Imaging Mass Spectrometry: A New Tool to Assess Molecular Underpinnings of Neurodegeneration

 ,
,
Abstract
1. Introduction
2. Imaging Mass Spectrometry: Advantages and Disadvantages
2.1. Advantages
2.2. Disadvantages
3. Ionization Methods
3.1. MALDI
3.2. DESI
3.3. SIMS
4. IMS Analysis
5. Neurodegenerative Disorders
5.1. Alzheimer’s Disease (AD)
5.2. Parkinson’s Disease (PD)
5.3. Huntington’s Disease (HD)
5.4. Multiple Sclerosis (MS)
5.5. Amyotrophic Lateral Sclerosis (ALS)
6. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Watrous, J.D.; Alexandrov, T.; Dorrestein, P.C. The evolving field of imaging mass spectrometry and its impact on future biological research. J. Mass Spectrom. 2011, 46, 209–222. [Google Scholar] [CrossRef] [PubMed]
- Seeley, E.H.; Schwamborn, K.; Caprioli, R.M. Imaging of Intact Tissue Sections: Moving beyond the Microscope. J. Boil. Chem. 2011, 286, 25459–25466. [Google Scholar] [CrossRef] [PubMed]
- Chaurand, P.; Cornett, D.S.; Angel, P.M.; Caprioli, R.M. From whole-body sections down to cellular level, multiscale imaging of phospholipids by MALDI mass spectrometry. Mol. Cell Proteom. 2011, 10. [Google Scholar] [CrossRef] [PubMed]
- Castellino, S.; Groseclose, M.R.; Wagner, D. MALDI imaging mass spectrometry: Bridging biology and chemistry in drug development. Bioanalysis 2011, 3, 2427–2441. [Google Scholar] [CrossRef]
- Caprioli, R.M.; Farmer, T.B.; Gile, J. Molecular Imaging of Biological Samples: Localization of Peptides and Proteins Using MALDI-TOF MS. Anal. Chem. 1997, 69, 4751–4760. [Google Scholar] [CrossRef]
- Arnold, J.M.; Choi, W.T.; Sreekumar, A.; Maletić-Savatić, M. Analytical strategies for studying stem cell metabolism. Front. Boil. 2015, 10, 141–153. [Google Scholar] [CrossRef][Green Version]
- Miura, D.; Fujimura, Y.; Yamato, M.; Hyodo, F.; Utsumi, H.; Tachibana, H.; Wariishi, H. Ultrahighly Sensitive in Situ Metabolomic Imaging for Visualizing Spatiotemporal Metabolic Behaviors. Anal. Chem. 2010, 82, 9789–9796. [Google Scholar] [CrossRef]
- Kumar, A.; Agarwal, S.; Heyman, J.A.; Matson, S.; Heidtman, M.; Piccirillo, S.; Umansky, L.; Drawid, A.; Jansen, R.; Liu, Y.; et al. Subcellular localization of the yeast proteome. Genome Res. 2002, 16, 707–719. [Google Scholar] [CrossRef]
- Caprioli, R.M. Imaging mass spectrometry: Molecular microscopy for enabling a new age of discovery. Proteomics 2014, 14, 807–809. [Google Scholar] [CrossRef]
- Groseclose, M.R.; Laffan, S.B.; Frazier, K.S.; Hughes-Earle, A.; Castellino, S. Imaging MS in Toxicology: An Investigation of Juvenile Rat Nephrotoxicity Associated with Dabrafenib Administration. J. Am. Soc. Mass Spectrom. 2015, 26, 887–898. [Google Scholar] [CrossRef]
- Groseclose, M.R.; Castellino, S. A Mimetic Tissue Model for the Quantification of Drug Distributions by MALDI Imaging Mass Spectrometry. Anal. Chem. 2013, 85, 10099–10106. [Google Scholar] [CrossRef]
- Nilsson, A.; Peric, A.; Strimfors, M.; Goodwin, R.J.A.; Hayes, M.A.; Andrén, P.E.; Hilgendorf, C. Mass Spectrometry Imaging proves differential absorption profiles of well-characterised permeability markers along the crypt-villus axis. Sci. Rep. 2017, 7, 6352. [Google Scholar] [CrossRef] [PubMed]
- Goodwin, R.; Bunch, J.; McGinnity, D. Mass Spectrometry Imaging in Oncology Drug Discovery. Adv. Cancer Res. 2017, 134, 133–171. [Google Scholar]
- Choi, W.T.; Tosun, M.; Jeong, H.-H.; Karakas, C.; Semerci, F.; Liu, Z.; Maletić-Savatić, M. Metabolomics of mammalian brain reveals regional differences. BMC Syst. Boil. 2018, 12, 127. [Google Scholar] [CrossRef] [PubMed]
