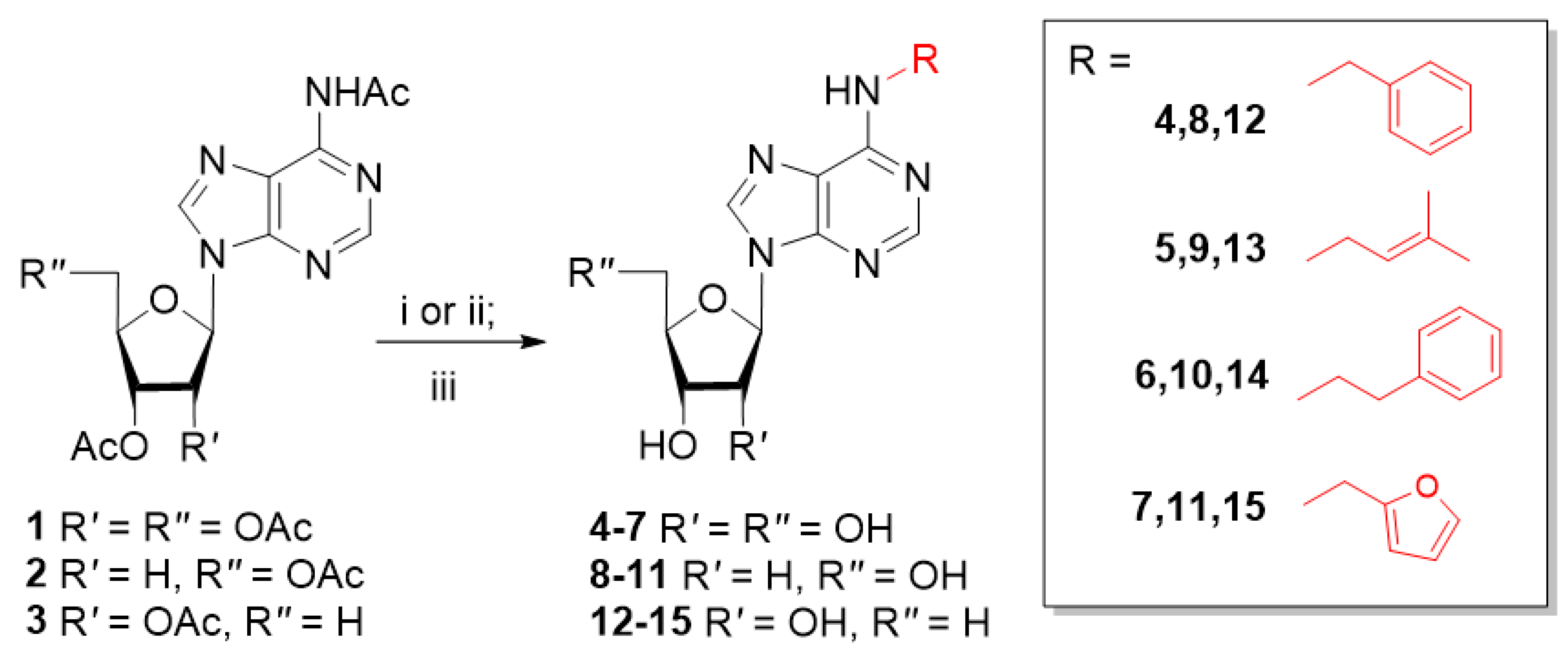
2.1. Synthesis
2.1.1. General
2.1.2. Typical Procedure for Preparation of Nucleosides by Alkylation with Alkyl Halides
2.1.3. Typical Procedure for Preparation of Nucleosides by Mitsunobu Reaction with Alcohols
2.2. Cytokinin Activity Assays
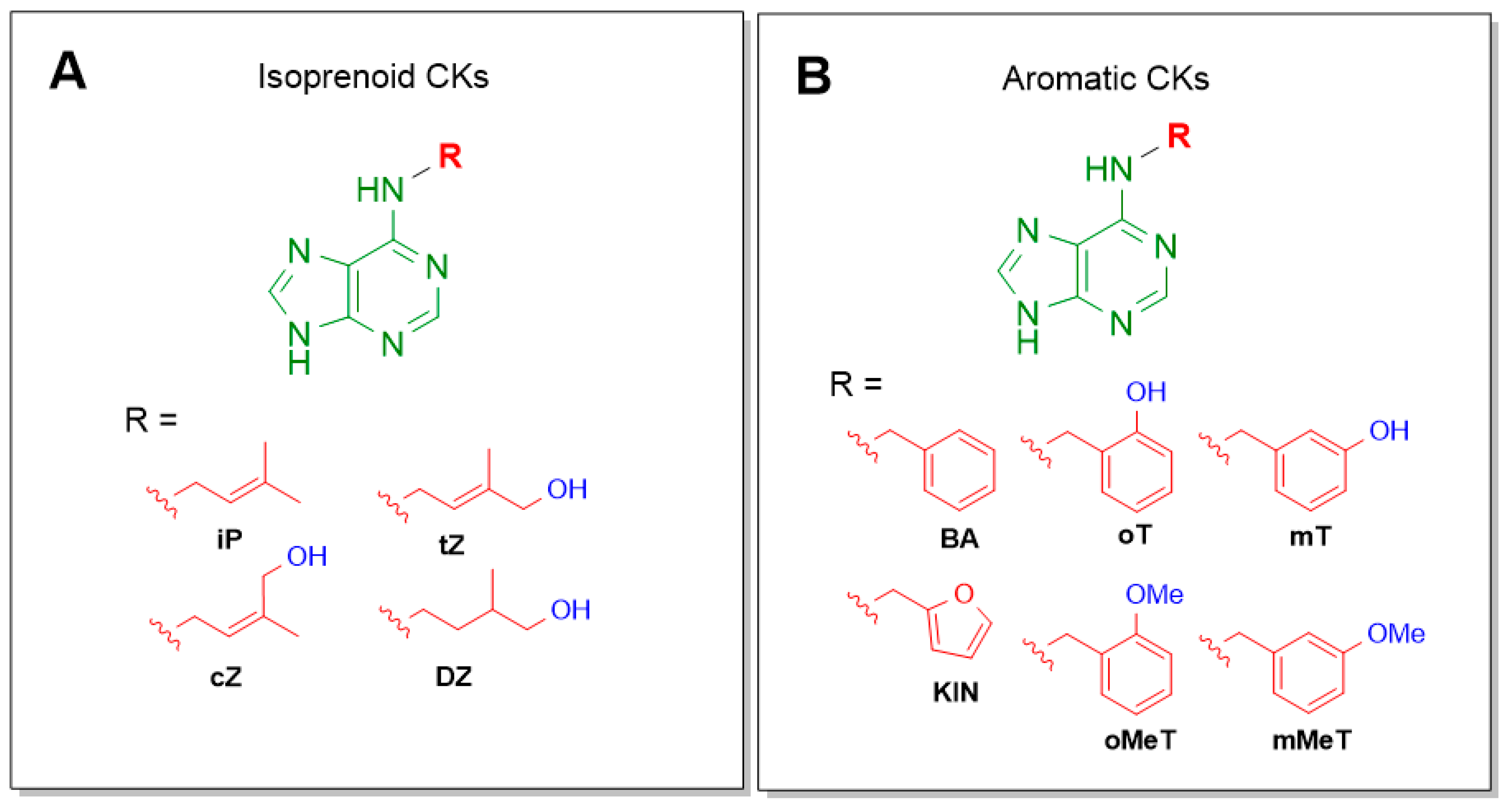
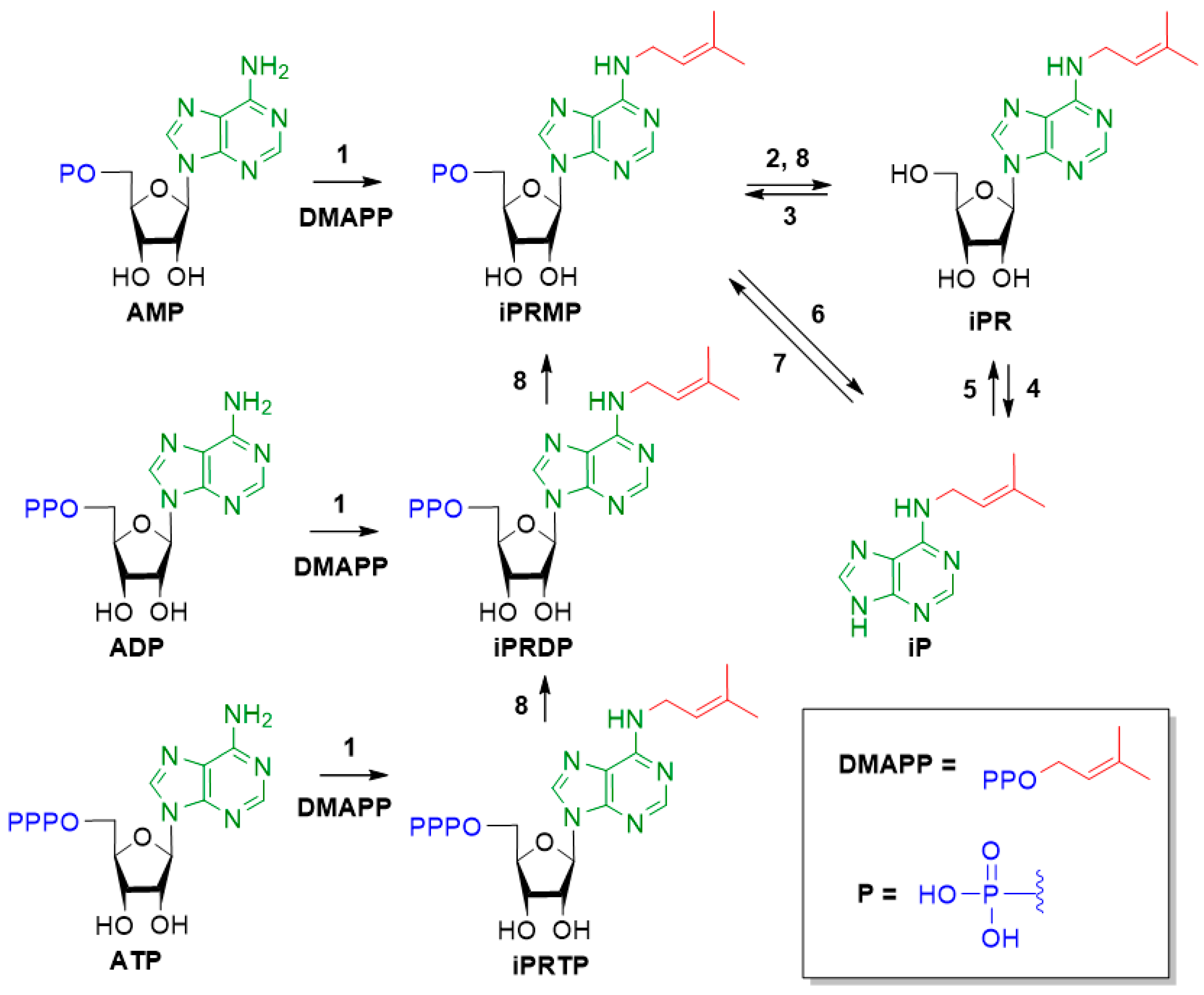
3. Results and Discussion
3.1. Chemistry
3.2. Cytokinin Activity
3.3. Analysis of Results with Arabidopsis Bioassay
3.4. Analysis of Results with Amaranthus Bioassay
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Sakakibara, H. Cytokinins: Activity, biosynthesis, and translocation. Annu. Rev. Plant Biol. 2006, 57, 431–449. [Google Scholar] [CrossRef] [PubMed]
- Romanov, G.A. How do cytokinins affect the cell? Rus. J. Plant Physiol. 2009, 56, 268–290. [Google Scholar] [CrossRef]
- Miller, C.; Skoog, F.; Saltza, M.; Strong, M. Kinetin, a cell division factor from deoxyribonucleic acid. J. Am. Chem. Soc. 1955, 77, 1329–1334. [Google Scholar] [CrossRef]
- Plihalova, L.; Vylíčilová, H.; Doležal, K.; Zahajska, L.; Zatloukal, M.; Strnad, M. Synthesis of aromatic cytokinins for plant biotechnology. New Biotech. 2016, 33, 614–624. [Google Scholar] [CrossRef]
- Kamínek, M. Tracking the story of cytokinin research. J. Plant Growth Regul. 2015, 34, 723–739. [Google Scholar] [CrossRef]
