The Metabolomic Signatures of Weight Change



{kind=link}
Abstract
:1. Introduction
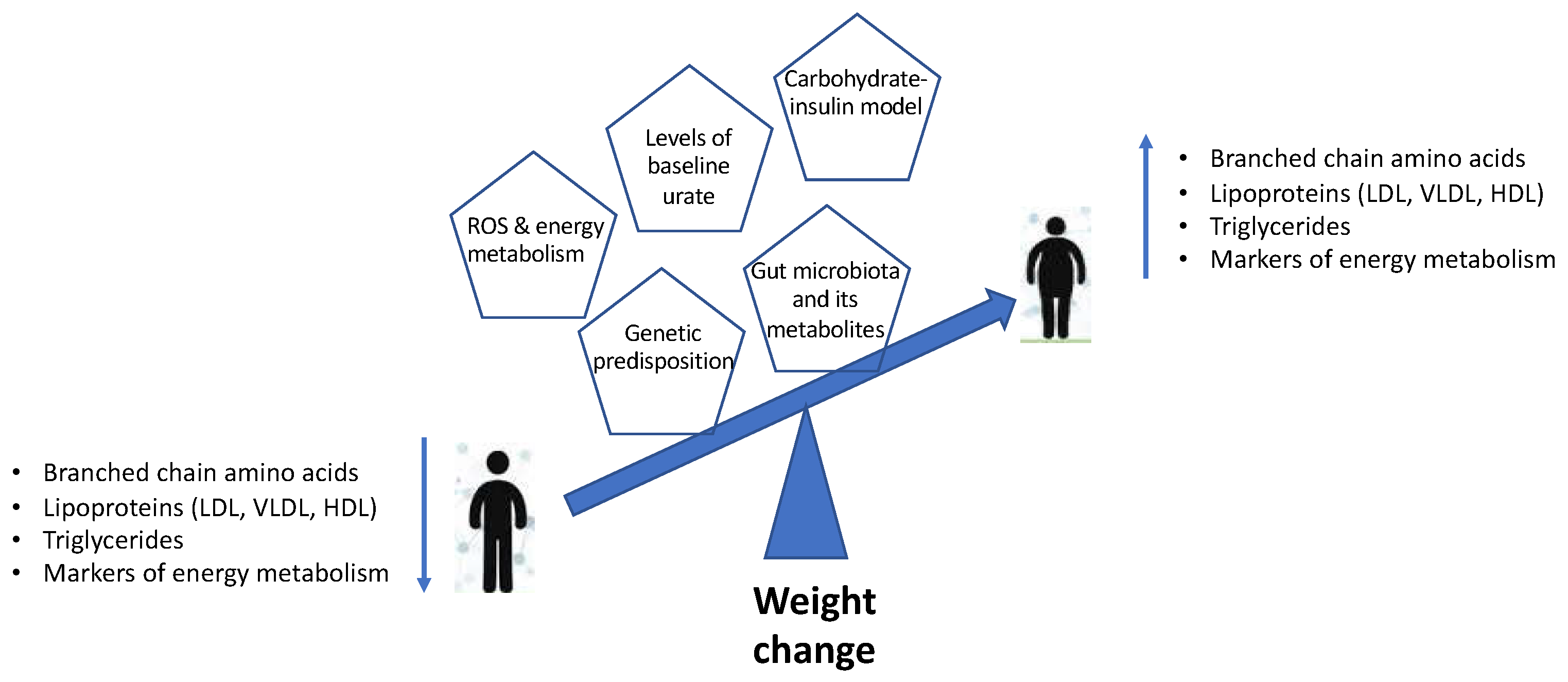
2. Metabolic Profile in Weight Change: What is Known So Far?
3. Metabolomic Profiling of Weight Change
4. Role of ROS and Mitochondrial Dysfunction
5. Role of Urate
6. Carbohydrate–Insulin Model of Weight Gain
7. Gut-Microbiome-Derived Metabolic Markers of Weight Change
8. Discussion and Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Kushner, R.F.; Kahan, S. Introduction: The State of Obesity in 2017. Med. Clin. N. Am. 2018, 102, 1–11. [Google Scholar] [PubMed]
- Muller, M.J.; Enderle, J.; Bosy-Westphal, A. Changes in Energy Expenditure with Weight Gain and Weight Loss in Humans. Curr. Obes. Rep. 2016, 5, 413–423. [Google Scholar] [CrossRef]
- Westerterp, K.R. Alterations in energy balance with exercise. Am. J. Clin. Nutr. 1998, 68, 974S. [Google Scholar] [CrossRef]
- Drenowatz, C.; Grieve, G.; DeMello, M. Change in energy expenditure and physical activity in response to aerobic and resistance exercise programs. SpringerPlus 2015, 4, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Müller, M.J.; Geisler, C.; Heymsfield, S.B.; Bosy-Westphal, A. Recent advances in understanding body weight homeostasis in humans. F1000Res 2018, 7. [Google Scholar] [CrossRef]
- Abu Bakar, M.H.; Sarmidi, M.R.; Cheng, K.-K.; Khan, A.A.; Suan, C.L.; Huri, H.Z.; Yaakob, H. Metabolomics—The complementary field in systems biology: A review on obesity and type 2 diabetes. Mol. Biosyst. 2015, 11, 1742–1774. [Google Scholar] [CrossRef] [PubMed]
- Park, S.; Sadanala, K.C.; Kim, E. A Metabolomic Approach to Understanding the Metabolic Link between Obesity and Diabetes. Mol. Cells 2015, 38, 587–596. [Google Scholar] [CrossRef]
- Wahl, S.; Yu, Z.; Reinehr, T. Childhood obesity is associated with changes in the serum metabolite profile. Obes. Facts 2012, 5, 660–670. [Google Scholar] [CrossRef]
- Tulipani, S.; Palau-Rodriguez, M.; Andres-Lacueva, C. Biomarkers of Morbid Obesity and Prediabetes by Metabolomic Profiling of Human Discordant Phenotypes. Clin. Chim. Acta 2016, 463, 53–61. [Google Scholar] [CrossRef]
- Moore, L.L.; Chadid, S.; Singer, M.R.; Kreger, B.E.; Denis, G.V. Metabolic health reduces risk of obesity-related cancer in framingham study adults. Cancer Epidemiol. Biomarkers Prev. 2014, 23, 2057–2065. [Google Scholar] [CrossRef]
- Tulipani, S.; Griffin, J.; Andres-Lacueva, C. Metabolomics-Guided Insights on Bariatric Surgery Versus Behavioral Interventions for Weight Loss. Obesity 2016, 24, 2451–2466. [Google Scholar] [CrossRef]
- Yao, J.; Kovalik, J.P.; Lai, O.F.; Lee, P.C.; Eng, A.; Chan, W.H.; Tham, K.W.; Lim, E.; Bee, Y.M.; Tan, H.C. Comprehensive Assessment of the Effects of Sleeve Gastrectomy on Glucose, Lipid, and Amino Acid Metabolism in Asian Individuals with Morbid Obesity. Obes. Surg. 2019, 29, 149–158. [Google Scholar] [CrossRef]