- Reyzer, M.L.; Caprioli, R.M. MALDI-MS-based imaging of small molecules and proteins in tissues. Curr. Opin. Chem. Boil. 2007, 11, 29–35. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; MacKenzie, K.R.; Putluri, N.; Maletić-Savatić, M.; Bellen, H.J. The glia-neuron lactate shuttle and elevated ROS promote lipid synthesis in neurons and lipid droplet accumulation in glia via APOE/D. Cell Metab. 2017, 26, 719–737. [Google Scholar] [CrossRef]
- Botas, A.; Campbell, H.M.; Han, X.; Maletic-Savatic, M. Metabolomics of Neurodegenerative Diseases. Int. Rev. Neurobiol. 2015, 122, 53–80. [Google Scholar] [PubMed]
- Calvano, C.D.; Palmisano, F.; Cataldi, T.R. Understanding neurodegenerative disorders by MS-based lipidomics. Bioanalysis 2018, 10, 787–790. [Google Scholar] [CrossRef] [PubMed]
- Sugiura, Y.; Shimma, S.; Konishi, Y.; Yamada, M.K.; Setou, M. Imaging Mass Spectrometry Technology and Application on Ganglioside Study; Visualization of Age-Dependent Accumulation of C20-Ganglioside Molecular Species in the Mouse Hippocampus. PLoS ONE 2008, 3, e3232. [Google Scholar] [CrossRef]
- Cobice, D.F.; Goodwin, R.J.A.; Andren, P.E.; Nilsson, A.; Mackay, C.L.; Andrew, R. Future technology insight: Mass spectrometry imaging as a tool in drug research and development. Br. J. Pharmacol. 2015, 172, 3266–3283. [Google Scholar] [CrossRef]
- Michno, W.; Wehrli, P.M.; Blennow, K.; Zetterberg, H.; Hanrieder, J. Molecular imaging mass spectrometry for probing protein dynamics in neurodegenerative disease pathology. J. Neurochem. 2018. [Google Scholar] [CrossRef] [PubMed]
- Chughtai, K.; Heeren, R.M.A. Mass Spectrometric Imaging for biomedical tissue analysis. Chem. Rev. 2010, 110, 3237–3277. [Google Scholar] [CrossRef] [PubMed]
- Jurchen, J.C.; Rubakhin, S.S.; Sweedler, J.V. MALDI-MS imaging of features smaller than the size of the laser beam. J. Am. Soc. Mass Spectrom. 2005, 16, 1654–1659. [Google Scholar] [CrossRef] [PubMed]
- Passarelli, M.K.; Wang, J.; Mohammadi, A.S.; Trouillon, R.; Gilmore, I.; Ewing, A.G. Development of an Organic Lateral Resolution Test Device for Imaging Mass Spectrometry. Anal. Chem. 2014, 86, 9473–9480. [Google Scholar] [CrossRef] [PubMed]
- Norris, J.L.; Caprioli, R.M. Analysis of Tissue Specimens by Matrix-Assisted Laser Desorption/Ionization Imaging Mass Spectrometry in Biological and Clinical Research. Chem. Rev. 2013, 113, 2309–2342. [Google Scholar] [CrossRef] [PubMed]
- Laiko, V.V.; Baldwin, M.A.; Burlingame, A.L. Atmospheric Pressure Matrix-Assisted Laser Desorption/Ionization Mass Spectrometry. Anal. Chem. 2000, 72, 652–657. [Google Scholar] [CrossRef]
- Schober, Y.; Guenther, S.; Spengler, B.; Römpp, A. Single Cell Matrix-Assisted Laser Desorption/Ionization Mass Spectrometry Imaging. Anal. Chem. 2012, 84, 6293–6297. [Google Scholar] [CrossRef]
- Nguyen, J.; Russell, S.C. Targeted proteomics approach to species-level identification of Bacillus thuringiensis spores by AP-MALDI-MS. J. Am. Soc. Mass Spectrom. 2010, 21, 993–1001. [Google Scholar] [CrossRef][Green Version]
- Baker, T.C.; Han, J.; Borchers, C.H. Recent advancements in matrix-assisted laser desorption/ionization mass spectrometry imaging. Curr. Opin. Biotechnol. 2017, 43, 62–69. [Google Scholar] [CrossRef]
- Fitzgerald, M.C.; Parr, G.R.; Smith, L.M. Basic matrices for the matrix-assisted laser desorption/ionization mass spectrometry of proteins and oligonucleotides. Anal. Chem. 1993, 65, 3204–3211. [Google Scholar] [CrossRef]
- Cheng, S.-W.; Chan, T.-W.D. Use of Ammonium Halides as Co-matrices for Matrix-assisted Laser Desorption/Ionization Studies of Oligonucleotides. Rapid Commun. Mass Spectrom. 1996, 10, 907–910. [Google Scholar] [CrossRef]
