Intracellular and Extracellular Metabolites from the Cyanobacterium Chlorogloeopsis fritschii, PCC 6912, During 48 Hours of UV-B Exposure



Abstract

{kind=link}
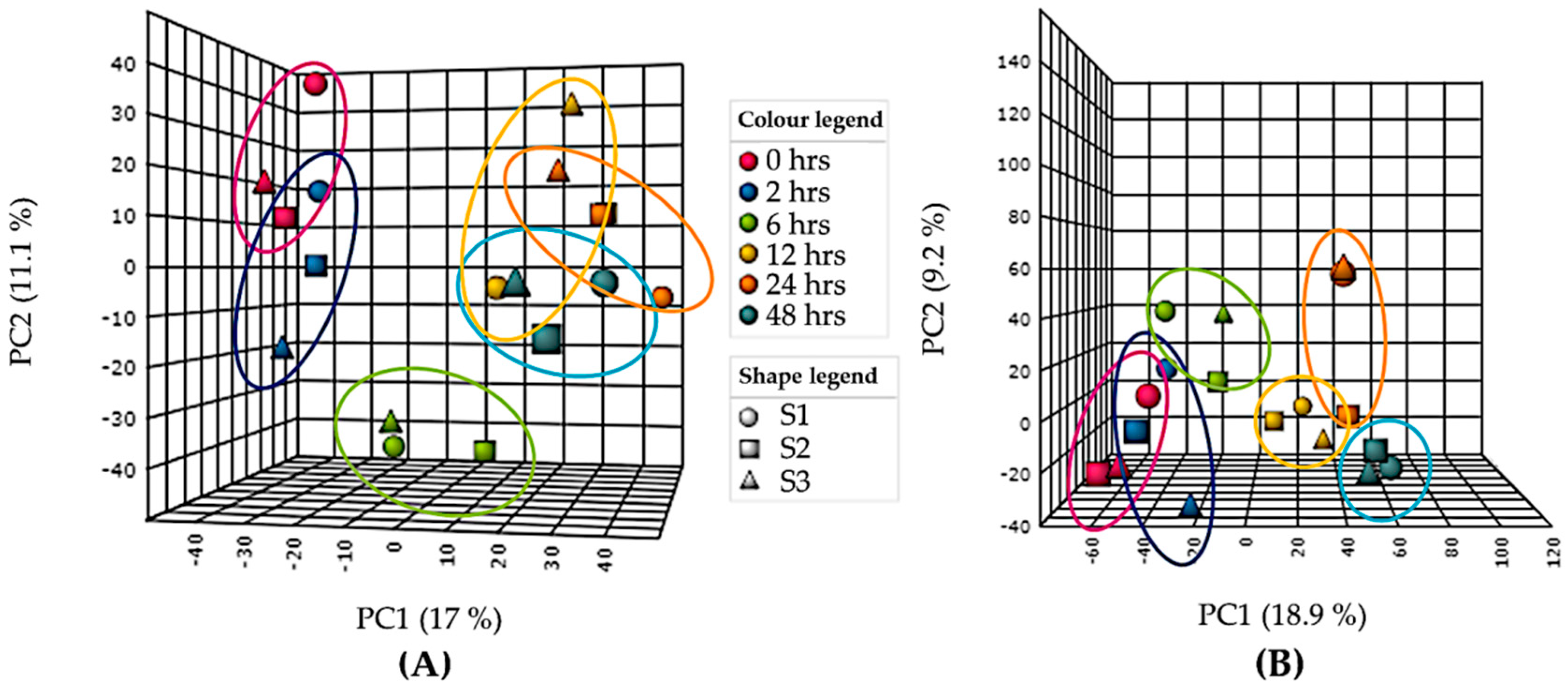{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
2.1. Intracellular and Extracellular Analysis of C. fritschii during UV-B Stress Response
2.1.1. Intracellular Metabolites
Carotenoid and MAA Analysis
2.1.2. Extracellular Metabolites
2.2. Intracellular Analysis of C. fritschii (PAR Only)
Intracellular Metabolites
3. Discussion
3.1 Intracellular Metabolite Changes and Pathway Analysis
3.2 Extracellular Metabolite Changes
4. Conclusions
5. Materials and Methods
5.1. Organism and Growth Conditions
5.2. Experimental Setup
5.2.1. Supplemented UV-B Experiment (PAR + UV-B)
5.2.2. PAR Only Experiment (PAR only)
5.3. Sample Harvest and Growth Analysis
5.4. GC–MS Analysis
5.4.1. Sample Preparation
5.4.2. Sample Derivatisation
5.4.3. GC–MS Analysis
5.4.4. GC–MS Data Processing
5.4.5. Identification
5.4.6. Statistical Analysis
5.5. MAA analysis
5.5.1. Sample Preparation
5.5.2. HPLC Analysis
5.6. Pigment Analysis
Sample Preparation
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Gupta, V.; Ratha, S.K.; Sood, A.; Chaudhary, V.; Prasanna, R. New Insights into the Biodiversity and Applications of Cyanobacteria (Blue-Green Algae)—Prospects and Challenges. Algal Res. 2013, 2, 79–97. [Google Scholar] [CrossRef]
- Schwarz, D.; Orf, I.; Kopka, J.; Hagemann, M. Recent Applications of Metabolomics Toward Cyanobacteria. Metabolites 2013, 3, 72–100. [Google Scholar] [CrossRef] [PubMed]
- Wijffels, R.H.; Kruse, O.; Hellingwerf, K.J. Potential of Industrial Biotechnology with Cyanobacteria and Eukaryotic Microalgae. Curr. Opin. Biotechnol. 2013, 24, 405–413. [Google Scholar] [CrossRef] [PubMed]
- Rastogi, R.P.; Sinha, R.P. Biotechnological and Industrial Significance of Cyanobacterial Secondary Metabolites. Biotechnol. Adv. 2009, 27, 521–539. [Google Scholar] [CrossRef] [PubMed]
- Rastogi, R.P.; Sinha, R.P.; Moh, S.H.; Lee, T.K.; Kottuparambil, S.; Kim, Y.J.; Rhee, J.S.; Choi, E.M.; Brown, M.T.; Häder, D.P.; et al. Ultraviolet Radiation and Cyanobacteria. J. Photochem. Photobiol. B Biol. 2014, 141, 154–169. [Google Scholar] [CrossRef]
- Rastogi, R.P. UV-Induced Oxidative Stress in Cyanobacteria: How Life Is Able to Survive? Biochem. Anal. Biochem. 2015, 4, 2–5. [Google Scholar] [CrossRef]
- Marangoni, R.; Paris, D.; Melck, D.; Fulgentini, L.; Colombetti, G.; Motta, A. In Vivo NMR Metabolic Profiling of Fabrea Salina Reveals Sequential Defense Mechanisms against Ultraviolet Radiation. Biophys. J. 2011, 100. [Google Scholar] [CrossRef]
- Sinha, R.P.; Häder, D.P. UV-Protectants in Cyanobacteria. Plant Sci. 2008, 174, 278–289. [Google Scholar] [CrossRef]