- Piening, B.D.; Zhou, W.; Snyder, M.P. Integrative Personal Omics Profiles during Periods of Weight Gain and Loss. Cell Syst. 2018, 6, 157–170. [Google Scholar] [CrossRef]
- Brennan, A.M.; Benson, M.; Ross, R. Plasma Metabolite Profiles in Response to Chronic Exercise. Med. Sci. Sports Exerc. 2018, 50, 1480–1486. [Google Scholar] [CrossRef]
- Naganuma, R.; Sakurai, M.; Stamler, J. Relation of long-term body weight change to change in lipoprotein particle size in Japanese men and women: The INTERMAP Toyama Study. Atherosclerosis 2009, 206, 282–286. [Google Scholar] [CrossRef]
- Mäntyselkä, P.; Kautiainen, H.; Vanhala, M. Weight change and lipoprotein particle concentration and particle size: A cohort study with 6.5-year follow-up. Atherosclerosis 2012, 223, 239–243. [Google Scholar] [CrossRef]
- Inouye, M.; Kettunen, J.; Peltonen, L. Metabonomic, transcriptomic, and genomic variation of a population cohort. Mol. Syst. Biol. 2010, 6, 441. [Google Scholar] [CrossRef] [PubMed]
- Xu, T.; Holzapfell, C.; Wang-Sattler, R. Effects of smoking and smoking cessation on human serum metabolite profile: Results from the KORA cohort study. BMC Med. 2013, 11, 60. [Google Scholar] [CrossRef]
- Oberbach, A.; Blüher, M.; von Bergen, M. Combined proteomic and metabolomic profiling of serum reveals association of the complement system with obesity and identifies novel markers of body fat mass changes. J. Proteome Res. 2011, 10, 4769–4788. [Google Scholar] [CrossRef] [PubMed]
- Wahl, S.; Vogt, S.; Grallert, H. Multi-omic signature of body weight change: Results from a population-based cohort study. BMC Med 2015, 13, 48. [Google Scholar] [CrossRef] [PubMed]
- Diaz-Vivancos, P.; de Simone, A.; Foyer, C.H. Glutathione—Linking cell proliferation to oxidative stress. Free Radic. Biol. Med. 2015, 89, 1154–1164. [Google Scholar] [CrossRef] [PubMed]
- Ojuka, E.; Andrew, B.; Bezuidenhout, N.; George, S.; Maarman, G.; Madlala, H.P.; Mendham, A.; Osiki, P.O. Measurement of beta-oxidation capacity of biological samples by respirometry: A review of principles and substrates. Am. J. Physiol. Endocrinol. Metab. 2016, 310, E715–E723. [Google Scholar] [CrossRef] [PubMed]
- Pallares-Mendez, R.; Aguilar-Salinas, C.A.; Del Bosque-Plata, L. Metabolomics in diabetes, a review. Ann. Med. 2016, 48, 89–102. [Google Scholar] [CrossRef] [PubMed]
- Ragab, S.M.; Abd Elghaffar, S.K.; Omar, H.M. Effect of a high fat, high sucrose diet on the promotion of non-alcoholic fatty liver disease in male rats: The ameliorative role of three natural compounds. Lipids Health. Dis. 2015, 14, 1. [Google Scholar] [CrossRef]
- Menni, C.; Jackson, M.A.; Pallister, T.; Steves, C.J.; Spector, T.D.; Valdes, A.M. Gut microbiome diversity and high-fibre intake are related to lower long-term weight gain. Int. J. Obes. (Lond.) 2017, 41, 1099–1105. [Google Scholar] [CrossRef]
- Richard, D.; Kefi, K.; Visioli, F. Polyunsaturated fatty acids as antioxidants. Pharmacol. Res. 2008, 57, 451–455. [Google Scholar] [CrossRef] [PubMed]