- Nordhoff, E.; Ingendoh, A.; Cramer, R.; Overberg, A.; Stahl, B.; Karas, M.; Hillenkamp, F.; Crain, P.F.; Chait, B. Matrix-assisted laser desorption/ionization mass spectrometry of nucleic acids with wavelengths in the ultraviolet and infrared. Rapid Commun. Mass Spectrom. 1992, 6, 771–776. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.-K.; Gross, M.L. Matrix-assisted laser desorption/ionization mass spectrometry methods for oligodeoxynucleotides: Improvements in matrix, detection limits, quantification, and sequencing. J. Am. Soc. Mass Spectrom. 2000, 11, 854–865. [Google Scholar] [CrossRef][Green Version]
- Lecchi, P.; Pannell, L.K. The detection of intact double-stranded DNA by MALDI. J. Am. Soc. Mass Spectrom. 1995, 6, 972–975. [Google Scholar] [CrossRef]
- Papac, D.I.; Wong, A.; Jones, A.J.S. Analysis of Acidic Oligosaccharides and Glycopeptides by Matrix-Assisted Laser Desorption/Ionization Time-of-Flight Mass Spectrometry. Anal. Chem. 1996, 68, 3215–3223. [Google Scholar] [CrossRef] [PubMed]
- Wu, K.J.; Shaler, T.A.; Becker, C.H. Time-of-Flight Mass Spectrometry Of Underivatized Single-Stranded DNA Oligomers by Matrix-Assisted Laser Desorption. Anal. Chem. 1994, 66, 1637–1645. [Google Scholar] [CrossRef]
- Shahgholi, M.; Garcia, B.A.; Chiu, N.H.L.; Heaney, P.J.; Tang, K. Sugar additives for MALDI matrices improve signal allowing the smallest nucleotide change (A:T) in a DNA sequence to be resolved. Nucleic Acids Res. 2001, 29, e91. [Google Scholar] [CrossRef]
- Beavis, R.C.; Chaudhary, T.; Chait, B.T. α-Cyano-4-hydroxycinnamic acid as a matrix for matrixassisted laser desorption mass spectromtry. J. Mass Spectrom. 1992, 27, 156–158. [Google Scholar] [CrossRef]
- Asara, J.M.; Allison, J. Enhanced detection of phosphopeptides in matrix-assisted laser desorption/ionization mass spectrometry using ammonium salts. J. Am. Soc. Mass Spectrom. 1999, 10, 35–44. [Google Scholar] [CrossRef]
- Gobom, J.; Schuerenberg, M.; Mueller, M.; Theiss, D.; Lehrach, H.; Nordhoff, E. Alpha-cyano-4-hydroxycinnamic acid affinity sample preparation. A protocol for MALDI-MS peptide analysis in proteomics. Anal. Chem. 2001, 73, 434–438. [Google Scholar] [CrossRef]
- Strupat, K.; Karas, M.; Hillenkamp, F. 2,5-Dihydroxybenzoic acid: A new matrix for laser desorption—Ionization mass spectrometry. Int. J. Mass Spectrom. Ion Process. 1991, 111, 89–102. [Google Scholar] [CrossRef]
- Zhu, L.; Parr, G.R.; Fitzgerald, M.C.; Nelson, C.M.; Smith, L.M. Oligodeoxynucleotide Fragmentation in MALDI/TOF Mass Spectrometry Using 355-nm Radiation. J. Am. Chem. Soc. 1995, 117, 6048–6056. [Google Scholar] [CrossRef]
- Macha, S.F.; Limbach, P.A.; Hanton, S.D.; Owens, K.G. Silver cluster interferences in matrix-assisted laser desorption/ionization (MALDI) mass spectrometry of nonpolar polymers. J. Am. Soc. Mass Spectrom. 2001, 12, 732–743. [Google Scholar] [CrossRef][Green Version]
- Vermillion-Salsbury, R.L.; Hercules, D.M.; Vermillion-Salsbury, R.L. 9-Aminoacridine as a matrix for negative mode matrix-assisted laser desorption/ionization. Rapid Commun. Mass Spectrom. 2002, 16, 1575–1581. [Google Scholar] [CrossRef]
- Eberlin, L.S.; Ferreira, C.R.; Dill, A.L.; Ifa, D.R.; Cooks, R.G. Desorption electrospray ionization mass spectrometry for lipid characterization and biological tissue imaging. Biochim. Biophys. Acta (BBA) 2011, 1811, 946–960. [Google Scholar] [CrossRef] [PubMed]
- Takats, Z.; Wiseman, J.M.; Gologan, B.; Cooks, R.G. Mass Spectrometry Sampling Under Ambient Conditions with Desorption Electrospray Ionization. Science 2004, 306, 471–473. [Google Scholar] [CrossRef] [PubMed]
- Eberlin, L.S.; Ferreira, C.R.; Dill, A.L.; Ifa, D.R.; Cheng, L.; Cooks, R.G. Non-Destructive, Histologically Compatible Tissue Imaging by Desorption Electrospray Ionization Mass Spectrometry. ChemBioChem 2011, 12, 2129–2132. [Google Scholar] [CrossRef] [PubMed]
