Plasma Metabolic Signature and Abnormalities in HIV-Infected Individuals on Long-Term Successful Antiretroviral Therapy

 ,
, 
Abstract
1. Introduction
2. Results
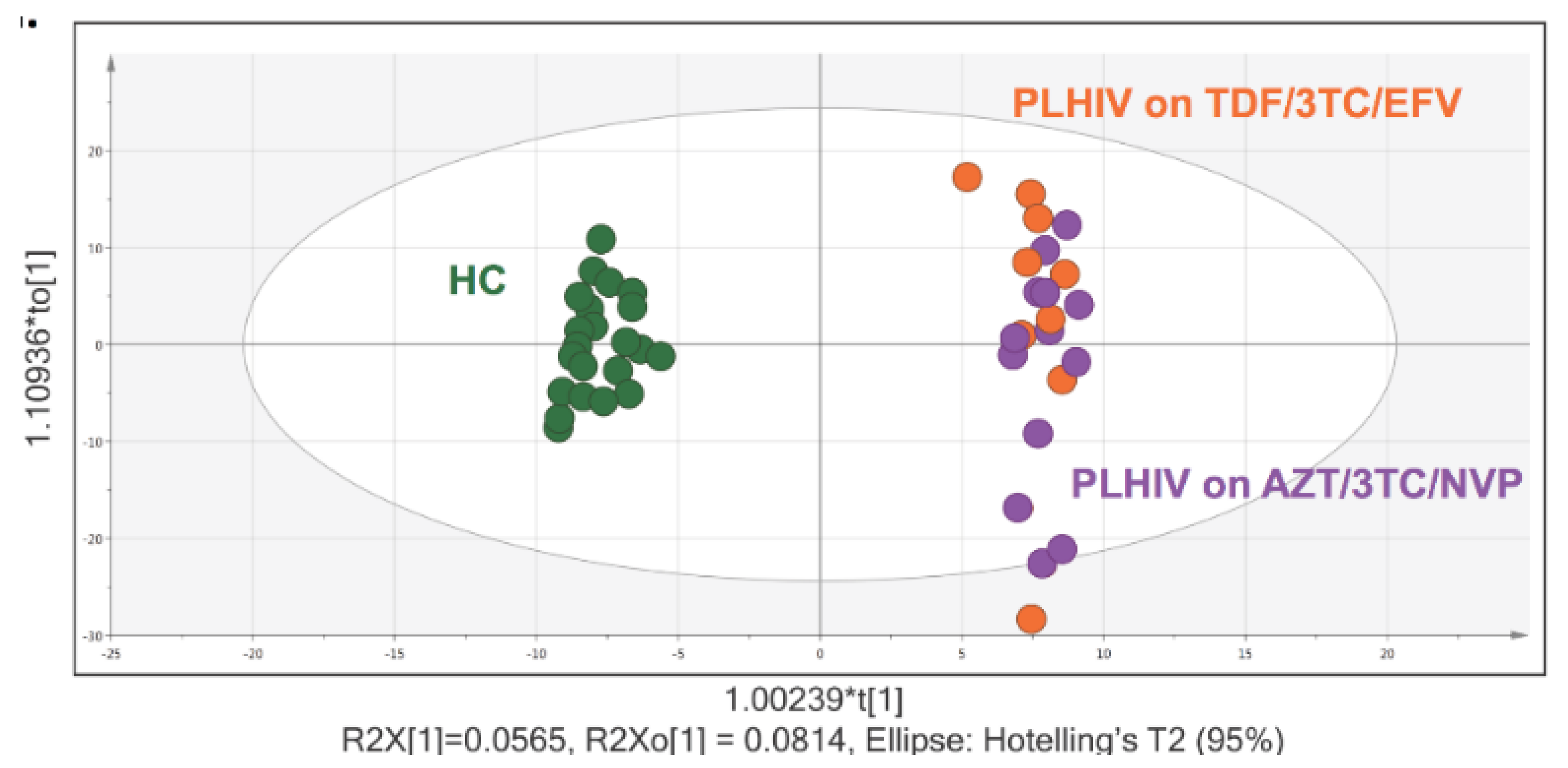
2.1. Cohort Characteristics
2.2. Metabolite Summary
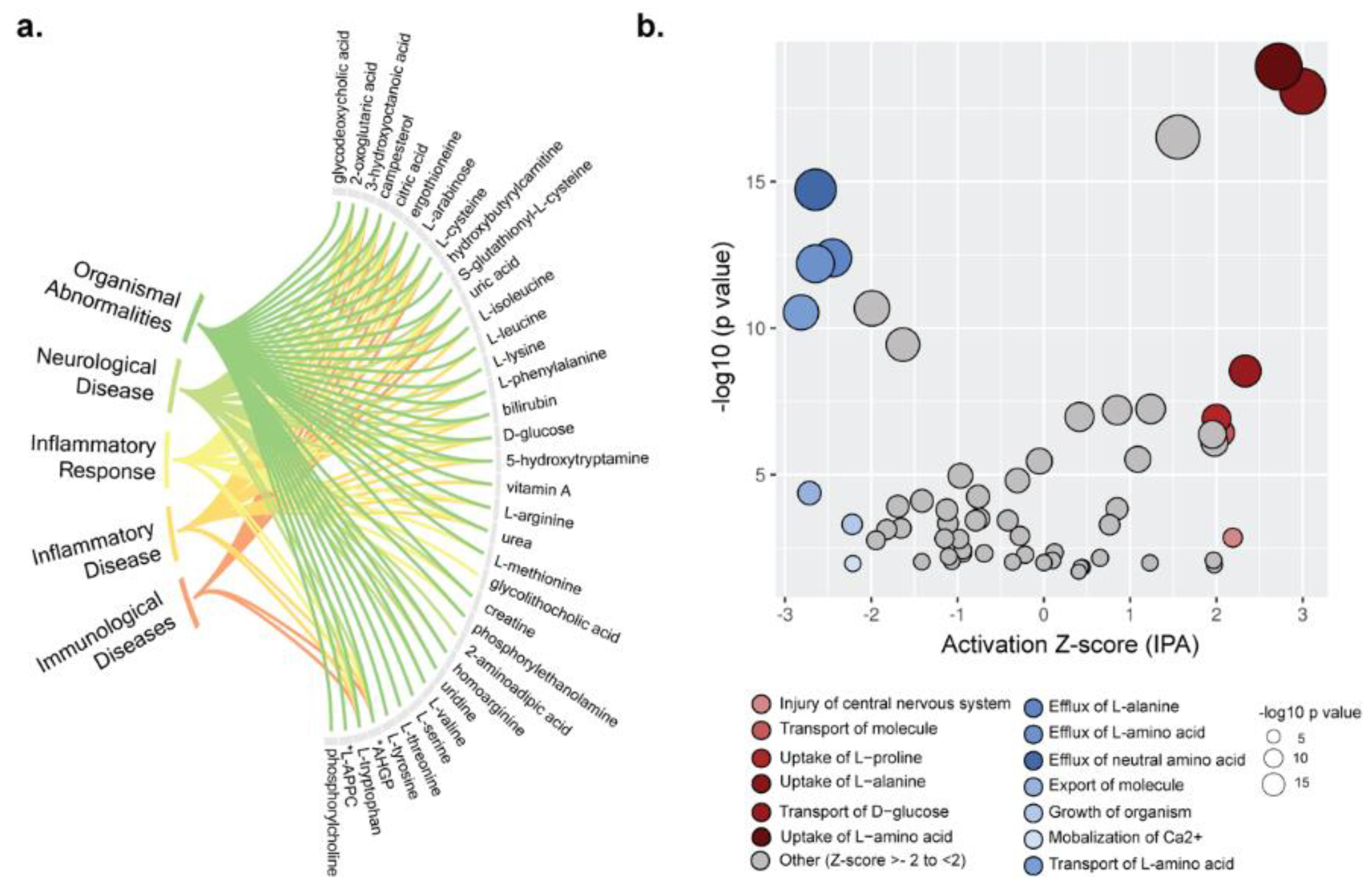
2.3. Disease and Biofunction Predicted by Ingenuity Pathway Analysis (IPA)
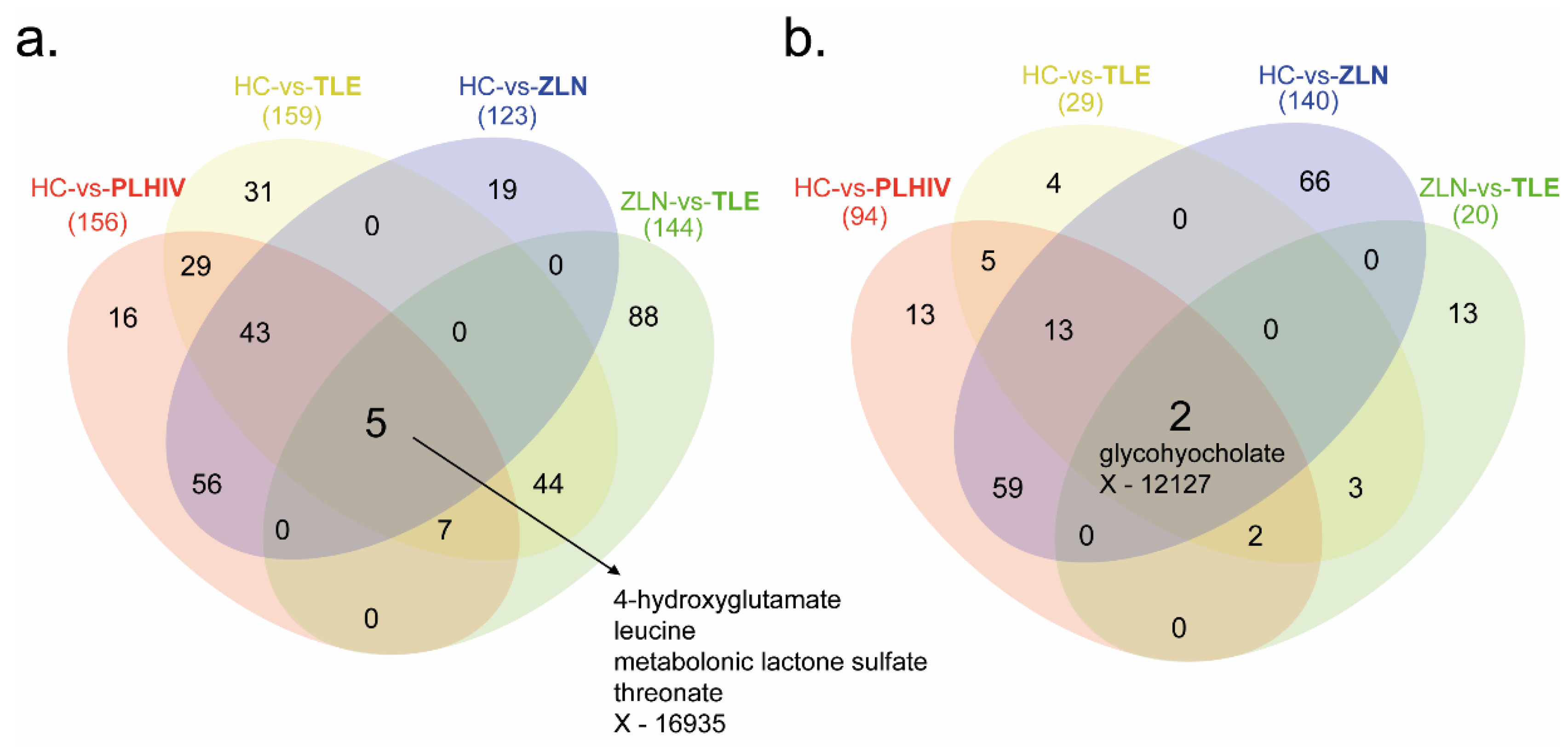
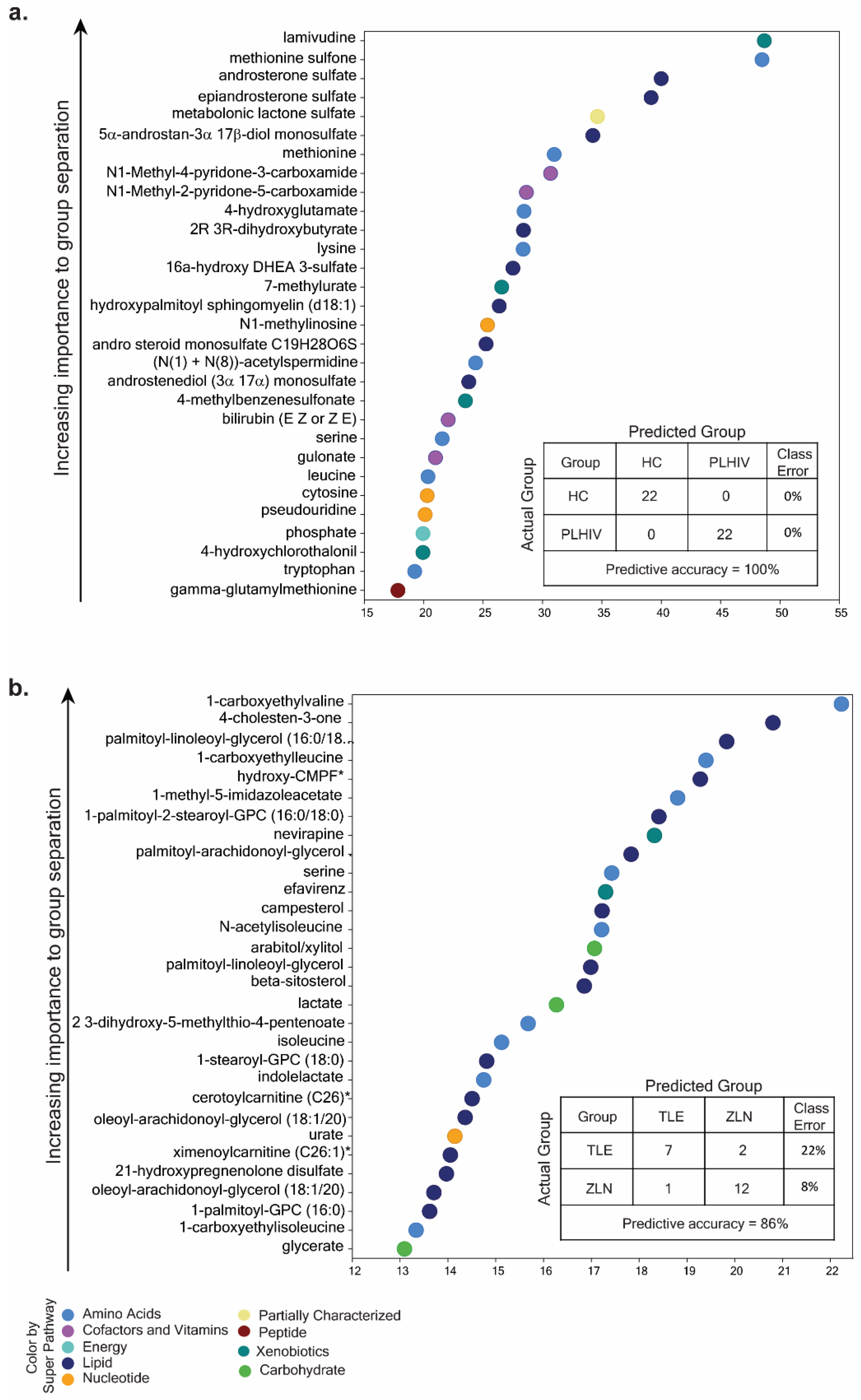
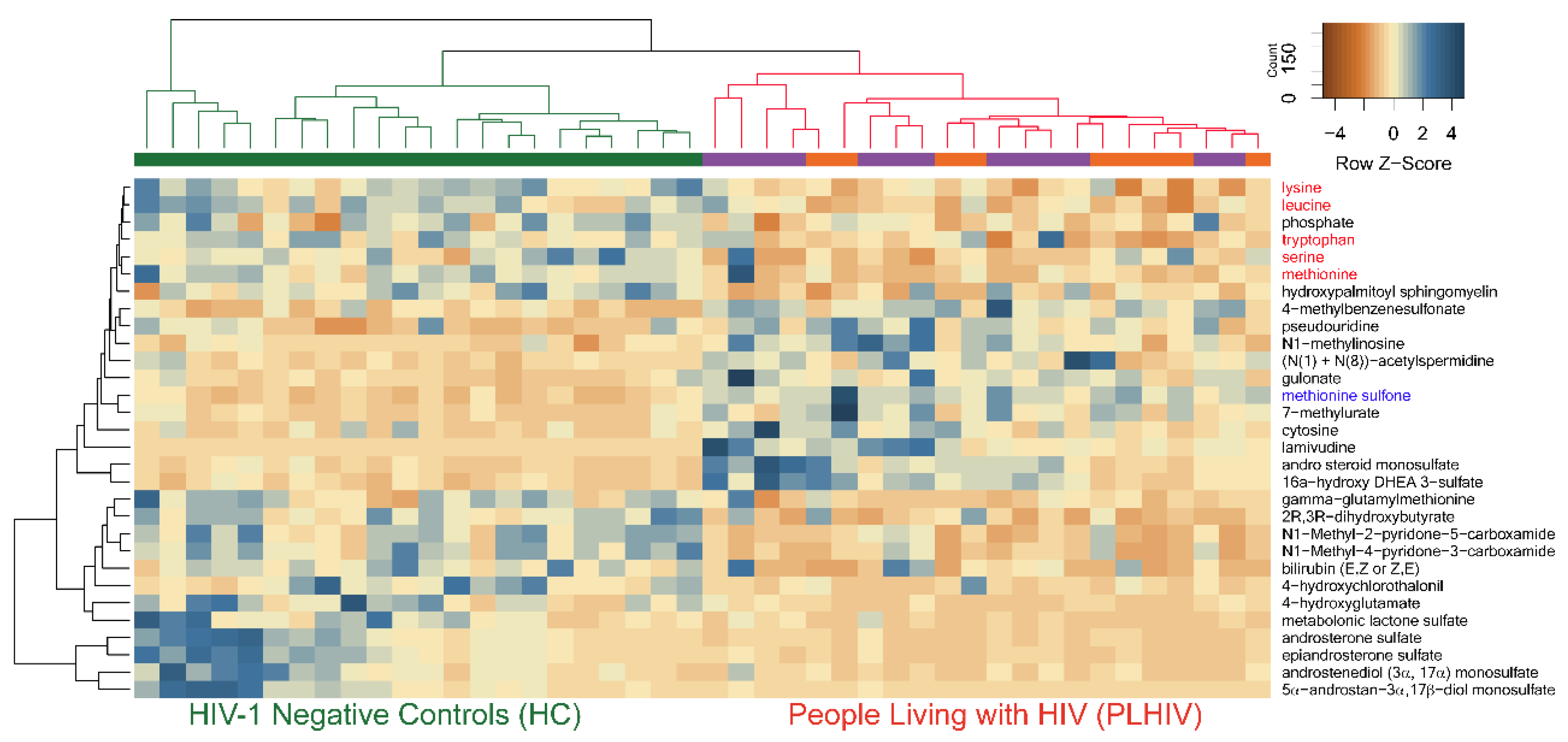
2.4. Identification of Potential Biomarkers
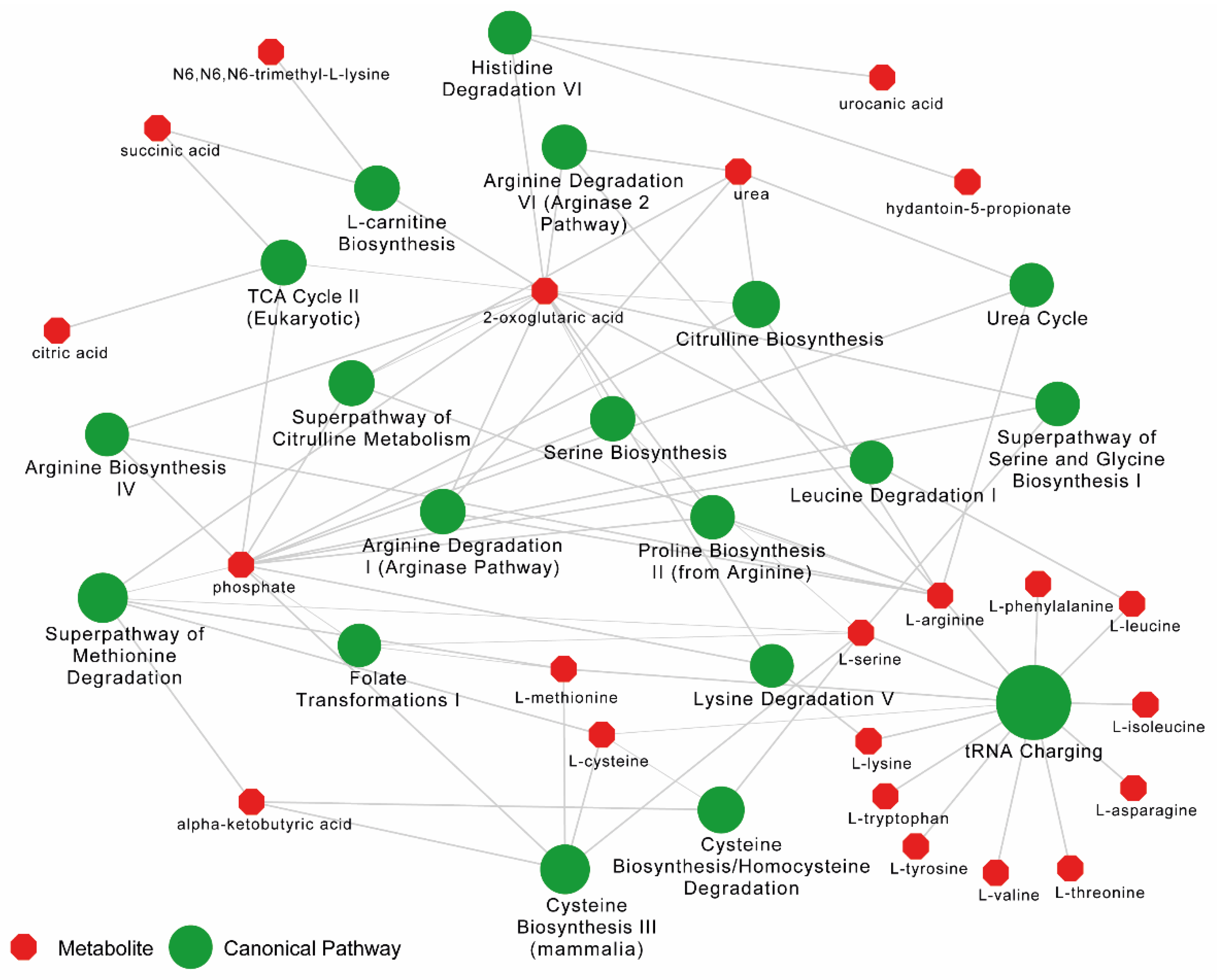
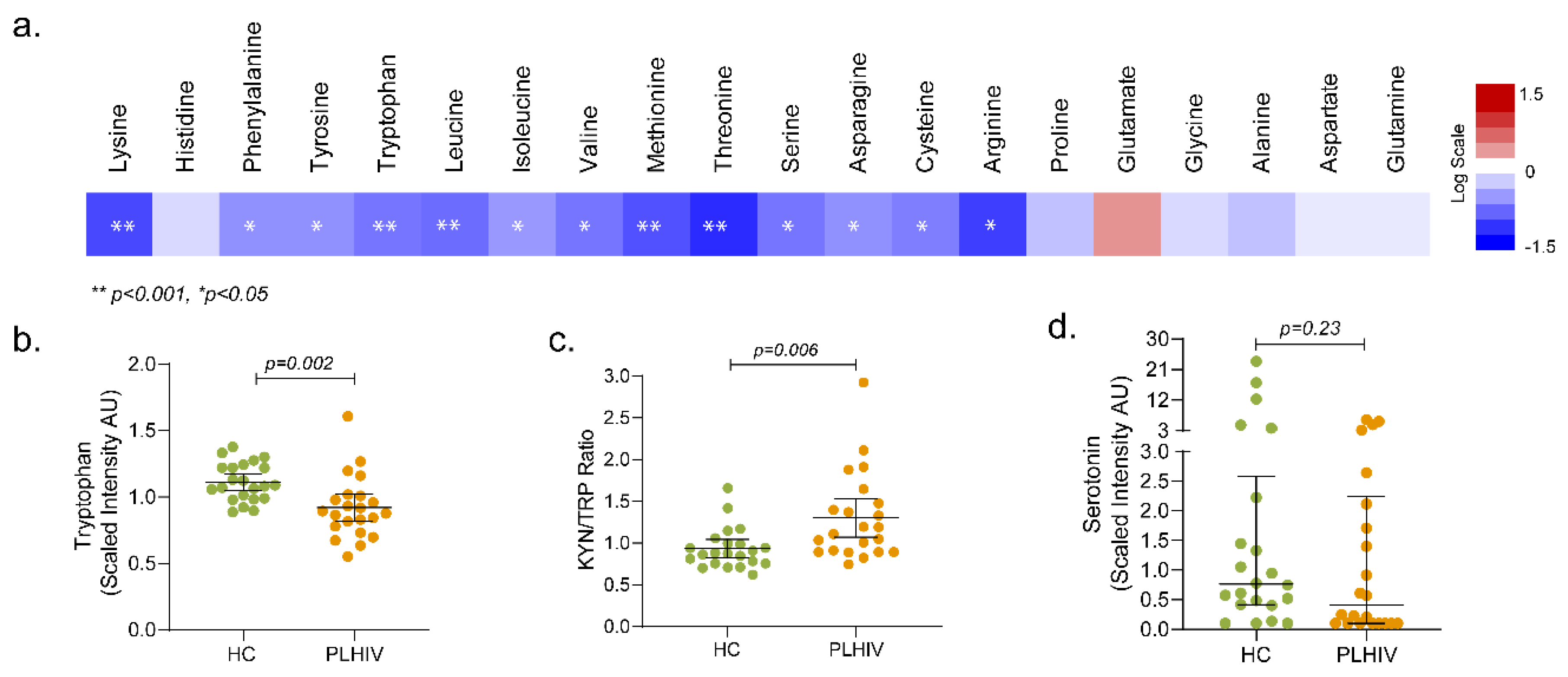
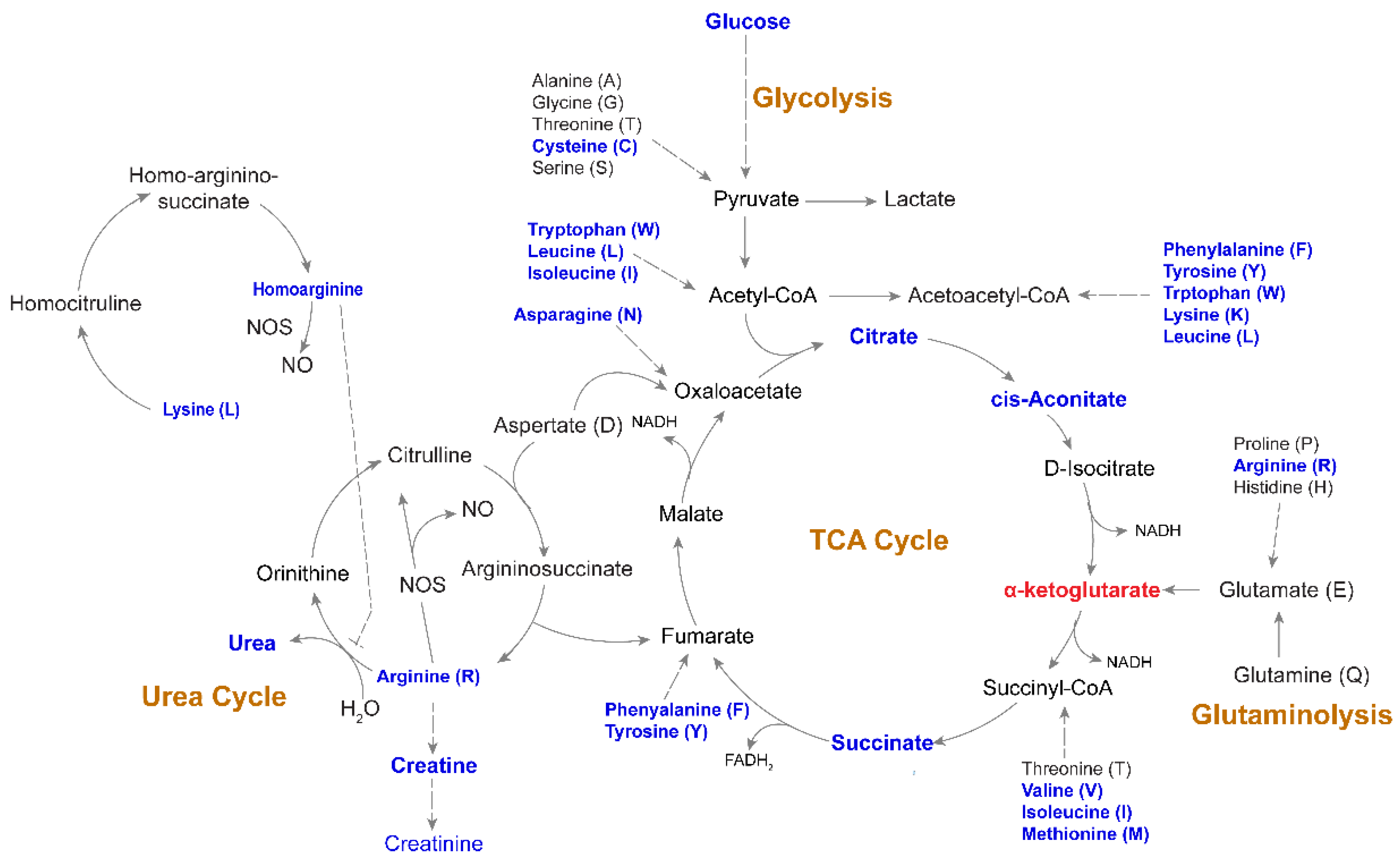
2.5. Altered Amino-Acid Metabolism and Kynurenine-to-Tryptophan (K/T) Ratio in PLHIV
2.6. Altered Energy Metabolism
2.7. Changes in Phospholipids, Ceramides, and Other Complex Lipids
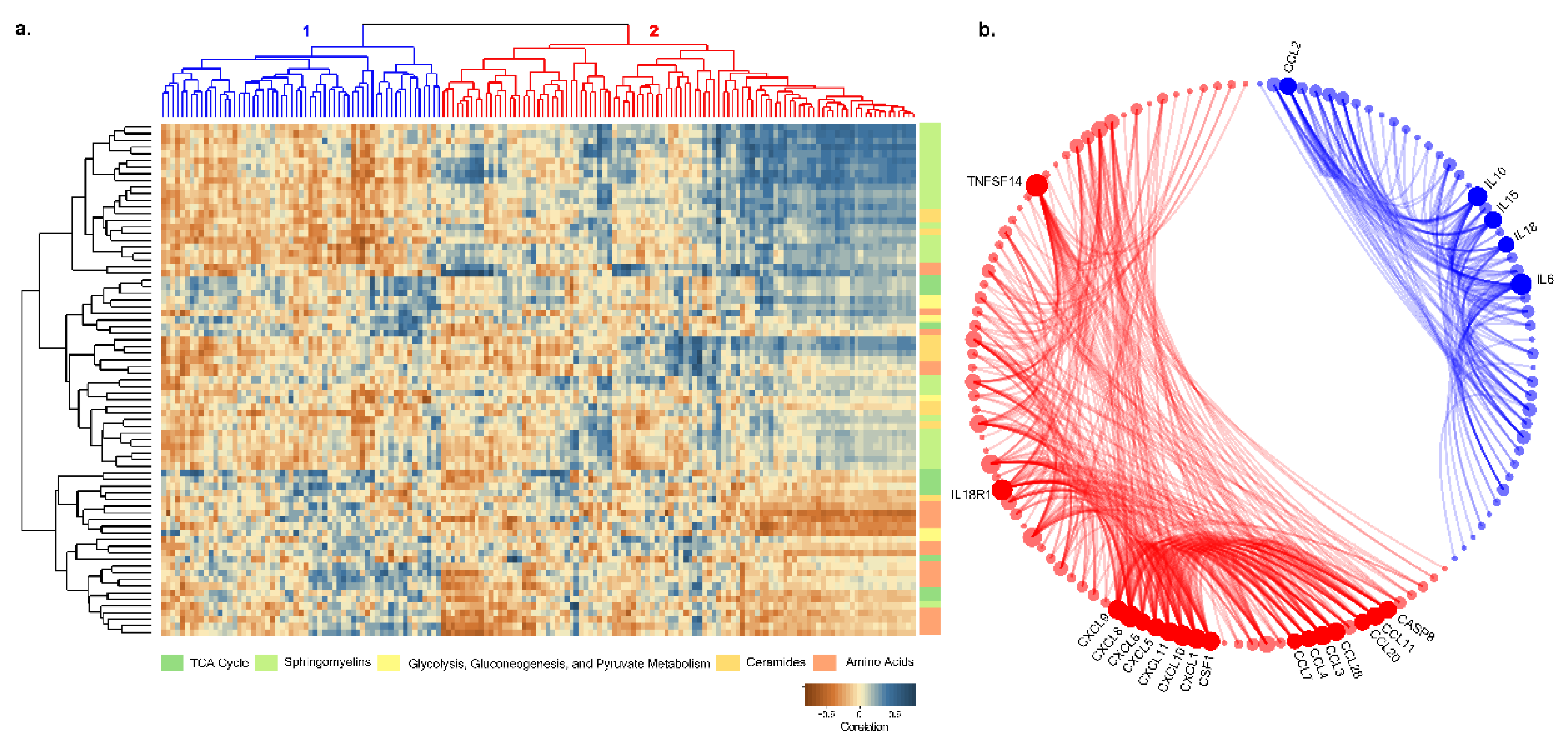
2.8. Plasma Proteomic Profile and Its Correlation with Metabolite
3. Discussion
4. Materials and Methods
4.1. Study Cohorts
4.2. Sample Preparation and Ultra-High-Performance Liquid Chromatography/Mass Spectrometry/Mass Spectrometry (UHPLC/MS/MS)
4.3. Data Extraction, Compound Identification, and Quantification
4.4. Plasma Proteomics Profiling
4.5. Statistical and Bioinformatics Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Deeks, S.G. HIV infection, inflammation, immunosenescence, and aging. Annu. Rev. Med. 2011, 62, 141–155. [Google Scholar] [CrossRef] [PubMed]