- Trammell, S.A.; Schmidt, M.S.; Brenner, C. Nicotinamide riboside is uniquely and orally bioavailable in mice and humans. Nat. Commun. 2016, 7, 12948. [Google Scholar] [CrossRef] [PubMed]
- Sigal, R.J.; El-Hashimy, M.; Warram, J.H. Acute postchallenge hyperinsulinemia predicts weight gain: A prospective study. Diabetes 1997, 46, 1025–1029. [Google Scholar] [CrossRef]
- Schwartz, M.W.; Boyko, E.J.; Bogardus, C. Reduced insulin secretion: An independent predictor of body weight gain. J. Clin. Endocrinol. Metab. 1995, 80, 1571–1576. [Google Scholar] [PubMed]
- Zavaroni, I.; Zuccarelli, A.; Reaven, G.M. Can weight gain in healthy, nonobese volunteers be predicted by differences in baseline plasma insulin concentration? J. Clin. Endocrinol. Metab. 1998, 83, 3498–3500. [Google Scholar]
- Gould, A.J.; Williams, D.E.; Byrne, C.D.; Hales, C.N.; Wareham, N.J. Prospective cohort study of the relationship of markers of insulin resistance and secretion with weight gain and changes in regional adiposity. Int. J. Obes. Relat. Metab. Disord. 1999, 23, 1256–1261. [Google Scholar] [CrossRef]
- Mayer-Davis, E.J.; Kirkner, G.J.; Zaccaro, D.J. Metabolic predictors of 5-year change in weight and waist circumference in a triethnic population—The Insulin Resistance Atherosclerosis study. Am. J. Epidemiol. 2003, 157, 592–601. [Google Scholar] [CrossRef]
- McLaughlin, T.; Abbasi, F.; Reaven, G. Differences in insulin resistance do not predict weight loss in response to hypocaloric diets in healthy obese women. J. Clin. Endocrinol. Metab. 1999, 84, 578–581. [Google Scholar] [CrossRef]
- Alikasifoglu, A.; Yordam, N. The metabolic parameters of obese children and the role of hyperinsulinism on weight loss. Eur. J. Pediatr. 1999, 158, 269–270. [Google Scholar] [CrossRef]
- Ludwig, D.S.; Majzoub, J.A.; Roberts, S.B. High glycemic index foods, overeating, and obesity. Pediatrics 1999, 103, E26. [Google Scholar] [CrossRef]
- Pawlak, D.B.; Kushner, J.A.; Ludwig, D.S. Effects of dietary glycaemic index on adiposity, glucose homoeostasis, and plasma lipids in animals. Lancet 2003, 364, 778–785. [Google Scholar] [CrossRef]
- Ludwig, D.S.; Friedman, M.I. Increasing Adiposity Consequence or Cause of Overeating? JAMA-J. Am. Med. Assoc. 2014, 311, 2167–2168. [Google Scholar] [CrossRef]
- Stolz, D.J.; Martin, R.J. Role of insulin in food intake, weight gain and lipid deposition in the Zucker obese rat. J. Nutr. 1982, 112, 997–1002. [Google Scholar] [CrossRef]
- Pawlak, D.B.; Bryson, J.M.; Brand-Miller, J.C. High glycemic index starch promotes hypersecretion of insulin and higher body fat in rats without affecting insulin sensitivity. J. Nutr. 2001, 131, 99–104. [Google Scholar] [CrossRef]
- Gardner, C.D.; Trepanowski, J.F.; Del Gobbo, L.C. Effect of low-fat vs low-carbohydrate diet on 12-month weight loss in overweight adults and the association with genotype pattern or insulin secretion: The DIETFITS randomized clinical trial (vol 319, pg 667, 2018). JAMA-J. Am. Med. Assoc. 2018, 319, 1728. [Google Scholar] [CrossRef]