- Lomin, S.N.; Krivosheev, D.M.; Steklov, M.Y.; Arkhipov, D.V.; Osolodkin, D.I.; Schmülling, T.; Romanov, G.A. Plant membrane assays with cytokinin receptors underpin the unique role of free cytokinin bases as biologically active ligands. J. Exp. Bot. 2015, 66, 1851–1863. [Google Scholar] [CrossRef]
- Savelieva, E.M.; Oslovsky, V.E.; Karlov, D.S.; Kurochkin, N.N.; Getman, I.A.; Lomin, S.N.; Sidorov, G.V.; Mikhailov, S.N.; Osolodkin, D.I.; Romanov, G.A. Cytokinin activity of N6-benzyladenine derivatives assayed by interaction with the receptors in planta, in vitro, and in silico. Phytochemistry 2018, 149, 161–177. [Google Scholar] [CrossRef]
- Sakakibara, H. Cytokinin biosynthesis and metabolism. In Plant Hormones, 3rd ed.; Davies, P.J., Ed.; Springer: Dordrecht, The Netherlands, 2010; pp. 95–114. [Google Scholar]
- Komada-Nobusada, T.; Sakakibara, H. Molecular basis for cytokinin biosynthesis. Phytochemistry 2009, 70, 444–449. [Google Scholar] [CrossRef]
- Tokunaga, H.; Kojima, M.; Kuroha, T.; Ishida, T.; Sugimoto, K.; Kiba, T.; Sakakibara, H. Arabidopsis lonely guy (LOG) multiple mutants reveal a central role of the LOG-dependent pathway in cytokinin activation. Plant J. 2012, 69, 355–365. [Google Scholar] [CrossRef]