- Dill, A.L.; Eberlin, L.S.; Costa, A.B.; Ifa, D.R.; Cooks, R.G. Data quality in tissue analysis using desorption electrospray ionization. Anal. Bioanal. Chem. 2011, 401, 1949–1961. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Hou, Y.; Hou, Z.; Xiong, W.; Huang, G. Mass Spectrometry Imaging of Brain Cholesterol and Metabolites with Trifluoroacetic Acid-Enhanced Desorption Electrospray Ionization. Anal. Chem. 2019, 91, 2719–2726. [Google Scholar] [CrossRef] [PubMed]
- Wiseman, J.M.; Ifa, D.R.; Song, Q.; Cooks, R.G. Tissue Imaging at Atmospheric Pressure Using Desorption Electrospray Ionization (DESI) Mass Spectrometry. Angew. Chem. Int. Ed. 2006, 45, 7188–7192. [Google Scholar] [CrossRef] [PubMed]
- Todd, P.J.; Schaaff, T.G.; Chaurand, P.; Caprioli, R.M. Organic ion imaging of biological tissue with secondary ion mass spectrometry and matrix-assisted laser desorption/ionization. J. Mass Spectrom. 2001, 36, 355–369. [Google Scholar] [CrossRef] [PubMed]
- Falick, A.M.; Maltby, D.A. Derivatization of hydrophilic peptides for liquid secondary ion mass spectrometry at the picomole level. Anal. Biochem. 1989, 182, 165–169. [Google Scholar] [CrossRef]
- Weibel, D.; Wong, S.; Lockyer, N.; Blenkinsopp, P.; Hill, R.; Vickerman, J.C. A C60Primary Ion Beam System for Time of Flight Secondary Ion Mass Spectrometry: Its Development and Secondary Ion Yield Characteristics. Anal. Chem. 2003, 75, 1754–1764. [Google Scholar] [CrossRef] [PubMed]
- Hebert, L.E.; Weuve, J.; Scherr, P.A.; Evans, D.A. Alzheimer disease in the United States (2010–2050) estimated using the 2010 census. Neurology 2013, 80, 1778–1783. [Google Scholar] [CrossRef]
- Wimo, A.; Jönsson, L.; Bond, J.; Prince, M.; Winblad, B. The worldwide economic impact of dementia 2010. Alzheimer’s Dement. 2013, 9, 1–11. [Google Scholar] [CrossRef]
- Marras, C.; Beck, J.C.; Bower, J.H.; Roberts, E.; Ritz, B.; Ross, G.W.; Abbott, R.D.; Savica, R.; Van Den Eeden, S.K.; Willis, A.W.; et al. Prevalence of Parkinson’s disease across North America. NPJ Parkinsons Dis. 2018, 4, 21. [Google Scholar] [CrossRef]
- Wang, X.; Xu, Z.; Xu, L. SP100B Expression Indexed Hemorrhage in Mouse Models of Cerebral Hemorrhage. Indian J. Clin. Biochem. 2018, 33, 361–364. [Google Scholar] [CrossRef]
- Alzheimer’s Association. 2018 Alzheimer’s disease facts and figures. Alzheimer’s Dement. 2018, 14, 367–429. [Google Scholar] [CrossRef]
- Serrano-Pozo, A.; Frosch, M.P.; Masliah, E.; Hyman, B.T. Neuropathological Alterations in Alzheimer Disease. Cold Spring Harb. Perspect. Med. 2011, 1, a006189. [Google Scholar] [CrossRef]
- Rodriguez, A.; Ehlenberger, D.B.; Dickstein, D.L.; Hof, P.R.; Wearne, S.L. Automated Three-Dimensional Detection and Shape Classification of Dendritic Spines from Fluorescence Microscopy Images. PLoS ONE 2008, 3, e1997. [Google Scholar] [CrossRef]
- Román, E.G.D.S.; Manuel, I.; Giralt, M.; Ferrer, I.; Rodríguez-Puertas, R. Imaging mass spectrometry (IMS) of cortical lipids from preclinical to severe stages of Alzheimer’s disease. Biochim. Biophys. Acta (BBA) 2017, 1859, 1604–1614. [Google Scholar] [CrossRef] [PubMed]
- Chan, R.B.; Oliveira, T.G.; Cortes, E.P.; Honig, L.S.; Duff, K.E.; Small, S.A.; Wenk, M.R.; Shui, G.; Di Paolo, G. Comparative lipidomic analysis of mouse and human brain with Alzheimer disease. J. Biol. Chem. 2012, 287, 2678–2688. [Google Scholar] [CrossRef] [PubMed]
- Donovan, M.H.; Yazdani, U.; Norris, R.D.; Games, D.; German, D.C.; Eisch, A.J. Decreased adult hippocampal neurogenesis in the PDAPP mouse model of Alzheimer’s disease. J. Comp. Neurol. 2006, 495, 70–83. [Google Scholar] [CrossRef] [PubMed]
- Semerci, F.; Maletic-Savatic, M. Transgenic mouse models for studying adult neurogenesis. Front. Boil. 2016, 11, 151–167. [Google Scholar] [CrossRef] [PubMed]
- Beccari, S.; Valero, J.; Maletic-Savatic, M.; Sierra, A. A simulation model of neuroprogenitor proliferation dynamics predicts age-related loss of hippocampal neurogenesis but not astrogenesis. Sci. Rep. 2017, 7, 16528. [Google Scholar] [CrossRef] [PubMed]