- Wada, N.; Sakamoto, T.; Matsugo, S. Multiple Roles of Photosynthetic and Sunscreen Pigments in Cyanobacteria Focusing on the Oxidative Stress. Metabolites 2013, 3, 463–483. [Google Scholar] [CrossRef]
- Rastogi, R.P.; Incharoensakdi, A. Characterization of UV-Screening Compounds, Mycosporine-like Amino Acids, and Scytonemin in the Cyanobacterium Lyngbya Sp. CU2555. FEMS Microbiol. Ecol. 2014, 87, 244–256. [Google Scholar] [CrossRef]
- Ehling-Schulz, M.; Bilger, W.; Scherer, S. UV-B-Induced Synthesis of Photoprotective Pigments and Extracellular Polysaccharides in the Terrestrial Cyanobacterium Nostoc Commune. J. Bacteriol. 1997, 179, 1940–1945. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.P.; Klisch, M.; Sinha, R.P.; Hader, D.-P. Effects of Abiotic Stressors on Synthesis of the Mycosporine-like Amino Acid Shinorine in the Cyanobacterium Anabaena Variabilis PCC 7937. Photochem. Photobiol. 2008, 84, 1500–1505. [Google Scholar] [CrossRef] [PubMed]
- Hartmann, A.; Albert, A.; Ganzera, M. Effects of Elevated Ultraviolet Radiation on Primary Metabolites in Selected Alpine Algae and Cyanobacteria. J. Photochem. Photobiol. B Biol. 2015, 149, 149–155. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Portwich, A.; Garcia-Pichel, F. A Novel Prokaryotic UVB Photoreceptor in the Cyanobacterium Chlorogloeopsis PCC 6912. Photochem. Photobiol. 2000, 71, 493–498. [Google Scholar] [CrossRef]
- Ehling-Schulz, M.; Schulz, S.; Wait, R.; Görg, A.; Scherer, S. The UV-B Stimulon of the Terrestrial Cyanobacterium Nostoc Commune Comprises Early Shock Proteins and Late Acclimation Proteins. Mol. Microbiol. 2002, 46, 827–843. [Google Scholar] [CrossRef] [PubMed]
- Shrivastava, A.K.; Chatterjee, A.; Yadav, S.; Singh, P.K.; Singh, S.; Rai, L.C. UV-B Stress Induced Metabolic Rearrangements Explored with Comparative Proteomics in Three Anabaena Species. J. Proteomics 2015, 127, 122–133. [Google Scholar] [CrossRef] [PubMed]
- Shen, S.G.; Jia, S.R.; Yan, R.R.; Wu, Y.K.; Wang, H.Y.; Lin, Y.H.; Zhao, D.X.; Tan, Z.L.; Lv, H.X.; Han, P.P. The Physiological Responses of Terrestrial Cyanobacterium: Nostoc Flagelliforme to Different Intensities of Ultraviolet-B Radiation. RSC Adv. 2018, 8, 21065–21074. [Google Scholar] [CrossRef]
- Wase, N.; Pham, T.K.; Ow, S.Y.; Wright, P.C. Quantitative Analysis of UV-A Shock and Short Term Stress Using ITRAQ, Pseudo Selective Reaction Monitoring (PSRM) and GC-MS Based Metabolite Analysis of the Cyanobacterium Nostoc Punctiforme ATCC 29133. J. Proteomics 2014, 109, 332–355. [Google Scholar] [CrossRef] [PubMed]
- Fiore, C.L.; Longnecker, K.; Kido Soule, M.C.; Kujawinski, E.B. Release of Ecologically Relevant Metabolites by the Cyanobacterium S Ynechococcus Elongatus CCMP 1631. Environ. Microbiol. 2015, 17, 3949–3963. [Google Scholar] [CrossRef]