- Osugi, A.; Sakakibara, H. Q&A: How do plants respond to cytokinins and what is their importance? BMC Biol. 2015, 13, 102. [Google Scholar]
- Jordheim, L.P.; Durantel, D.; Zoulim, F.; Dumontet, C. Advances in the development of nucleoside and nucleotide analogues for cancer and viral diseases. Nat. Rev. Drug Discov. 2013, 12, 447–464. [Google Scholar] [CrossRef] [PubMed]
- Drenichev, M.S.; Oslovsky, V.E.; Tararov, V.I.; Mikhailov, S.N. Synthesis of N6-substituted adenosines as cytokinin nucleosides. Curr. Protoc. Nucleic Acid Chem. 2018, 72, 14.15.1–14.15.16. [Google Scholar] [CrossRef] [PubMed]
- Drenichev, M.S.; Oslovsky, V.E.; Sun, L.; Tijsma, A.; Kurochkin, N.N.; Tararov, V.I.; Chizhov, A.O.; Neyts, J.; Pannecouque, C.; Leyssen, P.; et al. Modification of the length and structure of the linker of N6-benzyladenosine modulates its selective antiviral activity against enterovirus 71. Eur. J. Med. Chem. 2016, 111, 84–94. [Google Scholar] [CrossRef] [PubMed]
- Riefler, M.; Novak, O.; Strnad, M.; Schmülling, T. Arabidopsis cytokinin receptor mutants reveal functions in shoot growth, leaf senescence, seed size, germination, root development, and cytokinin metabolism. Plant Cell 2006, 18, 40–54. [Google Scholar] [CrossRef] [PubMed]