- Moreno-Jiménez, E.P.; Flor-García, M.; Terreros-Roncal, J.; Rábano, A.; Cafini, F.; Pallas-Bazarra, N.; Ávila, J.; Llorens-Martín, M. Adult hippocampal neurogenesis is abundant in neurologically healthy subjects and drops sharply in patients with Alzheimer’s disease. Nat. Med. 2019, 25, 554–560. [Google Scholar] [CrossRef] [PubMed]
- Crews, L.; Masliah, E. Molecular mechanisms of neurodegeneration in Alzheimer’s disease. Hum. Mol. Genet. 2010, 19, R12–R20. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Sierra, A.; Deudero, J.J.; Semerci, F.; Laitman, A.; Kimmel, M.; Maletic-Savatic, M. Multitype Bellman-Harris branching model provides biological predictors of early stages of adult hippocampal neurogenesis. BMC Syst. Boil. 2017, 11, 90. [Google Scholar] [CrossRef]
- González-Domínguez, R.; García-Barrera, T.; Vitorica, J.; Gómez-Ariza, J.L. Region-specific metabolic alterations in the brain of the APP/PS1 transgenic mice of Alzheimer’s disease. Biochim. Biophys. Acta (BBA) 2014, 1842, 2395–2402. [Google Scholar] [CrossRef]
- González-Domínguez, R.; García-Barrera, T.; Vitorica, J.; Gómez-Ariza, J.L. Metabolomic screening of regional brain alterations in the APP/PS1 transgenic model of Alzheimer’s disease by direct infusion mass spectrometry. J. Pharm. Biomed. Anal. 2015, 102, 425–435. [Google Scholar] [CrossRef]
- Xu, J.; Begley, P.; Church, S.J.; Patassini, S.; Hollywood, K.A.; Jüllig, M.; Curtis, M.A.; Waldvogel, H.J.; Faull, R.L.; Unwin, R.D.; et al. Graded perturbations of metabolism in multiple regions of human brain in Alzheimer’s disease: Snapshot of a pervasive metabolic disorder. Biochim. Biophys. Acta (BBA) 2016, 1862, 1084–1092. [Google Scholar] [CrossRef] [PubMed]
- Kaya, I.; Zetterberg, H.; Blennow, K.; Hanrieder, J. Shedding Light on the Molecular Pathology of Amyloid Plaques in Transgenic Alzheimer’s Disease Mice Using Multimodal MALDI Imaging Mass Spectrometry. ACS Chem. Neurosci. 2018, 9, 1802–1817. [Google Scholar] [CrossRef] [PubMed]
- Mendis, L.H.S.; Grey, A.C.; Faull, R.L.M.; Curtis, M.A. Hippocampal lipid differences in Alzheimer’s disease: A human brain study using matrix-assisted laser desorption/ionization-imaging mass spectrometry. Brain Behav. 2016, 6, e00517. [Google Scholar] [CrossRef] [PubMed]
- Caughlin, S.; Maheshwari, S.; Agca, Y.; Agca, C.; Harris, A.J.; Jurcic, K.; Yeung, K.K.-C.; Cechetto, D.F.; Whitehead, S.N. Membrane-lipid homeostasis in a prodromal rat model of Alzheimer’s disease: Characteristic profiles in ganglioside distributions during aging detected using MALDI imaging mass spectrometry. Biochim. Biophys. Acta (BBA) 2018, 1862, 1327–1338. [Google Scholar] [CrossRef] [PubMed]
- Cruz-Alonso, M.; Fernandez, B.; Navarro, A.; Junceda, S.; Astudillo, A.; Pereiro, R. Laser ablation ICP-MS for simultaneous quantitative imaging of iron and ferroportin in hippocampus of human brain tissues with Alzheimer’s disease. Talanta 2019, 197, 413–421. [Google Scholar] [CrossRef] [PubMed]
- Hare, D.J.; Raven, E.P.; Roberts, B.R.; Bogeski, M.; Portbury, S.D.; McLean, C.A.; Masters, C.L.; Connor, J.R.; Bush, A.I.; Crouch, P.J.; et al. Laser ablation-inductively coupled plasma-mass spectrometry imaging of white and gray matter iron distribution in Alzheimer’s disease frontal cortex. NeuroImage 2016, 137, 124–131. [Google Scholar] [CrossRef] [PubMed]
- Markesbery, W.R. Oxidative Stress Hypothesis in Alzheimer’s Disease. Free. Radic. Boil. Med. 1997, 23, 134–147. [Google Scholar] [CrossRef]
- Dauer, W.; Przedborski, S. Parkinson’s disease: Mechanisms and models. Neuron 2003, 39, 889–909. [Google Scholar] [CrossRef]
