Tear Film Amphiphilic and Anti-Inflammatory Lipids in Bovine Pink Eye

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
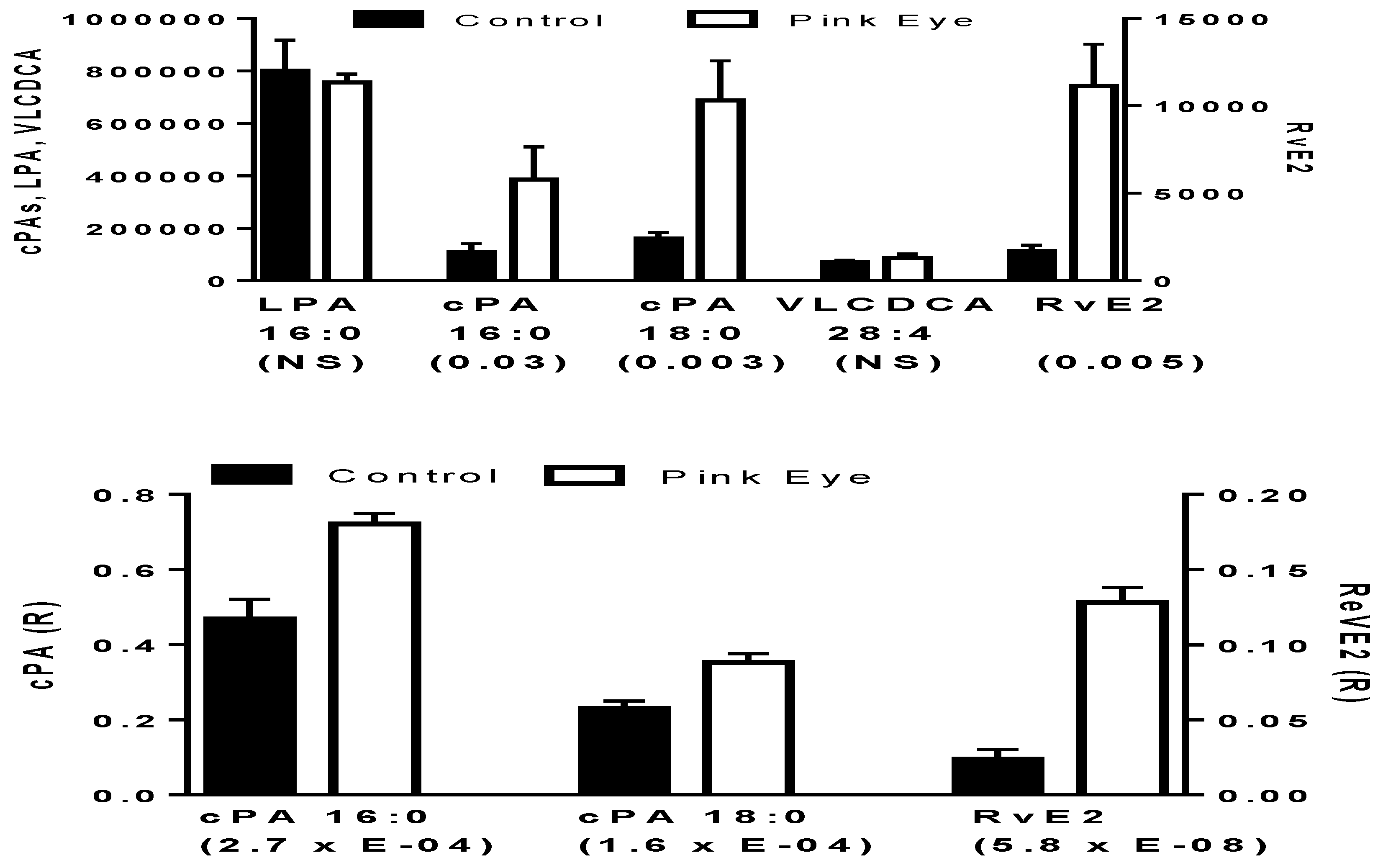
2.1. Anti-Inflammatory Lipids
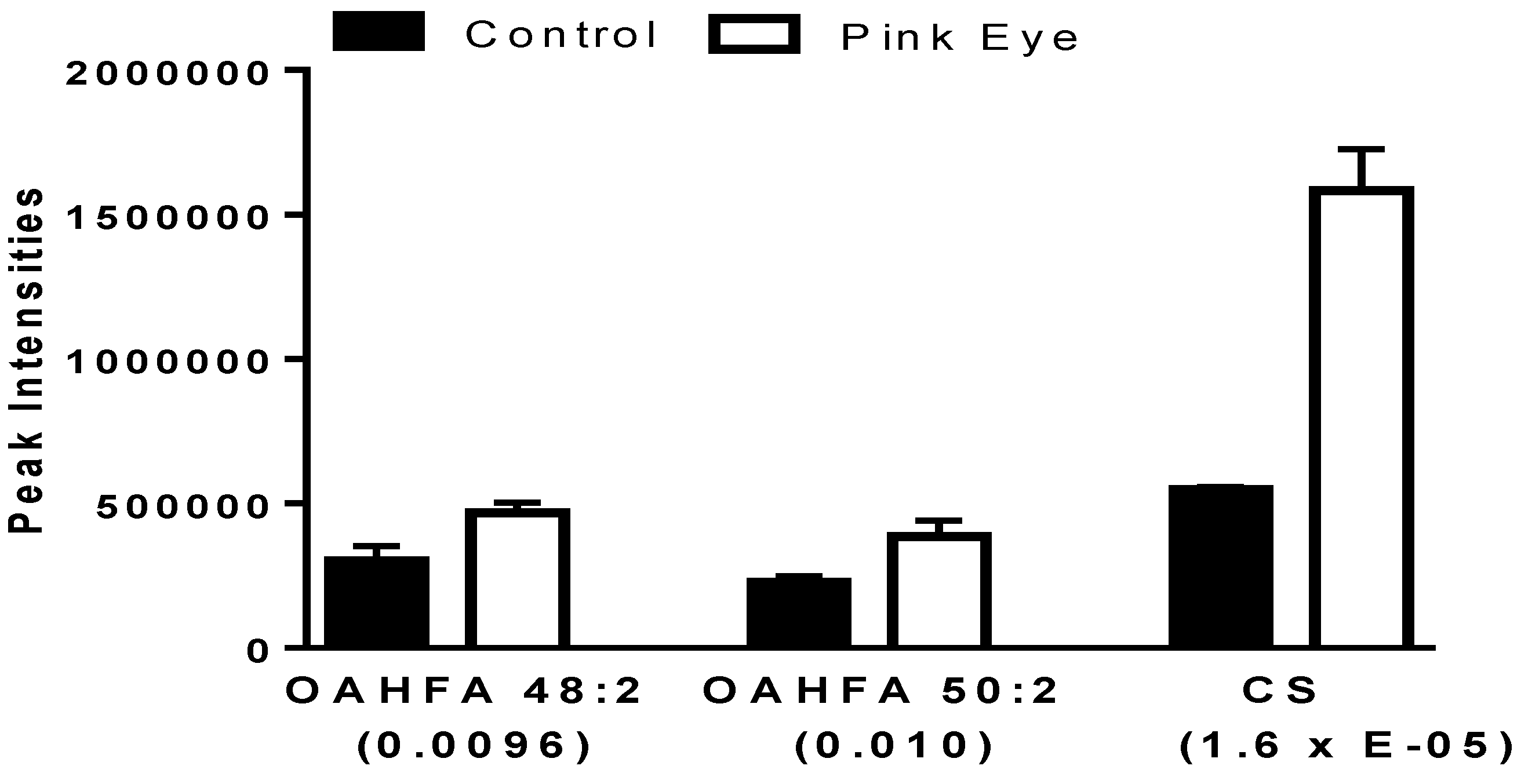
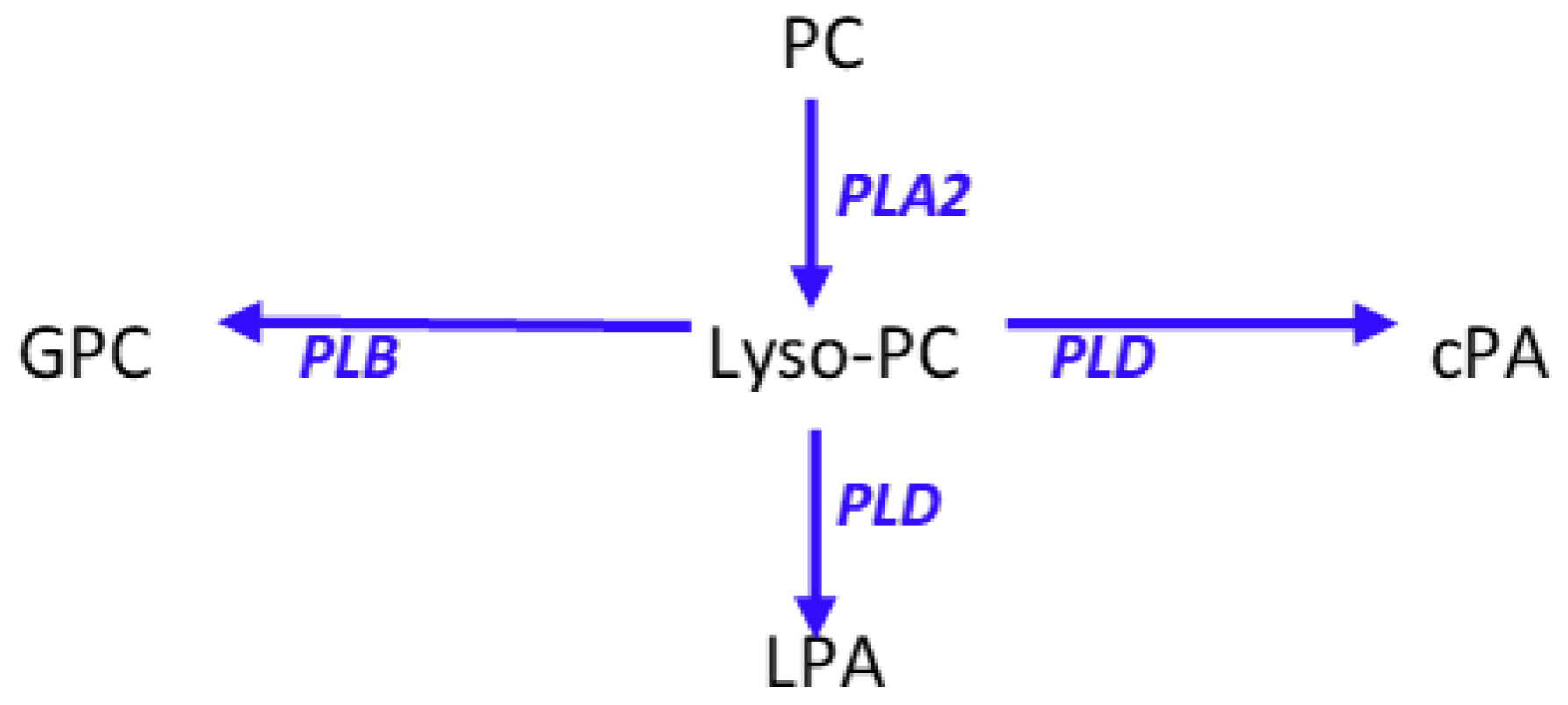
2.2. Amphiphilic Lipids.
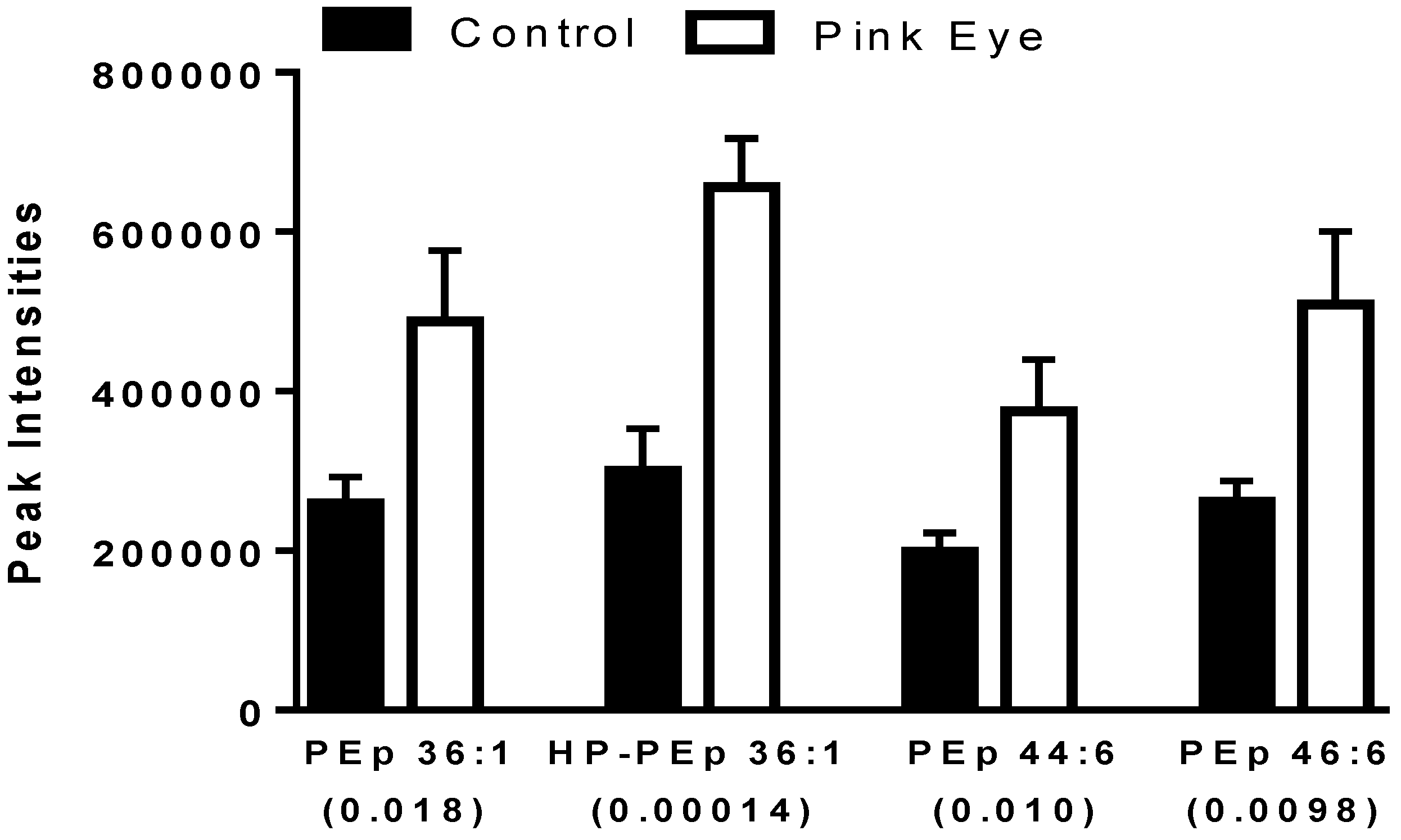
2.3. Structural Glycerophospholipids
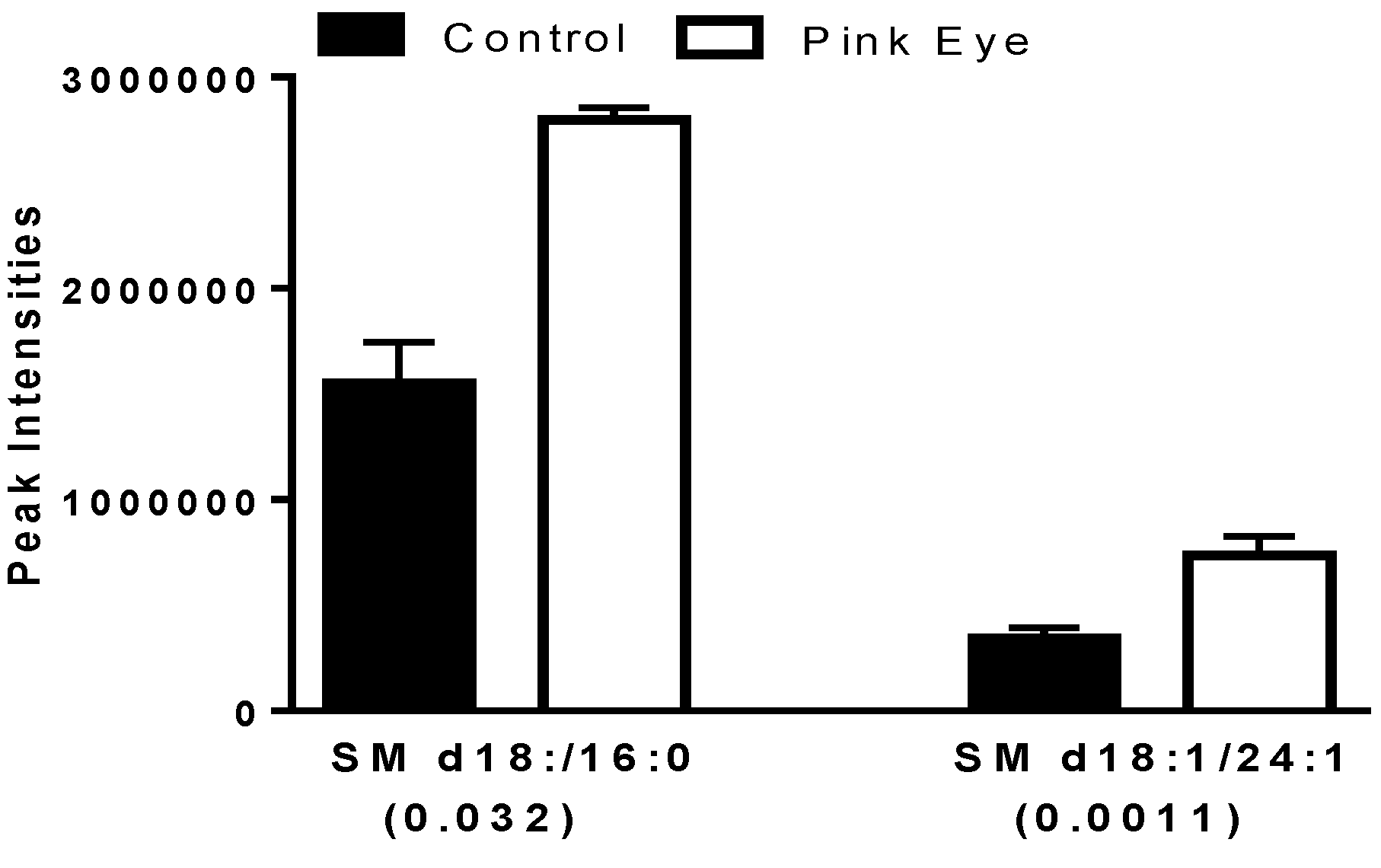
2.4. Structural Sphingolipids
3. Discussion
4. Materials and Methods
4.1. Clinical Samples
4.2. Lipidomics
4.3. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Kasikara, C.; Doran, A.C.; Cai, B.; Tabas, I. The role of non-resolving inflammation in atherosclerosis. J. Clin. Investig. 2018, 128, 2713–2723. [Google Scholar] [CrossRef] [PubMed]
- Falconer, J.; Murphy, A.N.; Young, S.P.; Clark, A.R.; Tiziani, S.; Guma, M.; Buckley, C.D. Review: Synovial Cell Metabolism and Chronic Inflammation in Rheumatoid Arthritis. Arthritis Rheumatol. 2018, 70, 984–999. [Google Scholar] [CrossRef] [PubMed]