- Stuart, R.K.; Mayali, X.; Lee, J.Z.; Craig Everroad, R.; Hwang, M.; Bebout, B.M.; Weber, P.K.; Pett-Ridge, J.; Thelen, M.P. Cyanobacterial Reuse of Extracellular Organic Carbon in Microbial Mats. ISME J. 2016, 10, 1240–1251. [Google Scholar] [CrossRef] [PubMed]
- Kujawinski, E.B. The Impact of Microbial Metabolism on Marine Dissolved Organic Matter. Ann. Rev. Mar. Sci. 2011, 3, 567–599. [Google Scholar] [CrossRef]
- Yadav, S.; Sinha, R.P.; Tyagi, M.B.; Kumar, A. Cyanobacterial Secondary Metabolites. Int. J. Pharma Bio Sci. 2011, 2, 144–167. [Google Scholar]
- Mota, R.; Guimarães, R.; Büttel, Z.; Rossi, F.; Colica, G.; Silva, C.J.; Santos, C.; Gales, L.; Zille, A.; De Philippis, R.; et al. Production and Characterization of Extracellular Carbohydrate Polymer from Cyanothece Sp. CCY 0110. Carbohydr. Polym. 2013, 92, 1408–1415. [Google Scholar] [CrossRef] [PubMed]
- Sue, T.; Obolonkin, V.; Griffiths, H.; Villas-Bôas, S.G. An Exometabolomics Approach to Monitoring Microbial Contamination in Microalgal Fermentation Processes by Using Metabolic Footprint Analysis. Appl. Environ. Microbiol. 2011, 77, 7605–7610. [Google Scholar] [CrossRef] [PubMed]
- Pinu, F.; Villas-Boas, S. Extracellular Microbial Metabolomics: The State of the Art. Metabolites 2017, 7, 43. [Google Scholar] [CrossRef] [PubMed]
- Granucci, N.; Pinu, F.R.; Han, T.-L.; Villas-Boas, S.G. Can We Predict the Intracellular Metabolic State of a Cell Based on Extracellular Metabolite Data? Mol. Biosyst. 2015, 11, 3297–3304. [Google Scholar] [CrossRef] [PubMed]
- Balasundaram, B.; Skill, S.C.; Llewellyn, C. a. A Low Energy Process for the Recovery of Bioproducts from Cyanobacteria Using a Ball Mill. Biochem. Eng. J. 2012, 69, 48–56. [Google Scholar] [CrossRef]
- Evans, E.H.; Foulds, I.; Carr, N.G. Environmental Conditions and Morphological Variation in the Blue-Green Alga Chlorogloea Fritschii. J. Gen. Microbiol. 1976, 92, 147–155. [Google Scholar] [CrossRef]
- Portwich, A.; Garcia-Pichel, F. Ultraviolet and Osmotic Stresses Induce and Regulate the Synthesis of Mycosporines in the Cyanobacterium Chlorogloeopsis PCC 6912. Arch. Microbiol. 1999, 172, 187–192. [Google Scholar] [CrossRef] [PubMed]
- Airs, R.L.; Temperton, B.; Sambles, C.; Farnham, G.; Skill, S.C.; Llewellyn, C.A. Chlorophyll f and Chlorophyll d Are Produced in the Cyanobacterium Chlorogloeopsis Fritschii When Cultured under Natural Light and Near-Infrared Radiation. FEBS Lett. 2014, 588, 3770–3777. [Google Scholar] [CrossRef] [PubMed]
- Stanier, R.Y.; Deruelles, J.; Rippka, R.; Herdman, M.; Waterbury, J.B. Generic Assignments, Strain Histories and Properties of Pure Cultures of Cyanobacteria. Microbiology 1979, 111, 1–61. [Google Scholar]