- Stauber, J.; Lemaire, R.; Franck, J.; Bonnel, D.; Croix, D.; Day, R.; Wisztorski, M.; Fournier, I.; Salzet, M. MALDI Imaging of Formalin-Fixed Paraffin-Embedded Tissues: Application to Model Animals of Parkinson Disease for Biomarker Hunting. J. Proteome Res. 2008, 7, 969–978. [Google Scholar] [CrossRef]
- Rose, S.; Chen, F.; Chalk, J.; Zelaya, F.; Strugnell, W.; Benson, M.; Semple, J.; Doddrell, D.; Rose, S.E.; Stuerenburg, H.J.; et al. Loss of connectivity in Alzheimer’s disease: An evaluation of white matter tract integrity with colour coded MR diffusion tensor imaging. J. Neurol. Neurosurg. Psychiatry 2000, 69, 528–530. [Google Scholar] [CrossRef]
- Gu, Y.; Hamajima, N.; Ihara, Y. Neurofibrillary Tangle-Associated Collapsin Response Mediator Protein-2 (CRMP-2) Is Highly Phosphorylated on Thr-509, Ser-518, and Ser-522. Biochemistry 2000, 39, 4267–4275. [Google Scholar] [CrossRef] [PubMed]
- Sköld, K.; Svensson, M.; Nilsson, A.; Zhang, X.; Nydahl, K.; Caprioli, R.M.; Svenningsson, P.; Andrén, P.E. Decreased Striatal Levels of PEP-19 Following MPTP Lesion in the Mouse. J. Proteome Res. 2006, 5, 262–269. [Google Scholar] [CrossRef] [PubMed]
- Ljungdahl, A.; Hanrieder, J.; Fälth, M.; Bergquist, J.; Andersson, M. Imaging Mass Spectrometry Reveals Elevated Nigral Levels of Dynorphin Neuropeptides in L-DOPA-Induced Dyskinesia in Rat Model of Parkinson’s Disease. PLoS ONE 2011, 6, e25653. [Google Scholar] [CrossRef] [PubMed]
- Schapira, A.H.; Agid, Y.; Barone, P.; Jenner, P.; Lemke, M.R.; Poewe, W.; Rascol, O.; Reichmann, H.; Tolosa, E. Perspectives on recent advances in the understanding and treatment of Parkinson’s disease. Eur. J. Neurol. 2009, 16, 1090–1099. [Google Scholar] [CrossRef] [PubMed]
- Shariatgorji, M.; Nilsson, A.; Goodwin, R.J.; Källback, P.; Schintu, N.; Zhang, X.; Crossman, A.R.; Bezard, E.; Svenningsson, P.; Andren, P.E. Direct Targeted Quantitative Molecular Imaging of Neurotransmitters in Brain Tissue Sections. Neuron 2014, 84, 697–707. [Google Scholar] [CrossRef] [PubMed]
- Hunter, M.; DeMarais, N.J.; Faull, R.L.M.; Grey, A.C.; Curtis, M.A. Subventricular zone lipidomic architecture loss in Huntington’s disease. J. Neurochem. 2018, 146, 613–630. [Google Scholar] [CrossRef] [PubMed]
- Arribat, Y.; Talmat-Amar, Y.; Paucard, A.; Lesport, P.; Bonneaud, N.; Bauer, C.; Bec, N.; Parmentier, M.-L.; Benigno, L.; Larroque, C.; et al. Systemic delivery of P42 peptide: A new weapon to fight Huntington’s disease. Acta Neuropathol. Commun. 2014, 2, 905. [Google Scholar] [CrossRef] [PubMed]
- Maccarrone, G.; Nischwitz, S.; Deininger, S.-O.; Hornung, J.; König, F.B.; Stadelmann, C.; Turck, C.W.; Weber, F. MALDI imaging mass spectrometry analysis—A new approach for protein mapping in multiple sclerosis brain lesions. J. Chromatogr. B 2017, 1047, 131–140. [Google Scholar] [CrossRef]
- Ekegren, T.; Hanrieder, J.; Andersson, M.; Bergquist, J. MALDI imaging of post-mortem human spinal cord in amyotrophic lateral sclerosis. J. Neurochem. 2013, 124, 695–707. [Google Scholar]
- Cepeda, C.; Wu, N.; Andre, V.M.; Cummings, D.M.; Levine, M.S. The corticostriatal pathway in Huntington’s disease. Prog. Neurobiol. 2007, 81, 253–271. [Google Scholar] [CrossRef]
- Zuccato, C.; Valenza, M.; Cattaneo, E. Molecular Mechanisms and Potential Therapeutical Targets in Huntington’s Disease. Physiol. Rev. 2010, 90, 905–981. [Google Scholar] [CrossRef] [PubMed]
- Walker, T.M.; Lalor, M.K.; Broda, A.; Ortega, L.S.; Morgan, M.; Parker, L.; Churchill, S.; Bennett, K.; Golubchik, T.; Giess, A.P.; et al. Assessment of Mycobacterium tuberculosis transmission in Oxfordshire, UK, 2007–12, with whole pathogen genome sequences: An observational study. Lancet Respir. Med. 2014, 2, 285–292. [Google Scholar] [CrossRef]