- Stolz, A.; Riefler, M.; Lomin, S.N.; Achazi, K.; Romanov, G.A.; Schmülling, T. The specificity of cytokinin signalling in Arabidopsis thaliana is mediated by differing ligand affinities and expression profiles of the receptors. Plant J. 2011, 67, 157–168. [Google Scholar] [CrossRef] [PubMed]
- Romanov, G.A.; Kieber, J.J.; Schmülling, T. A rapid cytokinin response assay in Arabidopsis indicates a role for phospholipase D in cytokinin signalling. FEBS Lett. 2002, 515, 39–43. [Google Scholar] [CrossRef]
- Zvereva, S.D.; Romanov, G.A. Reporter genes for plant genetic engineering: Characteristics and detection. Russ. J. Plant Physiol. 2000, 47, 424–432. [Google Scholar]
- Romanov, G.A.; Getman, I.A.; Schmülling, T. Investigation of early cytokinin effects in a rapid Amaranthus seedling test. Plant Growth Regul. 2000, 34, 337–344. [Google Scholar] [CrossRef]
- Biddington, N.L.; Thomas, T.H. A modified Amaranthus betacyanin bioassay for the rapid determination of cytokinins in plant extracts. Planta 1973, 111, 183–186. [Google Scholar] [CrossRef]
- Wang, Y.; Hogenkamp, H.P.; Long, R.A.; Revankar, G.R.; Robins, R.K. A convenient synthesis of 5′-deoxyribonucleosides. Carbohydr. Res. 1977, 59, 449–457. [Google Scholar] [CrossRef]
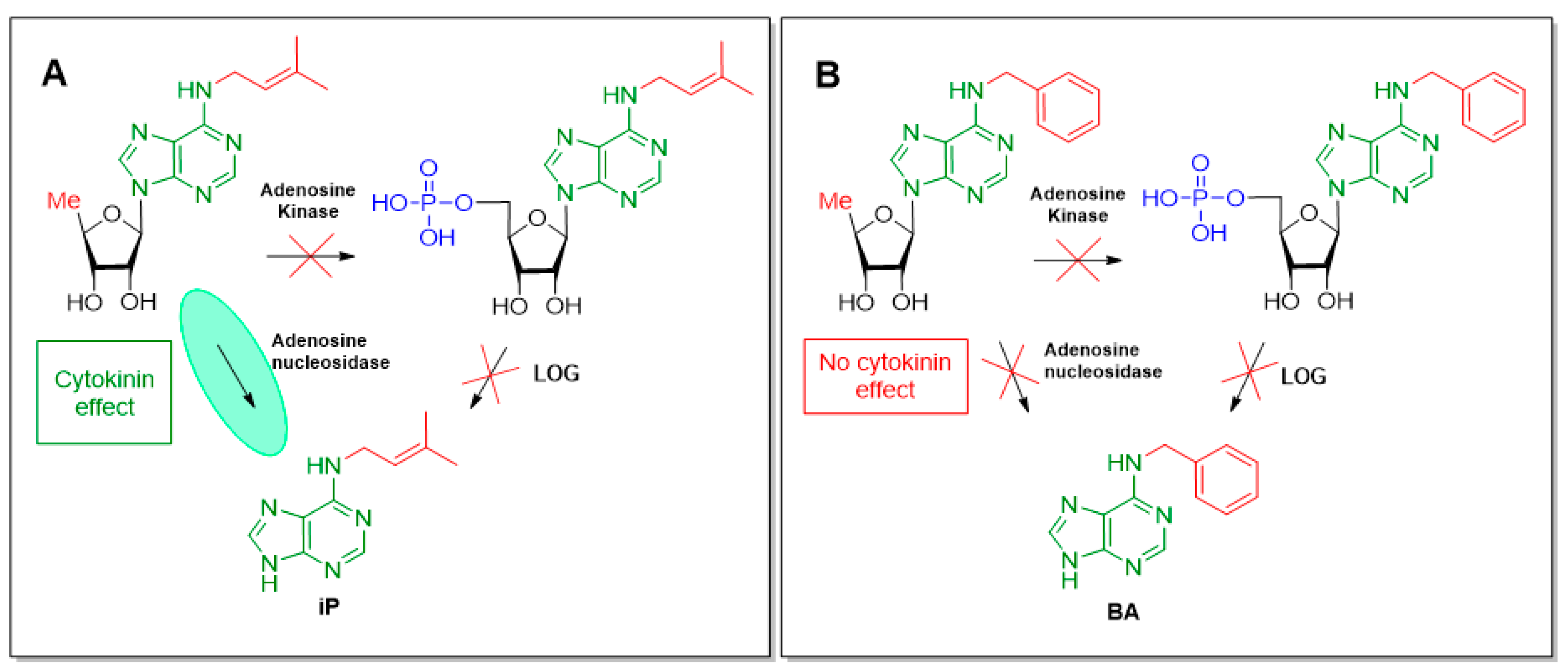