- Reyes, J.L.; Vannan, D.T.; Eksteen, B.; Avelar, I.J.; Rodríguez, T.; González, M.I.; Mendoza, A.V. Innate and Adaptive Cell Populations Driving Inflammation in Dry Eye Disease. Mediat. Inflamm. 2018, 2018, 2532314. [Google Scholar] [CrossRef] [PubMed]
- Tjonahen, E.; Oh, S.F.; Siegelman, J.; Elangovan, S.; Percarpio, K.B.; Hong, S.; Arita, M.; Serhan, C.N. Resolvin E2: Identification and anti-inflammatory actions: Pivotal role of human 5-lipoxygenase in resolvin E series biosynthesis. Chem. Biol. 2006, 13, 1193–1202. [Google Scholar] [CrossRef] [PubMed]
- Serhan, C.N.; Levy, B.D. Resolvins in inflammation: Emergence of the pro-resolving superfamily of mediators. J. Clin. Investig. 2018, 128, 2657–2669. [Google Scholar] [CrossRef] [PubMed]
- Gotoh, M.; Nagano, A.; Tsukahara, R.; Murofushi, H.; Morohoshi, T.; Otsuka, K.; Murakami-Murofushi, K. Cyclic phosphatidic acid relieves osteoarthritis symptoms. Mol. Pain. 2014, 10, 52. [Google Scholar] [CrossRef] [PubMed]
- Hashimoto, K.; Nakashima, M.; Hamano, A.; Gotoh, M.; Ikeshima-Kataoka, H.; Murakami-Murofushi, K.; Miyamoto, Y. 2-carba cyclic phosphatidic acid suppresses inflammation via regulation of microglial polarisation in the stab-wounded mouse cerebral cortex. Sci. Rep. 2018, 8, 9715. [Google Scholar] [CrossRef] [PubMed]