- Patassini, S.; Begley, P.; Reid, S.J.; Xu, J.; Church, S.J.; Curtis, M.; Dragunow, M.; Waldvogel, H.J.; Unwin, R.D.; Snell, R.G.; et al. Identification of elevated urea as a severe, ubiquitous metabolic defect in the brain of patients with Huntington’s disease. Biochem. Biophys. Res. Commun. 2015, 468, 161–166. [Google Scholar] [CrossRef] [PubMed]
- Patassini, S.; Begley, P.; Xu, J.; Church, S.J.; Reid, S.J.; Kim, E.H.; Curtis, M.A.; Dragunow, M.; Waldvogel, H.J.; Snell, R.G.; et al. Metabolite mapping reveals severe widespread perturbation of multiple metabolic processes in Huntington’s disease human brain. Biochim. Biophys. Acta (BBA) 2016, 1862, 1650–1662. [Google Scholar] [CrossRef] [PubMed]
- Graham, S.F.; Pan, X.; Yilmaz, A.; Macias, S.; Robinson, A.; Mann, D.; Green, B.D. Targeted biochemical profiling of brain from Huntington’s disease patients reveals novel metabolic pathways of interest. Biochim. Biophys. Acta (BBA) 2018, 1864, 2430–2437. [Google Scholar] [CrossRef] [PubMed]
- International Multiple Sclerosis Genetics Consortium. Refining genetic associations in multiple sclerosis. Lancet Neurol. 2008, 7, 567–569. [Google Scholar] [CrossRef]
- Goldenberg, M.M. Multiple sclerosis review. Pharm. Ther. 2012, 37, 175–184. [Google Scholar]
- Wheeler, D.; Bandaru, V.V.R.; Calabresi, P.A.; Nath, A.; Haughey, N.J. A defect of sphingolipid metabolism modifies the properties of normal appearing white matter in multiple sclerosis. Brain 2008, 131, 3092–3102. [Google Scholar] [CrossRef]
- Mangalam, A.K.; Poisson, L.M.; Nemutlu, E.; Datta, I.; Denic, A.; Dzeja, P.; Rodriguez, M.; Rattan, R.; Giri, S. Profile of Circulatory Metabolites in a Relapsing-remitting Animal Model of Multiple Sclerosis using Global Metabolomics. J. Clin. Cell. Immunol. 2013, 4, 1–12. [Google Scholar]
- Rowland, L.P.; Shneider, N.A. Amyotrophic Lateral Sclerosis. N. Engl. J. Med. 2001, 344, 1688–1700. [Google Scholar] [CrossRef]
- Clement, A.M. Wild-Type Nonneuronal Cells Extend Survival of SOD1 Mutant Motor Neurons in ALS Mice. Science 2003, 302, 113–117. [Google Scholar] [CrossRef] [PubMed]
- Cudkowicz, M.E.; van den Berg, L.H.; Shefner, J.M.; Mitsumoto, H.; Mora, J.S.; Ludolph, A.; Hardiman, O.; Bozik, M.E.; Ingersoll, E.W.; Archibald, D.; et al. Dexpramipexole versus placebo for patients with amyotrophic lateral sclerosis (EMPOWER): A randomised, double-blind, phase 3 trial. Lancet Neurol. 2013, 12, 1059–1067. [Google Scholar] [CrossRef]



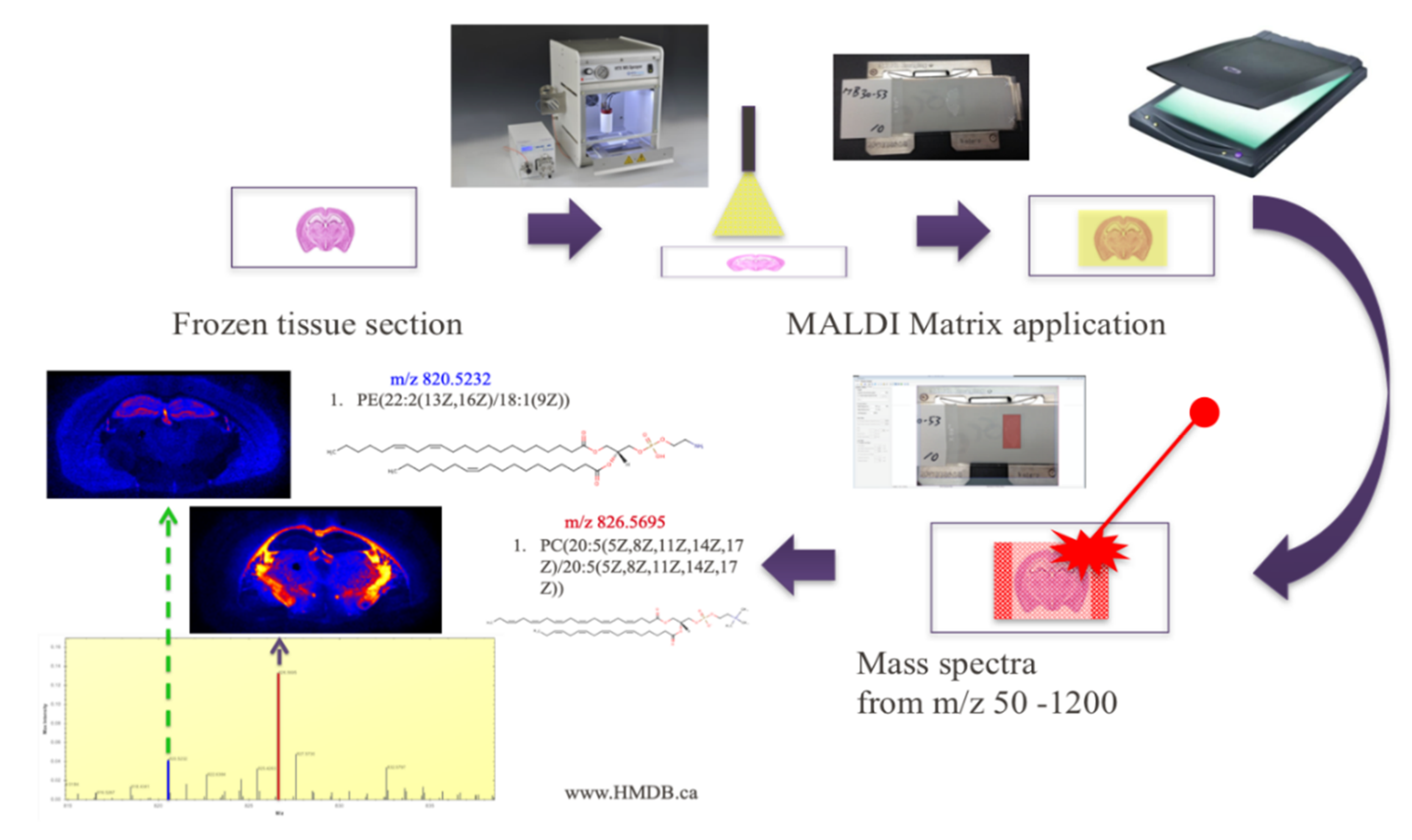{kind=link}
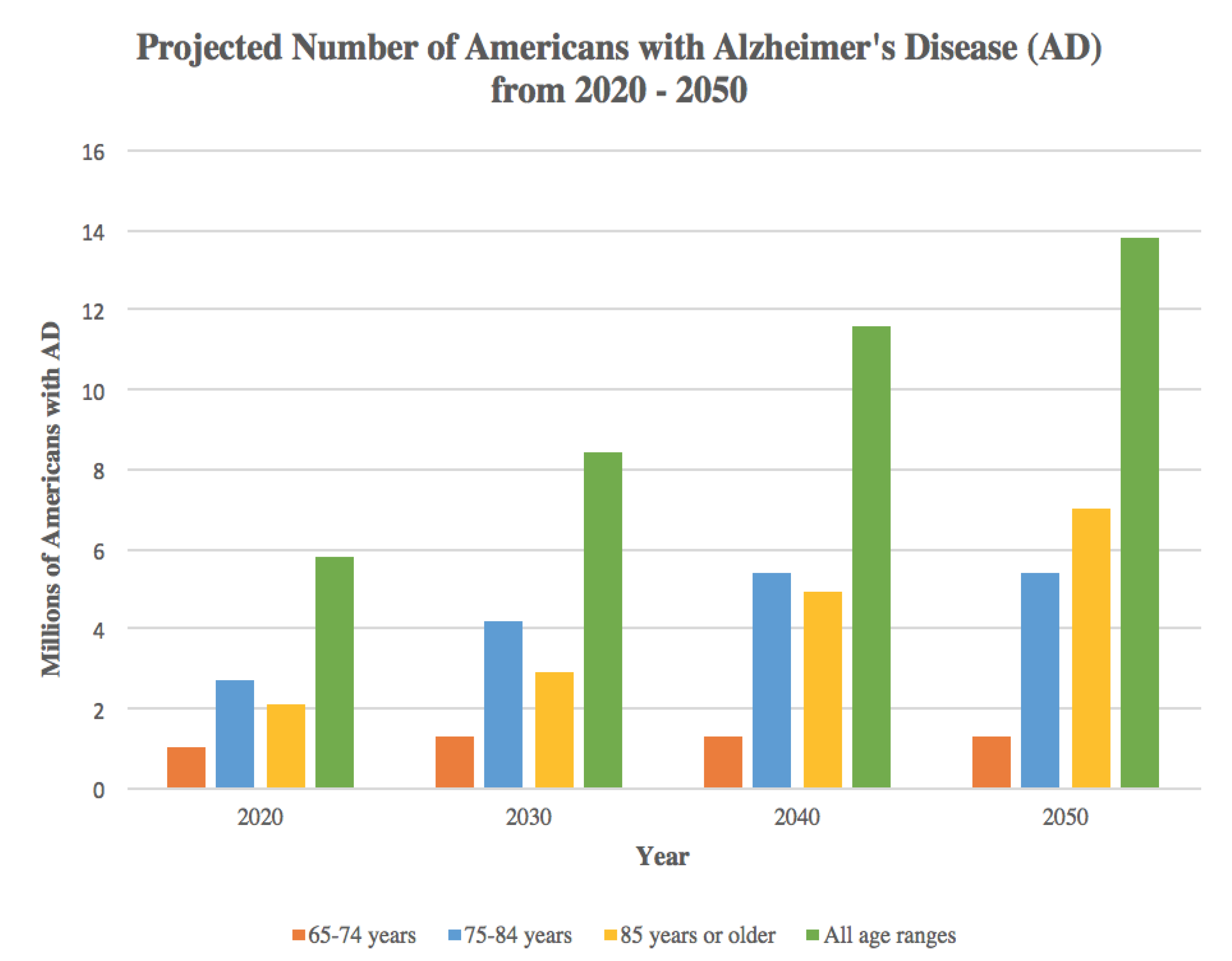{kind=link}
| Platform | Mechanism | Advantages | Disadvantages |
|---|---|---|---|
| Matrix-assisted laser desorption/ionization (MALDI) | UVlaser used for the desorption and ionization of analytes after application of an UV-absorbing matrix into a gaseous state |
|
|
| Desorption electrospray ionization (DESI) | Electrically-charged solvent drops sprayed onto sample surface to eject analyte molecules into a gaseous state |
|
|
| Secondary ion mass spectrometry (SIMS) | High-energy primary ion beam (i.e., gallium and indium ions) facilitates the desorption and ionization of analytes in the form of secondary ions in the gaseous state |
|
|
| Matrix | Application | References |
|---|---|---|
| 2-amino-5-nitropyridine (ANP) | Oligonucleotides < 20 Bases, MALDI (−) | [30,31] |
| 80% anthranilic acid + 20% nicotinic acid (80/20 AA/NA) | Oligonucleotides < 20 Bases, MALDI (+ & −) | [32,33] |
| 6-aza-2-thiothymine (6-ATT) | Oligonucleotides and carbohydrates MALDI (−) | [34,35] |
| 3-hydroxypicolinic acid (3-HPA) | Oligonucleotides 1 kDa−30 kDa | [36,37] |
| α cyano 4 hydroxycinnamic acid (CHCA) | small molecules, peptides/proteins < 6 kDa | [38,39,40] |
| 2,5 dihydroxybenzoic acid (2,5 DHB) | small molecules, peptides/proteins < 6 kDa, polymers, carbohydrates | [39,41,42,43] |
| 9-aminoacridine (9-AA) | small molecules, lipids, MALDI (−) | [44] |