- Singh, S.P.; Häder, D.P.; Sinha, R.P. Cyanobacteria and Ultraviolet Radiation (UVR) Stress: Mitigation Strategies. Ageing Res. Rev. 2010, 9, 79–90. [Google Scholar] [CrossRef]
- Latifi, A.; Ruiz, M.; Zhang, C.C. Oxidative Stress in Cyanobacteria. FEMS Microbiol. Rev. 2009, 33, 258–278. [Google Scholar] [CrossRef]
- Kumar, A.; Sinha, R.P.; Häder, D.-P. Effect of UV-B on Enzymes of Nitrogen Metabolism in the Cyanobacterium Nostoc Calcicola. J. Plant Physiol. 1996, 148, 86–91. [Google Scholar] [CrossRef]
- Kultschar, B.; Llewellyn, C. Secondary Metabolites in Cyanobacteria. In Secondary Metabolites—Sources and Applications; InTech: London, UK, 2018; Volume 2, p. 64. [Google Scholar]
- Cameron, J.C.; Pakrasi, H.B. Essential Role of Glutathione in Acclimation to Environmental and Redox Perturbations in the Cyanobacterium Synechocystis Sp. PCC 6803. Plant Physiol. 2010, 154, 1672–1685. [Google Scholar] [CrossRef] [PubMed]
- Liang, X.; Zhang, L.; Natarajan, S.K.; Becker, D.F. Proline Mechanisms of Stress Survival. Antioxid. Redox Signal. 2013, 19, 998–1011. [Google Scholar] [CrossRef] [PubMed]
- Verbruggen, N.; Hermans, C. Proline Accumulation in Plants: A Review. Amino Acids 2008, 35, 753–759. [Google Scholar] [CrossRef] [PubMed]
- Chris, A.; Zeeshan, M.; Abraham, G.; Prasad, S.M. Proline Accumulation in Cylindrospermum sp. Environ. Exp. Bot. 2006, 57, 154–159. [Google Scholar] [CrossRef]
- Kusano, M.; Tohge, T.; Fukushima, A.; Kobayashi, M.; Hayashi, N.; Otsuki, H.; Kondou, Y.; Goto, H.; Kawashima, M.; Matsuda, F.; et al. Metabolomics Reveals Comprehensive Reprogramming Involving Two Independent Metabolic Responses of Arabidopsis to UV-B Light. Plant J. 2011, 67, 354–369. [Google Scholar] [CrossRef] [PubMed]
- Pinu, F.R.; Granucci, N.; Daniell, J.; Han, T.-L.; Carneiro, S.; Rocha, I.; Nielsen, J.; Villas-Boas, S.G. Metabolite Secretion in Microorganisms: The Theory of Metabolic Overflow Put to the Test. Metabolomics 2018, 14, 43. [Google Scholar] [CrossRef]
- Shastri, A.A.; Morgan, J.A. A Transient Isotopic Labeling Methodology for 13C Metabolic Flux Analysis of Photoautotrophic Microorganisms. Phytochemistry 2007, 68, 2302–2312. [Google Scholar] [CrossRef] [PubMed]
- Styczynski, M.P.; Moxley, J.F.; Tong, L.V.; Walther, J.L.; Jensen, K.L.; Stephanopoulos, G.N. Systematic Identification of Conserved Metabolites in GC/MS Data for Metabolomics and Biomarker Discovery. Anal. Chem. 2007, 79, 966–973. [Google Scholar] [CrossRef] [PubMed]
- Kopka, J.; Schauer, N.; Krueger, S.; Birkemeyer, C.; Usadel, B.; Bergmüller, E.; Dörmann, P.; Weckwerth, W.; Gibon, Y.; Stitt, M.; et al. GMD@CSB.DB: The Golm Metabolome Database. Bioinformatics 2005, 21, 1635–1638. [Google Scholar] [CrossRef] [PubMed]