- Wood, P.L. Endogenous Anti-Inflammatory Very-Long-Chain Dicarboxylic Acids: Potential Chemopreventive Lipids. Metabolites 2018, 8, 76. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.E.; Sun, Y.; Gjorstrup, P.; Pearlman, E. Inhibition of Corneal Inflammation by the Resolvin E1. Investig. Ophthalmol. Vis. Sci. 2015, 56, 2728–2736. [Google Scholar] [CrossRef] [PubMed]
- Oh, S.F.; Dona, M.; Fredman, G.; Krishnamoorthy, S.; Irimia, D.; Serhan, C.N. Resolvin E2 formation and impact in inflammation resolution. J. Immunol. 2012, 188, 4527–4534. [Google Scholar] [CrossRef] [PubMed]
- Ritchie, S.A.; Jayasinghe, D.; Davies, G.F.; Ahiahonu, P.; Ma, H.; Goodenowe, D.B. Human serum-derived hydroxy long-chain fatty acids exhibit anti-inflammatory and anti-proliferative activity. J. Exp. Clin. Cancer Res. 2011, 30, 59. [Google Scholar] [CrossRef] [PubMed]
- Butovich, I.A. Tear film lipids. Exp. Eye Res. 2013, 117, 4–27. [Google Scholar] [CrossRef] [PubMed]