| Disease | Organism | Findings | Ref. |
|---|---|---|---|
| Alzheimer’s Disease (AD) | Humans | Relative abundance of lipid species differed between AD and control brains, predominantly in the CA1 and dentate gyrus regions of the hippocampus. | [73] |
| - | Humans | Sulfatide concentrations start to decrease during the early stages of AD (determined by Braak’s histochemical criteria) in the white and gray matter of the frontal cortex. | [61] |
| - | Humans | Sphingolipids (e.g., ceramides, sulfatides, and gangliosides) show selective localization to the β-amyloid aggregates within the cortex and hippocampus, with specific localization of gangliosides and ceramides to β-amyloid positive plaques. | [72] |
| - | Humans | The CA1 region of the hippocampus in the AD brain contains higher levels of iron compared to the control brain, but there is no significant difference in the levels of ferroportin transport protein between AD and control brains. | [75] |
| - | Mice | Lipid signatures of the AD mouse brain exhibit early shifts in lipid homeostasis: its white matter is composed of higher levels of simple gangliosides and lower levels of complex gangliosides, such as the GD1 d18:1 species. | [74] |
| Parkinson’s Disease (PD) | Mice | Collapsin response mediator protein 2 (CRMP-2) detected in the PD brain though usually only found in hippocampal and cerebellar cells. | [79] |
| - | Mice | Calmodulin-binding protein (PEP-19), normally localized to the striatum, was significantly downregulated in the stratium of the PD brain. | [82] |
| - | Mice | Distinct differences in both the levels and localization of various neurotransmitters and amino acid between PD and control brains established. | [85] |
| - | Mice | Dynorphin B and alpha-neoendrophin nigral levels are positively correlated with the severity of L-DOPA-induced-dyskinesia. | [83] |
| Huntington’s Disease (HD) | Mice | Within the HD myelin layer, sulfatide and triglyceride levels are decreased and sphingomyelin and ceramide-1-phosphate levels are increased in the lamina; within the HD ependymal layer, phosphatidylinositols levels are decreased. | [86] |
| - | Mice | Efficacy of P42, a 23 amino acid peptide sequence, as a novel therapy for HD was analyzed with IMS to confirm drug delivery, investigate pharmokinetic properties, and observe post-delivery molecular change. | [87] |
| Multiple Sclerosis (MS) | Humans | Thymosin beta-4 protein localized to active MS lesions that were either chronically demyelinated or only partially remyelinated. | [88] |
| Amyotrophic Lateral Sclerosis (ALS) | Humans | Truncated ubiquition form (Ubc-174) levels decreased significantly in ALS spinal cords compared to control, which paralleled normal histological distributes of metabolites in the gray matter. | [89] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, K.; Baluya, D.; Tosun, M.; Li, F.; Maletic-Savatic, M. Imaging Mass Spectrometry: A New Tool to Assess Molecular Underpinnings of Neurodegeneration. Metabolites 2019, 9, 135. https://doi.org/10.3390/metabo9070135
Chen K, Baluya D, Tosun M, Li F, Maletic-Savatic M. Imaging Mass Spectrometry: A New Tool to Assess Molecular Underpinnings of Neurodegeneration. Metabolites. 2019; 9(7):135. https://doi.org/10.3390/metabo9070135
Chicago/Turabian StyleChen, Kevin, Dodge Baluya, Mehmet Tosun, Feng Li, and Mirjana Maletic-Savatic. 2019. "Imaging Mass Spectrometry: A New Tool to Assess Molecular Underpinnings of Neurodegeneration" Metabolites 9, no. 7: 135. https://doi.org/10.3390/metabo9070135
APA StyleChen, K., Baluya, D., Tosun, M., Li, F., & Maletic-Savatic, M. (2019). Imaging Mass Spectrometry: A New Tool to Assess Molecular Underpinnings of Neurodegeneration. Metabolites, 9(7), 135. https://doi.org/10.3390/metabo9070135