- Smit, M.; Brinkman, K.; Geerlings, S.; Smit, C.; Thyagarajan, K.; Sighem, A.; de Wolf, F.; Hallett, T.B. Future challenges for clinical care of an ageing population infected with HIV: A modelling study. Lancet Infect. Dis. 2015, 15, 810–818. [Google Scholar] [CrossRef]
- Scarpellini, B.; Zanoni, M.; Sucupira, M.C.; Truong, H.M.; Janini, L.M.; Segurado, I.D.; Diaz, R.S. Plasma Metabolomics Biosignature According to HIV Stage of Infection, Pace of Disease Progression, Viremia Level and Immunological Response to Treatment. PLoS ONE 2016, 11, e0161920. [Google Scholar] [CrossRef] [PubMed]
- Siegel, M.O.; Borkowska, A.G.; Dubrovsky, L.; Roth, M.; Welti, R.; Roberts, A.D.; Parenti, D.M.; Simon, G.L.; Sviridov, D.; Simmens, S.; et al. HIV infection induces structural and functional changes in high density lipoproteins. Atherosclerosis 2015, 243, 19–29. [Google Scholar] [CrossRef] [PubMed]
- Williams, A.A.; Sitole, L.J.; Meyer, D. HIV/HAART-associated oxidative stress is detectable by metabonomics. Mol. Biosyst. 2017, 13, 2202–2217. [Google Scholar] [CrossRef] [PubMed]
- Reliquet, V.; Chirouze, C.; Allavena, C.; Muret, P.; Peytavin, G.; Andre-Garnier, E.; Bettinger, D.; Ferre, V.; Hoen, B.; Raffi, F. Nevirapine-raltegravir combination, an NRTI and PI/r sparing regimen, as maintenance antiretroviral therapy in virologically suppressed HIV-1-infected patients. Antivir. Ther. 2014, 19, 117–123. [Google Scholar] [CrossRef] [PubMed]