- Lam, S.M.; Tong, L.; Yong, S.S.; Li, B.; Chaurasia, S.S.; Shui, G.; Wenk, M.R. Meibum lipid composition in Asians with dry eye disease. PLoS ONE 2011, 6, e24339. [Google Scholar] [CrossRef] [PubMed]
- Fujiwara, Y. Cyclic phosphatidic acid—A unique bioactive phospholipid. Biochim. Biophys. Acta 2008, 1781, 519–524. [Google Scholar] [CrossRef] [PubMed]
- Wood, P.L.; Scoggin, K.; Ball, B.A.; Troedsson, M.H.; Squires, E.L. Lipidomics of equine sperm and seminal plasma: Identification of amphiphilic (O-acyl)-ω-hydroxy-fatty acids. Theriogenology 2016, 86, 1212–1221. [Google Scholar] [CrossRef] [PubMed]
- Wood, P.L.; Ball, B.A.; Scoggin, K.; Troedsson, M.H.; Squires, E.L. Lipidomics of equine amniotic fluid: Identification of amphiphilic (O-acyl)-ω-hydroxy-fatty acids. Theriogenology 2018, 105, 120–125. [Google Scholar] [CrossRef] [PubMed]
- Wood, P.L.; Steinman, M.; Erol, E.; Carter, C.; Christmann, U.; Verma, A. Lipidomic analysis of immune activation in equine leptospirosis and Leptospira-vaccinated horses. PLoS ONE 2018, 13, e0193424. [Google Scholar] [CrossRef] [PubMed]
