Urea Cycle Related Amino Acids Measured in Dried Bloodspots Enable Long-Term In Vivo Monitoring and Therapeutic Adjustment

 , , ,
, , ,  and
and
Abstract
1. Introduction
2. Results
2.1. Dried Blood Spot Extraction
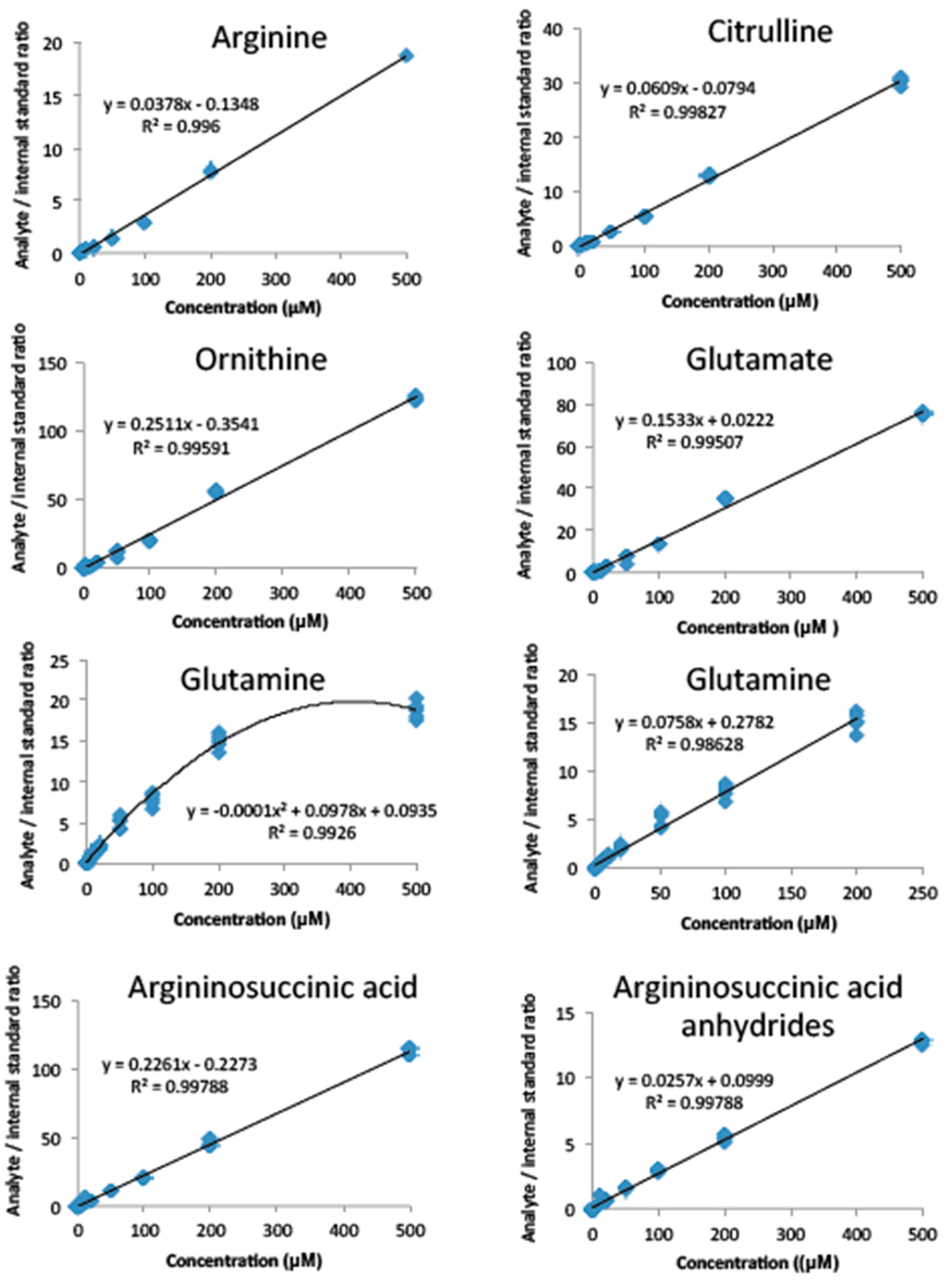
2.2. Method Validation
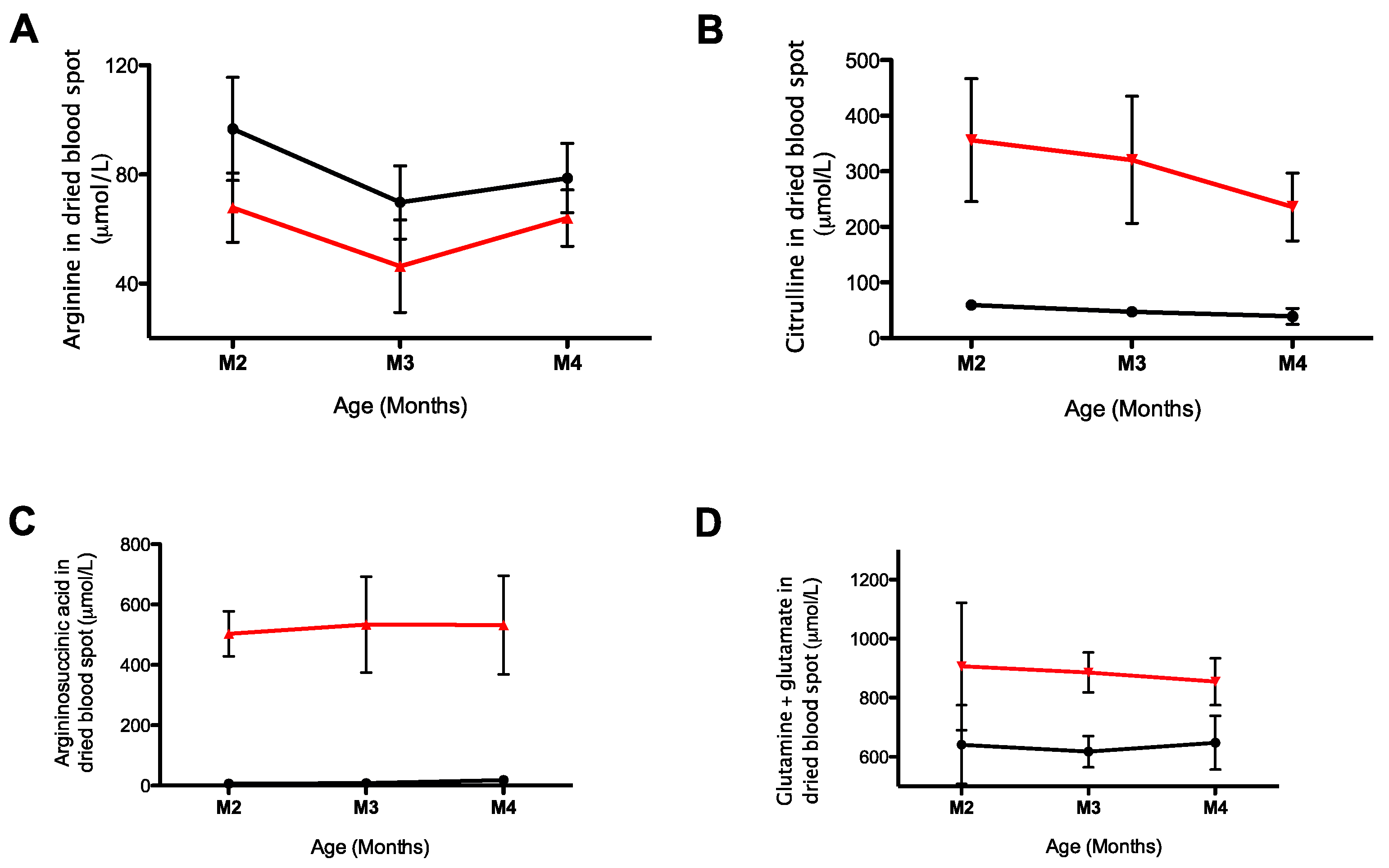
2.3. Monitoring of Urea Cycle Related Amino Acids in AslNeo/Neo Mice
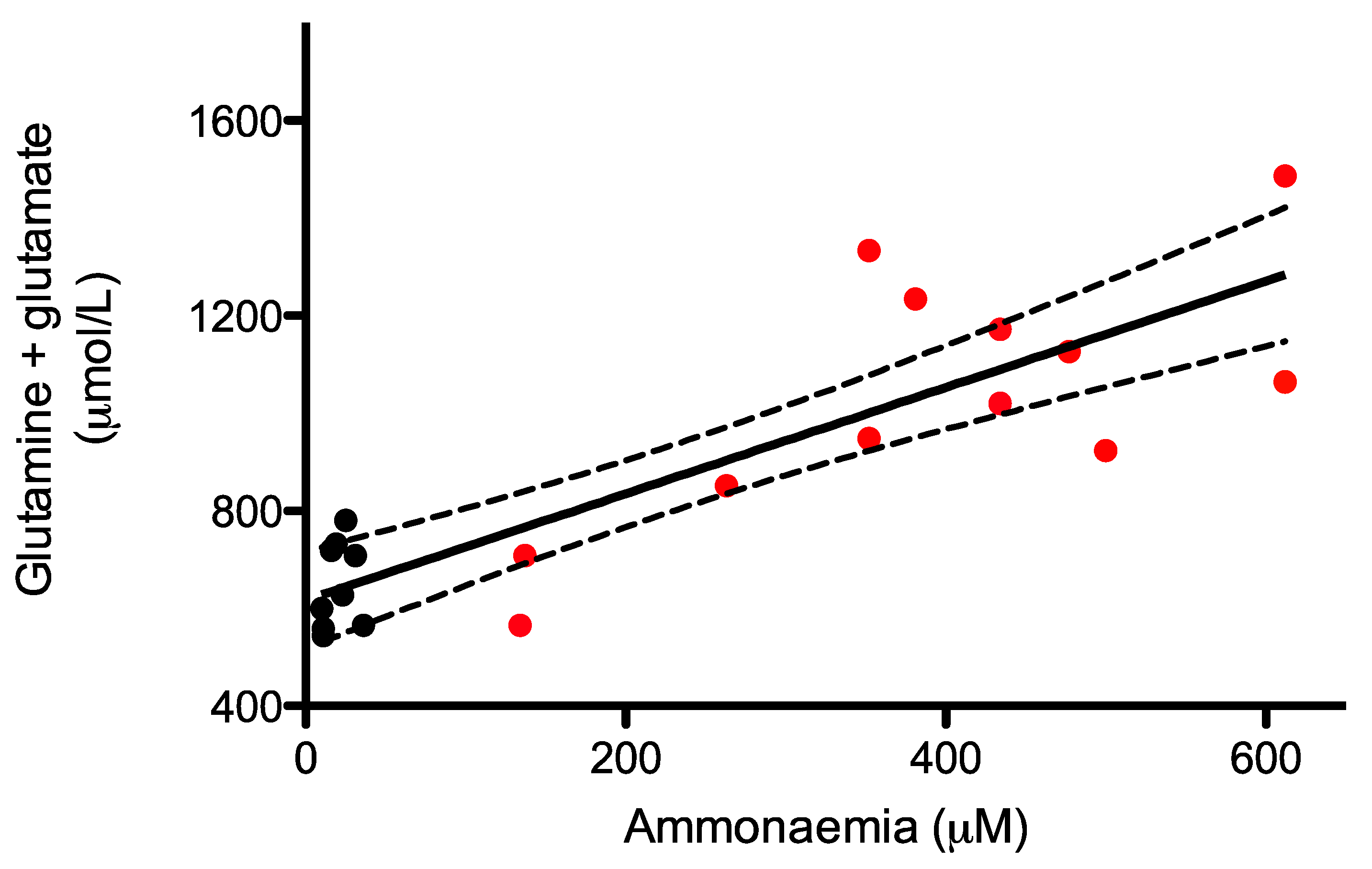
2.4. Monitoring of Glutamine and Glutamate Levels in Dried Blood Spots as A Surrogate of Ammonia Levels in AslNeo/Neo Mice
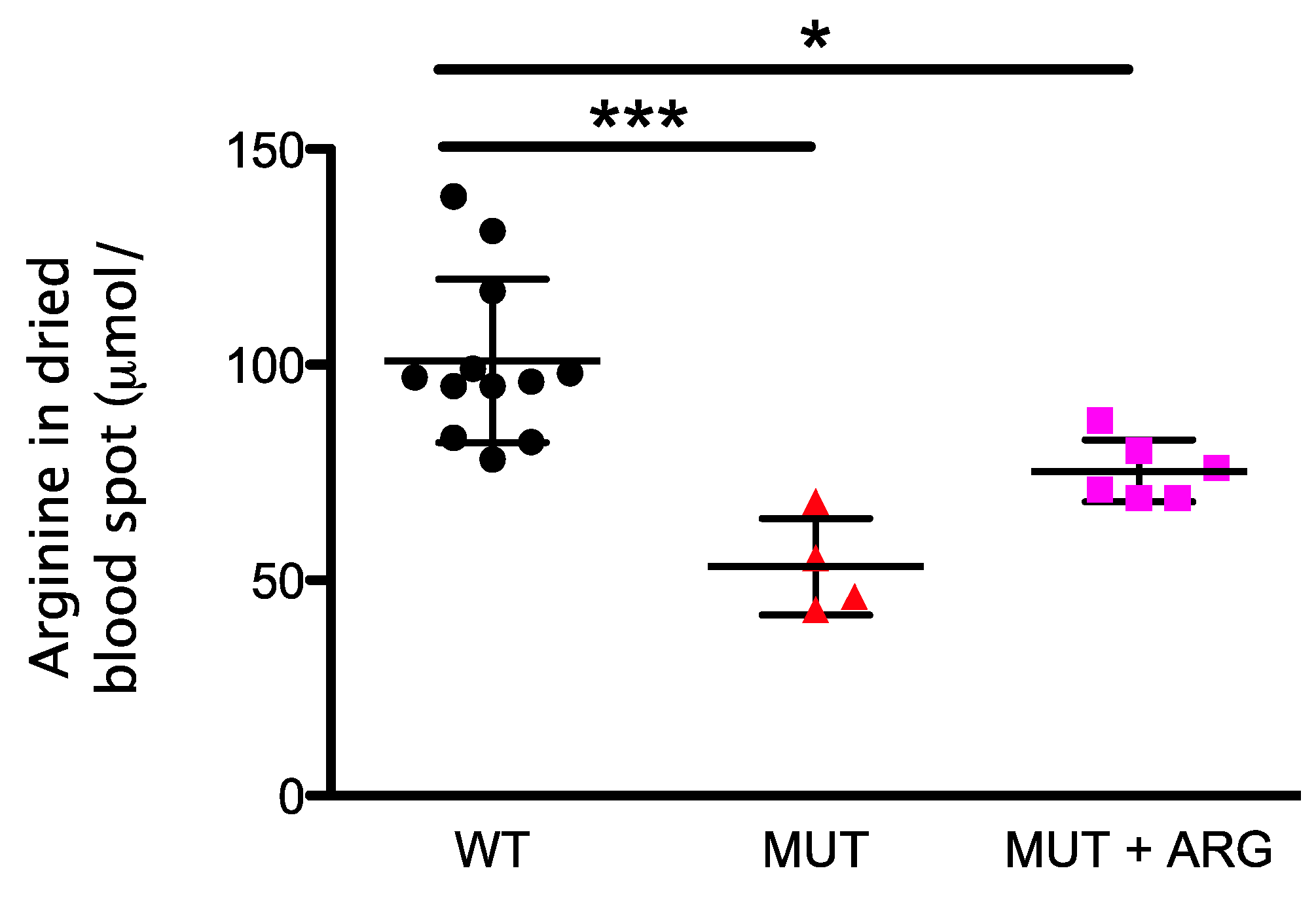
2.5. Monitoring of Arginine Supplementation in AslNeo/Neo Mice
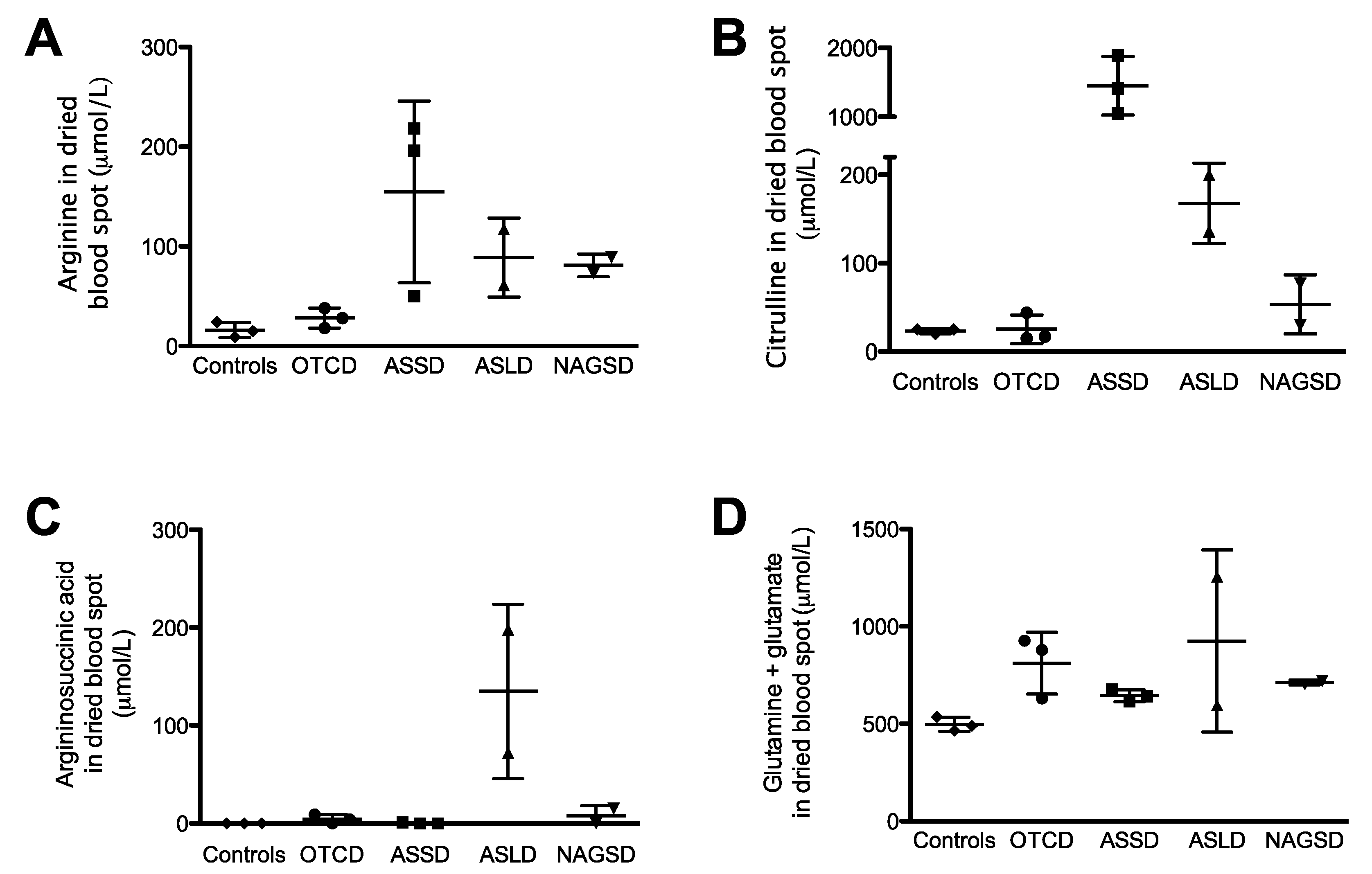
2.6. Urea Cycle Related Amino Acid Levels in Dried Blood Spots from Patients with Urea Cycle Defects
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Blood Spot Collection and Sample Preparation
4.3. LC-MSMS
4.4. Analysis
4.5. Animals
4.6. Ethics
4.7. Statistics
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Wang, C.; Zhu, H.; Zhang, W.; Song, F.; Liu, Z.; Liu, S. Second-tier test for quantification of underivatized amino acids in dry blood spot for metabolic diseases in newborn screening. Amino Acids 2013, 44, 661–671. [Google Scholar] [CrossRef] [PubMed]
- Wilson, M.P.; Footitt, E.J.; Papandreou, A.; Uudelepp, M.L.; Pressler, R.; Stevenson, D.C.; Gabriel, C.; McSweeney, M.; Baggot, M.; Burke, D.; et al. An LC-MS/MS-Based Method for the Quantification of Pyridox (am) ine 5’-Phosphate Oxidase Activity in Dried Blood Spots from Patients with Epilepsy. Anal. Chem. 2017, 89, 8892–8900. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Yi, F.; Kumar, A.B.; Kumar Chennamaneni, N.; Hong, X.; Scott, C.R.; Gelb, M.H.; Turecek, F. Multiplex Tandem Mass Spectrometry Enzymatic Activity Assay for Newborn Screening of the Mucopolysaccharidoses and Type 2 Neuronal Ceroid Lipofuscinosis. Clin. Chem. 2017, 63, 1118–1126. [Google Scholar] [CrossRef] [PubMed]