- Neogi, U.; Heylen, E.; Shet, A.; Chandy, S.; Shamsunder, R.; Sonnerborg, A.; Ekstrand, M.L. Long-term efficacy of first line antiretroviral therapy in Indian HIV-1 infected patients: A longitudinal cohort study. PLoS ONE 2013, 8, e55421. [Google Scholar] [CrossRef] [PubMed]
- Deedwania, P.C.; Gupta, R.; Sharma, K.K.; Achari, V.; Gupta, B.; Maheshwari, A.; Gupta, A. High prevalence of metabolic syndrome among urban subjects in India: A multisite study. Diabetes Metab. Syndr. 2014, 8, 156–161. [Google Scholar] [CrossRef]
- Naidu, S.; Ponnampalvanar, S.; Kamaruzzaman, S.B.; Kamarulzaman, A. Prevalence of Metabolic Syndrome Among People Living with HIV in Developing Countries: A Systematic Review. AIDS Patient Care STDs 2017, 31, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Peltenburg, N.C.; Schoeman, J.C.; Hou, J.; Mora, F.; Harms, A.C.; Lowe, S.H.; Bierau, J.; Bakker, J.A.; Verbon, A.; Hankemeier, T.; et al. Persistent metabolic changes in HIV-infected patients during the first year of combination antiretroviral therapy. Sci. Rep. 2018, 8, 16947. [Google Scholar] [CrossRef]
- Cassol, E.; Misra, V.; Dutta, A.; Morgello, S.; Gabuzda, D. Cerebrospinal fluid metabolomics reveals altered waste clearance and accelerated aging in HIV patients with neurocognitive impairment. Aids 2014, 28, 1579–1591. [Google Scholar] [CrossRef]