- Mori, N.; Fukano, Y.; Arita, R.; Shirakawa, R.; Kawazu, K.; Nakamura, M.; Amano, S. Rapid identification of fatty acids and (O-acyl)-ω-hydroxy fatty acids in human meibum by liquid chromatography/high-resolution mass spectrometry. J. Chromatogr. A 2014, 1347, 129–136. [Google Scholar] [CrossRef] [PubMed]
- Lam, S.M.; Tong, L.; Duan, X.; Petznick, A.; Wenk, M.R.; Shui, G. Extensive characterization of human tear fluid collected using different techniques unravels the presence of novel lipid amphiphiles. J. Lipid Res. 2015, 55, 289–298. [Google Scholar] [CrossRef] [PubMed]
- Brown, S.H.; Kunnen, C.M.; Duchoslav, E.; Dolla, N.K.; Kelso, M.J.; Papas, E.B.; Lazon de la Jara, P.; Willcox, M.D.; Blanksby, S.J.; Mitchell, T.W. A comparison of patient matched meibum and tear lipidomes. Investig. Ophthalmol. Vis. Sci. 2013, 54, 7417–7424. [Google Scholar] [CrossRef] [PubMed]
- Butovich, I.A. On the presence of (O-acyl)-omega-hydroxy fatty acids and of their esters in human meibomian gland secretions. Investig. Ophthalmol. Vis. Sci. 2011, 52, 639–641. [Google Scholar] [CrossRef] [PubMed]
- McMahon, A.; Lu, H.; Butovich, I.A. A role for ELOVL4 in the mouse meibomian gland and sebocyte cell biology. Investig. Ophthalmol. Vis. Sci. 2014, 55, 2832–2840. [Google Scholar] [CrossRef] [PubMed]