- Pittas, A.G.; Das, S.K.; Roberts, S.B. A low-glycemic load diet facilitates greater weight loss in over-weight adults with high insulin secretion but not in overweight adults with low insulin secretion in the CALERIE trial. Diabetes Care 2005, 28, 2939–2941. [Google Scholar] [CrossRef] [PubMed]
- Hron, B.M.; Ebbeling, C.B.; Ludwig, D.S. Relationship of insulin dynamics to body composition and resting energy expenditure following weight loss. Obesity 2015, 23, 2216–2222. [Google Scholar] [CrossRef] [PubMed]
- Whitlock, G.; Lewington, S.; Whitlock, G. Body-mass index and cause-specific mortality in 900 000 adults: Collaborative analyses of 57 prospective studies. Lancet 2009, 373, 1083–1096. [Google Scholar] [PubMed]
- Chaput, J.P.; Tremblay, A.; Ludwig, D.S. A novel interaction between dietary composition and insulin secretion: Effects on weight gain in the Quebec Family Study. Am. J. Clin. Nutr. 2008, 87, 303–309. [Google Scholar] [CrossRef] [PubMed]
- Brand-Miller, J.C.; Thomas, M.; Colagiuri, S. Physiological validation of the concept of glycemic load in lean young adults. J. Nutr. 2003, 133, 2728–2732. [Google Scholar] [CrossRef] [PubMed]
- Ebbeling, C.B.; Swain, J.F.; Ludwig, D.S. Effects of Dietary Composition on Energy Expenditure During Weight-Loss Maintenance. JAMA-J. Am. Med. Assoc. 2012, 307, 2627–2634. [Google Scholar] [CrossRef] [PubMed]
- Ebbeling, C.B. Effects of a low-glycemic load vs low-fat diet in obese young adults: A randomized trial (vol 297, pg 2092, 2007). JAMA-J. Am. Med. Assoc. 2007, 298, 627. [Google Scholar]
- Jensen, M.V.; Gooding, J.R.; Ferdaoussi, M.; Dai, X.Q.; Peterson, B.S.; MacDonald, P.E.; Newgard, C.B. Metabolomics applied to islet nutrient sensing mechanisms. Diabetes Obes. Metab. 2017, 19 (Suppl. 1), 90–94. [Google Scholar] [CrossRef] [PubMed]
- Gooding, J.R.; Jensen, M.V.; Newgard, C.B. Adenylosuccinate Is an Insulin Secretagogue Derived from Glucose-Induced Purine Metabolism. Cell. Rep. 2015, 13, 157–167. [Google Scholar] [CrossRef]
- Astley, C.M.; Todd, J.N.; Florez, J.C. Genetic Evidence That Carbohydrate-Stimulated Insulin Secretion Leads to Obesity. Clin. Chem. 2018, 64, 192–200. [Google Scholar] [CrossRef]
- Rahman, V.J. Low-Fat vs. Low-Carbohydrate Diets and Weight Loss. JAMA-J. Am. Med. Assoc. 2018, 320, 202. [Google Scholar] [CrossRef] [PubMed]
- Ebbeling, C.B.; Feldman, H.A.; Ludwig, D.S. Effects of a low carbohydrate diet on energy expenditure during weight loss maintenance: Randomized trial. BMJ-Br. Med. J. 2018, 363, k4583. [Google Scholar] [CrossRef] [PubMed]
- Baquero, F.; Nombela, C. The microbiome as a human organ. Clin. Microbiol. Infect. 2012, 18, 2–4. [Google Scholar] [CrossRef] [PubMed]
- Okeke, F.; Roland, B.C.; Mullin, G.E. The role of the gut microbiome in the pathogenesis and treatment of obesity. Glob. Adv. Health. Med. 2014, 3, 44–57. [Google Scholar] [CrossRef] [PubMed]