- Cassol, E.; Misra, V.; Holman, A.; Kamat, A.; Morgello, S.; Gabuzda, D. Plasma metabolomics identifies lipid abnormalities linked to markers of inflammation, microbial translocation, and hepatic function in HIV patients receiving protease inhibitors. BMC Infect. Dis. 2013, 13, 203. [Google Scholar] [CrossRef] [PubMed]
- Rosado-Sanchez, I.; Rodriguez-Gallego, E.; Peraire, J.; Vilades, C.; Herrero, P.; Fanjul, F.; Gutierrez, F.; Bernal, E.; Pelazas, R.; Leal, M.; et al. Glutaminolysis and lipoproteins are key factors in late immune recovery in successfully treated HIV-infected patients. Clin. Sci. 2019, 133, 997–1010. [Google Scholar] [CrossRef]
- Babu, H.; Ambikan, A.T.; Gabriel, E.E.; Akusaravi, S.S.; Palaniapan, A.N.; Sundaraj, V.; Muppani, N.R.; Sperk, M.; Cheedarla, N.; Sridhar, R.; et al. Systemic inflammation and the increased risk of inflamm-aging and age-associated diseases in people living with HIV on long term suppressive antiretroviral therapy. Front. Immunol. 2019. [Google Scholar] [CrossRef] [PubMed]
- Koczor, C.A.; Jiao, Z.; Fields, E.; Russ, R.; Ludaway, T.; Lewis, W. AZT-induced mitochondrial toxicity: An epigenetic paradigm for dysregulation of gene expression through mitochondrial oxidative stress. Physiol. Genom. 2015, 47, 447–454. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Ivanov, A.V.; Valuev-Elliston, V.T.; Ivanova, O.N.; Kochetkov, S.N.; Starodubova, E.S.; Bartosch, B.; Isaguliants, M.G. Oxidative Stress during HIV Infection: Mechanisms and Consequences. Oxid. Med. Cell. Longev. 2016, 2016, 8910396. [Google Scholar] [CrossRef] [PubMed]