- Kalužíková, A.; Vrkoslav, V.; Harazim, E.; Hoskovec, M.; Plavka, R.; Buděšínský, M.; Bosáková, Z.; Cvačka, J. Cholesteryl esters of ω-(O-acyl)-hydroxy fatty acids in vernix caseosa. J. Lipid Res. 2017, 58, 1579–1590. [Google Scholar] [CrossRef] [PubMed]
- Johnson, A.L.; Edson, K.Z.; Totah, R.A.; Rettie, A.E. Cytochrome P450 ω-Hydroxylases in Inflammation and Cancer. Adv. Pharmacol. 2015, 74, 223–262. [Google Scholar] [PubMed]
- Alexanian, A.; Sorokin, A. Targeting 20-HETE producing enzymes in cancer—Rationale, pharmacology, and clinical potential. Onco Targets Ther. 2013, 6, 243–255. [Google Scholar] [PubMed]
- English, J.T.; Norris, P.C.; Hodges, R.R.; Dartt, D.A.; Serhan, C.N. Identification and Profiling of Specialized Pro-Resolving Mediators in Human Tears by Lipid Mediator Metabolomics. Prostag. Leuk. Essent. Fat. Acids 2017, 117, 17–27. [Google Scholar] [CrossRef] [PubMed]
- Cholkar, K.; Gilger, B.C.; Mitra, A.K. Topical delivery of aqueous micellar resolvin E1 analog (RX-10045). Int. J. Pharm. 2016, 498, 326–334. [Google Scholar] [CrossRef] [PubMed]