- De Silva, V.; Oldham, C.D.; May, S.W. L-Phenylalanine concentration in blood of phenylketonuria patients: A modified enzyme colorimetric assay compared with amino acid analysis, tandem mass spectrometry, and HPLC methods. Clin. Chem. Lab. Med. 2010, 48, 1271–1279. [Google Scholar] [CrossRef] [PubMed]
- Bartl, J.; Chrastina, P.; Krijt, J.; Hodik, J.; Peskova, K.; Kozich, V. Simultaneous determination of cystathionine, total homocysteine, and methionine in dried blood spots by liquid chromatography/tandem mass spectrometry and its utility for the management of patients with homocystinuria. Clin. Chim. Acta. 2014, 437, 211–217. [Google Scholar] [CrossRef] [PubMed]
- Alodaib, A.; Carpenter, K.; Wiley, V.; Sim, K.; Christodoulou, J.; Wilcken, B. An improved ultra performance liquid chromatography-tandem mass spectrometry method for the determination of alloisoleucine and branched chain amino acids in dried blood samples. Ann. Clin. Biochem. 2011, 48, 468–470. [Google Scholar] [CrossRef] [PubMed]
- Haberle, J.; Burlina, A.; Chakrapani, A.; Dixon, M.; Karall, D.; Lindner, M.; Mandel, H.; Martinelli, D.; Pintos-Morell, G.; Santer, R.; et al. Suggested guidelines for the diagnosis and management of urea cycle disorders: First revision. J. Inherit. Metab. Dis. 2019. [Google Scholar] [CrossRef] [PubMed]
- Lee, B.; Diaz, G.A.; Rhead, W.; Lichter-Konecki, U.; Feigenbaum, A.; Berry, S.A.; Le Mons, C.; Bartley, J.A.; Longo, N.; Nagamani, S.C.; et al. Blood ammonia and glutamine as predictors of hyperammonemic crises in patients with urea cycle disorder. Genet. Med. 2015, 17, 561–568. [Google Scholar] [CrossRef] [PubMed]