- Haughey, N.J.; Cutler, R.G.; Tamara, A.; McArthur, J.C.; Vargas, D.L.; Pardo, C.A.; Turchan, J.; Nath, A.; Mattson, M.P. Perturbation of sphingolipid metabolism and ceramide production in HIV-dementia. Ann. Neurol. 2004, 55, 257–267. [Google Scholar] [CrossRef]
- Pacher, P.; Beckman, J.S.; Liaudet, L. Nitric oxide and peroxynitrite in health and disease. Physiol. Rev. 2007, 87, 315–424. [Google Scholar] [CrossRef] [PubMed]
- Halliwell, B. Role of free radicals in the neurodegenerative diseases: Therapeutic implications for antioxidant treatment. Drugs Aging 2001, 18, 685–716. [Google Scholar] [CrossRef]
- Pizzino, G.; Irrera, N.; Cucinotta, M.; Pallio, G.; Mannino, F.; Arcoraci, V.; Squadrito, F.; Altavilla, D.; Bitto, A. Oxidative Stress: Harms and Benefits for Human Health. Oxid. Med. Cell. Longev. 2017, 2017, 8416763. [Google Scholar] [CrossRef]
- Wenzel, E.D.; Avdoshina, V.; Mocchetti, I. HIV-associated neurodegeneration: Exploitation of the neuronal cytoskeleton. J. Neurovirol. 2019. [Google Scholar] [CrossRef] [PubMed]
- Pace, B.T.; Lackner, A.A.; Porter, E.; Pahar, B. The Role of Defensins in HIV Pathogenesis. Mediat. Inflamm. 2017, 2017, 5186904. [Google Scholar] [CrossRef] [PubMed]
- Colomer-Lluch, M.; Ruiz, A.; Moris, A.; Prado, J.G. Restriction Factors: From Intrinsic Viral Restriction to Shaping Cellular Immunity Against HIV-1. Front. Immunol. 2018, 9, 2876. [Google Scholar] [CrossRef]