- Chong, J.; Soufan, O.; Li, C.; Caraus, I.; Li, S.; Bourque, G.; Wishart, D.S.; Xia, J. MetaboAnalyst 4.0: Towards More Transparent and Integrative Metabolomics Analysis. Nucleic Acids Res. 2018, 46, W486–W494. [Google Scholar] [CrossRef]
- Xia, J.; Sinelnikov, I.V.; Wishart, D.S. MetATT: A Web-Based Metabolomics Tool for Analyzing Time-Series and Two-Factor Datasets. Bioinformatics 2011, 27, 2455–2456. [Google Scholar] [CrossRef]
- Fiehn, O. Metabolomics by Gas Chromatography-Mass Spectrometry: Combined Targeted and Untargeted Profiling. In Current Protocols in Molecular Biology; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2016; Volume 131 A, pp. 30.4.1–30.4.32. [Google Scholar]
- Sumner, L.W.; Amberg, A.; Barrett, D.; Beale, M.H.; Beger, R.; Daykin, C.A.; Fan, T.W.M.; Fiehn, O.; Goodacre, R.; Griffin, J.L.; et al. Proposed Minimum Reporting Standards for Chemical Analysis: Chemical Analysis Working Group (CAWG) Metabolomics Standards Initiative (MSI). Metabolomics 2007, 3, 211–221. [Google Scholar] [CrossRef]
- Rastogi, R.P.; Madamwar, D.; Incharoensakdi, A. Sun-Screening Bioactive Compounds Mycosporine-like Amino Acids in Naturally Occurring Cyanobacterial Biofilms: Role in Photoprotection. J. Appl. Microbiol. 2015, 119, 753–762. [Google Scholar] [CrossRef] [PubMed]
- Ritchie, R.J. Consistent Sets of Spectrophotometric Chlorophyll Equations for Acetone, Methanol and Ethanol Solvents. Photosynth. Res. 2006, 89, 27–41. [Google Scholar] [CrossRef] [PubMed]
- Henriques, M.; Silva, A.; Rocha, J. Extraction and Quantification of Pigments from a Marine Microalga: A Simple and Reproducible Method. Commun. Curr. Res. Educ. Top. Trends Appl. Microbiol. 2007, 2, 586–593. [Google Scholar]






© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kultschar, B.; Dudley, E.; Wilson, S.; Llewellyn, C.A. Intracellular and Extracellular Metabolites from the Cyanobacterium Chlorogloeopsis fritschii, PCC 6912, During 48 Hours of UV-B Exposure. Metabolites 2019, 9, 74. https://doi.org/10.3390/metabo9040074
Kultschar B, Dudley E, Wilson S, Llewellyn CA. Intracellular and Extracellular Metabolites from the Cyanobacterium Chlorogloeopsis fritschii, PCC 6912, During 48 Hours of UV-B Exposure. Metabolites. 2019; 9(4):74. https://doi.org/10.3390/metabo9040074
Chicago/Turabian StyleKultschar, Bethan, Ed Dudley, Steve Wilson, and Carole A. Llewellyn. 2019. "Intracellular and Extracellular Metabolites from the Cyanobacterium Chlorogloeopsis fritschii, PCC 6912, During 48 Hours of UV-B Exposure" Metabolites 9, no. 4: 74. https://doi.org/10.3390/metabo9040074
APA StyleKultschar, B., Dudley, E., Wilson, S., & Llewellyn, C. A. (2019). Intracellular and Extracellular Metabolites from the Cyanobacterium Chlorogloeopsis fritschii, PCC 6912, During 48 Hours of UV-B Exposure. Metabolites, 9(4), 74. https://doi.org/10.3390/metabo9040074