- McHugh, D.; Cameron, C.A.; Abdenur, J.E.; Abdulrahman, M.; Adair, O.; Al Nuaimi, S.A.; Ahlman, H.; Allen, J.J.; Antonozzi, I.; Archer, S.; et al. Clinical validation of cutoff target ranges in newborn screening of metabolic disorders by tandem mass spectrometry: A worldwide collaborative project. Genet. Med. 2011, 13, 230–254. [Google Scholar] [CrossRef] [PubMed]
- Chace, D.H.; Kalas, T.A.; Naylor, E.W. Use of tandem mass spectrometry for multianalyte screening of dried blood specimens from newborns. Clin. Chem. 2003, 49, 1797–1817. [Google Scholar] [CrossRef] [PubMed]
- Moore, S.; Spackman, D.H.; Stein, W.H. Automatic recording apparatus for use in the chromatography of amino acids. Fed. Proc. 1958, 17, 1107–1115. [Google Scholar] [PubMed]
- Dale, Y.; Mackey, V.; Mushi, R.; Nyanda, A.; Maleque, M.; Ike, J. Simultaneous measurement of phenylalanine and tyrosine in phenylketonuric plasma and dried blood by high-performance liquid chromatography. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2003, 788, 1–8. [Google Scholar] [CrossRef]
- Piraud, M.; Vianey-Saban, C.; Petritis, K.; Elfakir, C.; Steghens, J.P.; Morla, A.; Bouchu, D. ESI-MS/MS analysis of underivatised amino acids: A new tool for the diagnosis of inherited disorders of amino acid metabolism. Fragmentation study of 79 molecules of biological interest in positive and negative ionisation mode. Rapid Commun. Mass Spectrom. 2003, 17, 1297–1311. [Google Scholar] [CrossRef] [PubMed]