- Oslovsky, V.E.; Savelieva, E.M.; Drenichev, M.S.; Romanov, G.A.; Mikhailov, S.N. Comparative analysis of the biosynthesis of isoprenoid and aromatic cytokinins. Dokl. Biochem. Biophys. 2019, 488, 346–349. [Google Scholar] [CrossRef] [PubMed]
- Ogawa, J.; Takeda, S.; Xie, S.X.; Hatanaka, H.; Ashikari, T.; Amachi, T.; Shimizu, S. Purification, characterization, and gene cloning of purine nucleosidase from Ochrobactrum anthropi. Appl. Environ. Microbiol. 2001, 67, 1783–1787. [Google Scholar] [CrossRef] [PubMed]
- Abusamhadneh, E.; McDonald, N.E.; Kline, P.C. Isolation and characterization of adenosine nucleosidase from yellow lupin (Lupinus luteus). Plant Sci. 2000, 153, 25–32. [Google Scholar] [CrossRef]
- Chen, C.M.; Kristopeit, S.M. Metabolism of cytokinin: Deribosylation of cytokinin ribonucleoside by adenosine nucleosidase from wheat germ cells. Plant Physiol. 1981, 68, 1020–1023. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | N6-Substituent | Conc. | WT | AHK2 | AHK3 | AHK4 |
|---|---|---|---|---|---|---|
| Ribonucleosides | ||||||
| 4 * |  | 10−5 M | 116 ± 5 | 90 ± 5 | 59 ± 5 | 43 ± 1 |
| 10−6 M | 75 ± 3 | 94 ± 9 | 77 ± 4 | 51 ± 3 | ||
| 5 * |  | 10−5 M | 113 ± 6 | 94 ± 6 | 55 ± 4 | 40 ± 1 |
| 10−6 M | 52 ± 6 | 114 ± 5 | 80 ± 4 | 40 ± 5 | ||
| 6 * |  | 10−5 M | 53 ± 3 | 75 ± 8 | 32 ± 4 | 19 ± 1 |
| 10−6 M | 28 ± 3 | 38 ± 0.5 | 12 ± 1 | 16 ± 0.1 | ||