- Neil, S.J.; Zang, T.; Bieniasz, P.D. Tetherin inhibits retrovirus release and is antagonized by HIV-1 Vpu. Nature 2008, 451, 425–430. [Google Scholar] [CrossRef] [PubMed]
- Chiang, S.F.; Kan, C.Y.; Hsiao, Y.C.; Tang, R.; Hsieh, L.L.; Chiang, J.M.; Tsai, W.S.; Yeh, C.Y.; Hsieh, P.S.; Liang, Y.; et al. Bone Marrow Stromal Antigen 2 Is a Novel Plasma Biomarker and Prognosticator for Colorectal Carcinoma: A Secretome-Based Verification Study. Dis. Markers 2015, 2015, 874054. [Google Scholar] [CrossRef] [PubMed]
- Neuhaus, J.; Jacobs, D.R., Jr.; Baker, J.V.; Calmy, A.; Duprez, D.; La Rosa, A.; Kuller, L.H.; Pett, S.L.; Ristola, M.; Ross, M.J.; et al. Markers of inflammation, coagulation, and renal function are elevated in adults with HIV infection. J. Infect. Dis. 2010, 201, 1788–1795. [Google Scholar] [CrossRef]
- Van Hall, G.; Steensberg, A.; Fischer, C.; Keller, C.; Moller, K.; Moseley, P.; Pedersen, B.K. Interleukin-6 markedly decreases skeletal muscle protein turnover and increases nonmuscle amino acid utilization in healthy individuals. J. Clin. Endocrinol. Metab. 2008, 93, 2851–2858. [Google Scholar] [CrossRef] [PubMed]
- Van Hall, G. Cytokines: Muscle protein and amino acid metabolism. Curr. Opin. Clin. Nutr. Metab. Care 2012, 15, 85–91. [Google Scholar] [CrossRef]