- Waterval, W.A.; Scheijen, J.L.; Ortmans-Ploemen, M.M.; Habets-van der Poel, C.D.; Bierau, J. Quantitative UPLC-MS/MS analysis of underivatised amino acids in body fluids is a reliable tool for the diagnosis and follow-up of patients with inborn errors of metabolism. Clin. Chim. Acta 2009, 407, 36–42. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Zhang, W.; Song, F.; Liu, Z.; Liu, S. A simple method for the analysis by MS/MS of underivatized amino acids on dry blood spots from newborn screening. Amino Acids 2012, 42, 1889–1895. [Google Scholar] [CrossRef] [PubMed]
- Kruve, A.; Leito, I.; Herodes, K. Combating matrix effects in LC/ESI/MS: The extrapolative dilution approach. Anal. Chim. Acta 2009, 651, 75–80. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Tse, F.L. Dried blood spot sampling in combination with LC-MS/MS for quantitative analysis of small molecules. Biomed. Chromatogr. 2010, 24, 49–65. [Google Scholar] [CrossRef] [PubMed]
- Walter, J.H.; Patterson, A.; Till, J.; Besley, G.T.; Fleming, G.; Henderson, M.J. Bloodspot acylcarnitine and amino acid analysis in cord blood samples: Efficacy and reference data from a large cohort study. J. Inherit. Metab. Dis. 2009, 32, 95–101. [Google Scholar] [CrossRef] [PubMed]
- Odoardi, S.; Anzillotti, L.; Strano-Rossi, S. Simplifying sample pretreatment: Application of dried blood spot (DBS) method to blood samples, including postmortem, for UHPLC-MS/MS analysis of drugs of abuse. Forensic Sci. Int. 2014, 243, 61–67. [Google Scholar] [CrossRef] [PubMed]
- Bloom, K.; Meyers, G.D.; Bennett, M.J. D.; Bennett, J. A Quantitative Method for the Measurement of Dried Blood Spot Amino Acids Using Ultra-Performance Liquid Chromatography. J. Appl. Lab. Med. 2016, 1, 271–279. [Google Scholar] [CrossRef]