| 7 |  | 10−5 M | 78 ± 4 | 90 ± 6 | 61 ± 4 | 96 ± 12 |
| 10−6 M | 41 ± 11 | 59 ± 0.3 | 63 ± 7 | 55 ± 2.5 | ||
| 2′-Deoxyribonucleosides | ||||||
| 8 | | 10−5 M | 15 ± 1 | 32 ± 6 | 8 ± 0.3 | 13 ± 0.1 |
| 10−6 M | 10 ± 0.5 | 4 ± 0.2 | 7 ± 1 | 2 ± 0.1 | ||
| 9 | | 10−5 M | 105 ± 4 | 62 ± 10 | 51 ± 4 | 68 ± 5 |
| 10−6 M | 29 ± 2 | 34 ± 4 | 40 ± 4 | 7 ± 1 | ||
| 10 | | 10−5 M | 8 ± 2 | 4 ± 0.1 | 4 ± 0.1 | 3.5 ± 0.1 |
| 10−6 M | 8 ± 2 | 4 ± 0.1 | 3 ± 1 | 2 ± 0.3 | ||
| 11 | | 10−5 M | 42 ± 2 | 70 ± 1 | 4 ± 1 | 27 ± 5 |
| 10−6 M | 21 ± 2 | 33 ± 1.5 | 5 ± 0.2 | 25 ± 1 | ||
| 5′-Deoxyribonucleosides | ||||||
| 12 * | | 10−5 M | 22 ± 1 | 12 ± 1 | 7 ± 0.2 | 6 ± 0.4 |
| 10−6 M | 9 ± 1 | 3 ± 0.6 | 4 ± 1 | 1.5 ± 0.1 | ||
| 13 * | | 10−5 M | 87 ± 7 | 67 ± 9 | 34 ± 5 | 51 ± 2 |
| 10−6 M | 41 ± 6 | 20 ± 2 | 9 ± 1 | 11 ± 1 | ||
| 14 * | | 10−5 M | 15 ± 3 | 10 ± 1 | 14 ± 5 | 4 ± 0.2 |
| 10−6 M | 7 ± 1 | 5 ± 1 | 10 ± 1 | 4 ± 0.2 | ||
| 15 | | 10−5 M | 10 ± 1.1 | 13 ± 1 | 13 ± 2 | 12 ± 0.5 |
| 10−6 M | 8 ± 3 | 7 ± 0.5 | 7 ± 1 | 7 ± 1 | ||
| Compound | N6-Substituent | Conc. | Relative Activity, % |
|---|---|---|---|
| Ribonucleosides | |||
| 4 | | 10−6 M | 64.38 ± 10.94 |
| 5 | | 10−6 M | 78.29 ± 13.29 |
| 6 | | 10−6 M | 18.69 ± 8.22 |
| 7 | | 10−6 M | 21.28 ± 1.01 |
| 2′-Deoxyribonucleosides | |||