- Almutairi, S.M.; Ali, A.K.; He, W.; Yang, D.S.; Ghorbani, P.; Wang, L.; Fullerton, M.D.; Lee, S.H. Interleukin-18 up-regulates amino acid transporters and facilitates amino acid-induced mTORC1 activation in natural killer cells. J. Biol. Chem. 2019, 294, 4644–4655. [Google Scholar] [CrossRef]
- Gomez-Munoz, A.; Gangoiti, P.; Arana, L.; Ouro, A.; Rivera, I.G.; Ordonez, M.; Trueba, M. New insights on the role of ceramide 1-phosphate in inflammation. Biochim. Biophys. Acta 2013, 1831, 1060–1066. [Google Scholar] [CrossRef]
- Hait, N.C.; Maiti, A. The Role of Sphingosine-1-Phosphate and Ceramide-1-Phosphate in Inflammation and Cancer. Mediat. Inflamm. 2017, 2017, 4806541. [Google Scholar] [CrossRef] [PubMed]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software environment for integrated models of biomolecular interaction networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | HIV-1 Negative Healthy Control | PLHIV on Therapy | p-Values |
|---|---|---|---|
| Number | 22 | 22 | |
| Age in years, mean (95% CI) | 45 (43–47) | 45 (43–48) | 0.91 |
| Sex, Female, n (%) | 10 (45%) | 9 (41%) | 0.99 |
| Telomere length, Mean (95% CI) 1 | 5.9 (4.3–7.2) | 2.3 (1.6–2.9) | <0.001 |
| Nadir CD4 count, median (IQR)) | - | 229 (176–318) | - |
| CD4 count at sampling, median (IQR) | - | 624 (524–746) | - |
| CD8 count at sampling, median (IQR) | - | 757 (665–882) | - |
| Viral load, copies/mL | - | <150 | - |
| CD4:CD8 ratio, median (IQR) | - | 0.8 (0.58–1.02) | - |
| Years on therapy, median (IQR) | - | 9 (6–10) | - |
| Reservoir, log10 copies/106 cells, median (IQR) | - | 2.78 (2.58–3.01) | - |