- Moretti, F.; Birarelli, M.; Carducci, C.; Pontecorvi, A.; Antonozzi, I. Simultaneous high-performance liquid chromatographic determination of amino acids in a dried blood spot as a neonatal screening test. J. Chromatogr. 1990, 511, 131–136. [Google Scholar] [CrossRef]
- Nagy, K.; Takats, Z.; Pollreisz, F.; Szabo, T.; Vekey, K. Direct tandem mass spectrometric analysis of amino acids in dried blood spots without chemical derivatization for neonatal screening. Rapid Commun. Mass Spectrom. 2003, 17, 983–990. [Google Scholar] [CrossRef] [PubMed]
- Piraud, M.; Vianey-Saban, C.; Petritis, K.; Elfakir, C.; Steghens, J.P.; Bouchu, D. Ion-pairing reversed-phase liquid chromatography/electrospray ionization mass spectrometric analysis of 76 underivatized amino acids of biological interest: A new tool for the diagnosis of inherited disorders of amino acid metabolism. Rapid Commun. Mass Spectrom. 2005, 19, 1587–1602. [Google Scholar] [CrossRef] [PubMed]
- Wuyts, B.; Stove, V.; Goossens, L. Critical sample pretreatment in monitoring dried blood spot citrulline. Clin. Chim. Acta 2007, 386, 105–109. [Google Scholar] [CrossRef] [PubMed]
- Footitt, E.J.; Clayton, P.T.; Mills, K.; Heales, S.J.; Neergheen, V.; Oppenheim, M.; Mills, P.B. Measurement of plasma B6 vitamer profiles in children with inborn errors of vitamin B6 metabolism using an LC-MS/MS method. J. Inherit. Metab. Dis. 2013, 36, 139–145. [Google Scholar] [CrossRef] [PubMed]
- Zytkovicz, T.H.; Fitzgerald, E.F.; Marsden, D.; Larson, C.A.; Shih, V.E.; Johnson, D.M.; Strauss, A.W.; Comeau, A.M.; Eaton, R.B.; Grady, G.F. Tandem mass spectrometric analysis for amino, organic, and fatty acid disorders in newborn dried blood spots: A two-year summary from the New England Newborn Screening Program. Clin. Chem. 2001, 47, 1945–1955. [Google Scholar] [PubMed]