| 8 | | 10−6 M | 24.08 ± 6.94 |
| 9 | | 10−6 M | 12.64 ± 2.01 |
| 10 | | 10−6 M | 0 |
| 11 | | 10−6 M | 21.8 ± 5.0 |
| 5′-Deoxyribonucleosides | |||
| 12 | | 10−6 M | 0 |
| 13 | | 10−6 M | 0 |
| 14 | | 10−6 M | 0 |
| 15 | | 10−6 M | 9.52 ± 0.33 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Oslovsky, V.E.; Savelieva, E.M.; Drenichev, M.S.; Romanov, G.A.; Mikhailov, S.N. Distinct Peculiarities of In Planta Synthesis of Isoprenoid and Aromatic Cytokinins. Biomolecules 2020, 10, 86. https://doi.org/10.3390/biom10010086
Oslovsky VE, Savelieva EM, Drenichev MS, Romanov GA, Mikhailov SN. Distinct Peculiarities of In Planta Synthesis of Isoprenoid and Aromatic Cytokinins. Biomolecules. 2020; 10(1):86. https://doi.org/10.3390/biom10010086
Chicago/Turabian StyleOslovsky, Vladimir E., Ekaterina M. Savelieva, Mikhail S. Drenichev, Georgy A. Romanov, and Sergey N. Mikhailov. 2020. "Distinct Peculiarities of In Planta Synthesis of Isoprenoid and Aromatic Cytokinins" Biomolecules 10, no. 1: 86. https://doi.org/10.3390/biom10010086
APA StyleOslovsky, V. E., Savelieva, E. M., Drenichev, M. S., Romanov, G. A., & Mikhailov, S. N. (2020). Distinct Peculiarities of In Planta Synthesis of Isoprenoid and Aromatic Cytokinins. Biomolecules, 10(1), 86. https://doi.org/10.3390/biom10010086