| Treatment regimen AZT/3TC/NVP TDF/3TC/EFV | - | 13 (59%) 9 (41%) | - |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Babu, H.; Sperk, M.; Ambikan, A.T.; Rachel, G.; Viswanathan, V.K.; Tripathy, S.P.; Nowak, P.; Hanna, L.E.; Neogi, U. Plasma Metabolic Signature and Abnormalities in HIV-Infected Individuals on Long-Term Successful Antiretroviral Therapy. Metabolites 2019, 9, 210. https://doi.org/10.3390/metabo9100210
Babu H, Sperk M, Ambikan AT, Rachel G, Viswanathan VK, Tripathy SP, Nowak P, Hanna LE, Neogi U. Plasma Metabolic Signature and Abnormalities in HIV-Infected Individuals on Long-Term Successful Antiretroviral Therapy. Metabolites. 2019; 9(10):210. https://doi.org/10.3390/metabo9100210
Chicago/Turabian StyleBabu, Hemalatha, Maike Sperk, Anoop T. Ambikan, Gladys Rachel, Vinod Kumar Viswanathan, Srikanth P. Tripathy, Piotr Nowak, Luke Elizabeth Hanna, and Ujjwal Neogi. 2019. "Plasma Metabolic Signature and Abnormalities in HIV-Infected Individuals on Long-Term Successful Antiretroviral Therapy" Metabolites 9, no. 10: 210. https://doi.org/10.3390/metabo9100210
APA StyleBabu, H., Sperk, M., Ambikan, A. T., Rachel, G., Viswanathan, V. K., Tripathy, S. P., Nowak, P., Hanna, L. E., & Neogi, U. (2019). Plasma Metabolic Signature and Abnormalities in HIV-Infected Individuals on Long-Term Successful Antiretroviral Therapy. Metabolites, 9(10), 210. https://doi.org/10.3390/metabo9100210