- Baruteau, J.; Perocheau, D.P.; Hanley, J.; Lorvellec, M.; Rocha-Ferreira, E.; Karda, R.; Ng, J.; Suff, N.; Diaz, J.A.; Rahim, A.A.; et al. Argininosuccinic aciduria fosters neuronal nitrosative stress reversed by Asl gene transfer. Nat. Commun. 2018, 9, 3505. [Google Scholar] [CrossRef] [PubMed]
- Baruteau, J.; Jameson, E.; Morris, A.A.; Chakrapani, A.; Santra, S.; Vijay, S.; Kocadag, H.; Beesley, C.E.; Grunewald, S.; Murphy, E.; et al. Expanding the phenotype in argininosuccinic aciduria: Need for new therapies. J. Inherit. Metab. Dis. 2017, 40, 357–368. [Google Scholar] [CrossRef] [PubMed]
- Yunus, Z.M.; Rahman, S.A.; Choy, Y.S.; Keng, W.T.; Ngu, L.H. Pilot study of newborn screening of inborn error of metabolism using tandem mass spectrometry in Malaysia: Outcome and challenges. J. Pediatr. Endocrinol. Metab. 2016, 29, 1031–1039. [Google Scholar] [CrossRef] [PubMed]
- Burrage, L.C.; Jain, M.; Gandolfo, L.; Lee, B.H.; Nagamani, S.C.; Urea Cycle Disorders Consortium. Sodium phenylbutyrate decreases plasma branched-chain amino acids in patients with urea cycle disorders. Mol. Genet. Metab. 2014, 113, 131–135. [Google Scholar] [CrossRef] [PubMed]
- Boenzi, S.; Pastore, A.; Martinelli, D.; Goffredo, B.M.; Boiani, A.; Rizzo, C.; Dionisi-Vici, C. Creatine metabolism in urea cycle defects. J. Inherit. Metab. Dis. 2012, 35, 647–653. [Google Scholar] [CrossRef] [PubMed]
- Molema, F.; Gleich, F.; Burgard, P.; van der Ploeg, A.T.; Summar, M.L.; Chapman, K.A.; Lund, A.M.; Rizopoulos, D.; Kolker, S.; Williams, M.; et al. Decreased plasma l-arginine levels in organic acidurias (MMA and PA) and decreased plasma branched-chain amino acid levels in urea cycle disorders as a potential cause of growth retardation: Options for treatment. Mol. Genet. Metab. 2019, 126, 397–405. [Google Scholar] [CrossRef] [PubMed]