- Ley, R.E.; Turnbaugh, P.J.; Gordon, J.I. Microbial ecology: Human gut microbes associated with obesity. Nature 2006, 444, 1022–1023. [Google Scholar] [CrossRef]
- Turnbaugh, P.J.; Ley, R.E.; Gordon, J.I. An obesity-associated gut microbiome with increased capacity for energy harvest. Nature 2006, 444, 1027–1031. [Google Scholar] [CrossRef]
- Turnbaugh, P.J.; Hamady, M.; Gordon, J.I. A core gut microbiome in obese and lean twins. Nature 2009, 457, 480–484. [Google Scholar] [CrossRef]
- Cani, P.D.; Bibiloni, R.; Burcelin, R. Changes in gut microbiota control metabolic endotoxemia-induced inflammation in high-fat diet-induced obesity and diabetes in mice. Diabetes 2008, 57, 1470–1481. [Google Scholar] [CrossRef]
- Mayengbam, S.; Lambert, J.E.; Reimer, R.A. Impact of dietary fiber supplementation on modulating microbiota-host-metabolic axes in obesity. J. Nutr. Biochem. 2018, 64, 228–236. [Google Scholar] [CrossRef]
- Cani, P.D.; Amar, J.; Burcelin, R. Metabolic endotoxemia initiates obesity and insulin resistance. Diabetes 2007, 56, 1761–1772. [Google Scholar] [CrossRef]
- John, G.K.; Wang, L.; Mullin, G. Dietary Alteration of the Gut Microbiome and Its Impact on Weight and Fat Mass: A Systematic Review and Meta-Analysis. Genes 2018, 9, 167. [Google Scholar] [CrossRef] [PubMed]
- Sanna, S.; van Zuydam, N.R.; McCarthy, M.I. Causal relationships among the gut microbiome, short-chain fatty acids and metabolic diseases. Nat. Genet. 2019, 51, 600–605. [Google Scholar] [CrossRef] [PubMed]
- Tuomainen, M.; Lindstrom, J.; Hanhineva, K. Associations of serum indolepropionic acid, a gut microbiota metabolite, with type 2 diabetes and low-grade inflammation in high-risk individuals. Nutr. Diabetes 2018, 8, 9. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Contreras, M.; Tamayo-Garcia, A.; Buchwald, P. Metabolomics Study of the Effects of Inflammation, Hypoxia, and High Glucose on Isolated Human Pancreatic Islets. J. Proteome Res. 2017, 16, 2294–2306. [Google Scholar] [CrossRef] [PubMed]
- Pallister, T.; Jackson, M.A.; Menni, C. Hippurate as a metabolomic marker of gut microbiome diversity: Modulation by diet and relationship to metabolic syndrome. Sci. Rep. 2017, 7, 4. [Google Scholar] [CrossRef] [PubMed]
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vijay, A.; Valdes, A.M. The Metabolomic Signatures of Weight Change. Metabolites 2019, 9, 67. https://doi.org/10.3390/metabo9040067
Vijay A, Valdes AM. The Metabolomic Signatures of Weight Change. Metabolites. 2019; 9(4):67. https://doi.org/10.3390/metabo9040067
Chicago/Turabian StyleVijay, Amrita, and Ana M Valdes. 2019. "The Metabolomic Signatures of Weight Change" Metabolites 9, no. 4: 67. https://doi.org/10.3390/metabo9040067
APA StyleVijay, A., & Valdes, A. M. (2019). The Metabolomic Signatures of Weight Change. Metabolites, 9(4), 67. https://doi.org/10.3390/metabo9040067