- Frank, S.K.; Gerber, J.D. Hydrolytic enzymes of Moraxella bovis. J. Clin. Microbiol. 1981, 13, 269–271. [Google Scholar] [PubMed]
- Jiang, F.; Huang, S.; Imadad, K.; Li, C. Cloning and expression of a gene with phospholipase B activity from Pseudomonas fluorescens in Escherichia coli. Bioresour. Technol. 2012, 104, 518–522. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.; Grice, I.D.; Peak, I.R.; Frost, T.; Yue, G.; Wilson, J.C. The role of lipooligosaccharide in the biological activity of Moraxella bovis strains Epp63, Mb25 and L183/2, and isolation of capsular polysaccharide from L183/2. Carbohydr. Res. 2018, 467, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Ely, V.L.; Vargas, A.C.; Costa, M.M.; Oliveira, H.P.; Pötter, L.; Reghelin, M.A.; Fernandes, A.W.; Pereira, D.I.B.; Sangioni, L.A.; Botton, S.A. Moraxella bovis, Moraxella ovis and Moraxella bovoculi: Biofilm formation and lysozyme activity. J. Appl. Microbiol. 2018. [Google Scholar] [CrossRef] [PubMed]
- Wood, P.L. Non-Targeted Lipidomics Utilizing Constant Infusion High Resolution ESI Mass Spectrometry. In Lipidomics; Wood, P.L., Ed.; Neuromethods; Humana Press: New York, NY, 2017; Volume 125, pp. 13–19. [Google Scholar]





© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wood, P.L.; Donohue, M.N.; Cebak, J.E.; Beckmann, T.G.; Treece, M.; Johnson, J.W.; Miller, L.M.J. Tear Film Amphiphilic and Anti-Inflammatory Lipids in Bovine Pink Eye. Metabolites 2018, 8, 81. https://doi.org/10.3390/metabo8040081
Wood PL, Donohue MN, Cebak JE, Beckmann TG, Treece M, Johnson JW, Miller LMJ. Tear Film Amphiphilic and Anti-Inflammatory Lipids in Bovine Pink Eye. Metabolites. 2018; 8(4):81. https://doi.org/10.3390/metabo8040081
Chicago/Turabian StyleWood, Paul L., Michelle N. Donohue, John E. Cebak, Taylor G. Beckmann, MacKenzie Treece, Jason W. Johnson, and Lynda M. J. Miller. 2018. "Tear Film Amphiphilic and Anti-Inflammatory Lipids in Bovine Pink Eye" Metabolites 8, no. 4: 81. https://doi.org/10.3390/metabo8040081
APA StyleWood, P. L., Donohue, M. N., Cebak, J. E., Beckmann, T. G., Treece, M., Johnson, J. W., & Miller, L. M. J. (2018). Tear Film Amphiphilic and Anti-Inflammatory Lipids in Bovine Pink Eye. Metabolites, 8(4), 81. https://doi.org/10.3390/metabo8040081