- Lee, B.; Diaz, G.A.; Rhead, W.; Lichter-Konecki, U.; Feigenbaum, A.; Berry, S.A.; Le Mons, C.; Bartley, J.; Longo, N.; Nagamani, S.C.; et al. Glutamine and hyperammonemic crises in patients with urea cycle disorders. Mol. Genet. Metab. 2016, 117, 27–32. [Google Scholar] [CrossRef] [PubMed]
- Schulze, A.; Lindner, M.; Kohlmuller, D.; Olgemoller, K.; Mayatepek, E.; Hoffmann, G.F. Expanded newborn screening for inborn errors of metabolism by electrospray ionization-tandem mass spectrometry: Results, outcome, and implications. Pediatrics 2003, 111, 1399–1406. [Google Scholar] [CrossRef] [PubMed]
- Serra, A.; Gallart-Palau, X.; Wei, J.; Sze, S.K. Characterization of Glutamine Deamidation by Long-Length Electrostatic Repulsion-Hydrophilic Interaction Chromatography-Tandem Mass Spectrometry (LERLIC-MS/MS) in Shotgun Proteomics. Anal. Chem. 2016, 88, 10573–10582. [Google Scholar] [CrossRef] [PubMed]
- Darmaun, D.; Manary, M.J.; Matthews, D.E. A method for measuring both glutamine and glutamate levels and stable isotopic enrichments. Anal. Biochem. 1985, 147, 92–102. [Google Scholar] [CrossRef]
- Emery, J.L.; Howat, A.J.; Variend, S.; Vawter, G.F. Investigation of inborn errors of metabolism in unexpected infant deaths. Lancet 1988, 2, 29–31. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Amino acid Amound Added to Whole Blood | 50 μM | 300 μM | 1500 μM | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Parameter tested | Recovery (%) | Intrabatch CV (%) | Interbatch CV (%) | Recovery (%) | Intrabatch CV (%) | Interbatch CV (%) | Recovery (%) | Intrabatch CV (%) | Interbatch CV (%) |
| Arginine | 99 | 5 | 5 | 98 | 2 | 4 | 97 | 6 | 5 |
| Citrulline | 104 | 2 | 9 | 116 | 3 | 2 | 106 | 7 | 3 |
| Ornithine | 99 | 5 | 3 | 101 | 3 | 2 | 99 | 3 | 3 |
| Glutamate | 101 | 2 | 6 | 107 | 2 | 2 | 102 | 4 | 4 |
| Glutamine | 99 | 1 | 0 | 99 | 2 | 2 | 96 | 2 | 1 |
| Argininosuccinic acid and anhydrides | 74 | 9 | 27 | 110 | 3 | 14 | 103 | 0 | 12 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Baruteau, J.; Khalil, Y.; Grunewald, S.; Zancolli, M.; Chakrapani, A.; Cleary, M.; Davison, J.; Footitt, E.; Waddington, S.N.; Gissen, P.; et al. Urea Cycle Related Amino Acids Measured in Dried Bloodspots Enable Long-Term In Vivo Monitoring and Therapeutic Adjustment. Metabolites 2019, 9, 275. https://doi.org/10.3390/metabo9110275
Baruteau J, Khalil Y, Grunewald S, Zancolli M, Chakrapani A, Cleary M, Davison J, Footitt E, Waddington SN, Gissen P, et al. Urea Cycle Related Amino Acids Measured in Dried Bloodspots Enable Long-Term In Vivo Monitoring and Therapeutic Adjustment. Metabolites. 2019; 9(11):275. https://doi.org/10.3390/metabo9110275
Chicago/Turabian StyleBaruteau, Julien, Youssef Khalil, Stephanie Grunewald, Marta Zancolli, Anupam Chakrapani, Maureen Cleary, James Davison, Emma Footitt, Simon N. Waddington, Paul Gissen, and et al. 2019. "Urea Cycle Related Amino Acids Measured in Dried Bloodspots Enable Long-Term In Vivo Monitoring and Therapeutic Adjustment" Metabolites 9, no. 11: 275. https://doi.org/10.3390/metabo9110275
APA StyleBaruteau, J., Khalil, Y., Grunewald, S., Zancolli, M., Chakrapani, A., Cleary, M., Davison, J., Footitt, E., Waddington, S. N., Gissen, P., & Mills, P. (2019). Urea Cycle Related Amino Acids Measured in Dried Bloodspots Enable Long-Term In Vivo Monitoring and Therapeutic Adjustment. Metabolites, 9(11), 275. https://doi.org/10.3390/metabo9110275