Endogenous Anti-Inflammatory Very-Long-Chain Dicarboxylic Acids: Potential Chemopreventive Lipids †

Abstract
1. Introduction
2. Results
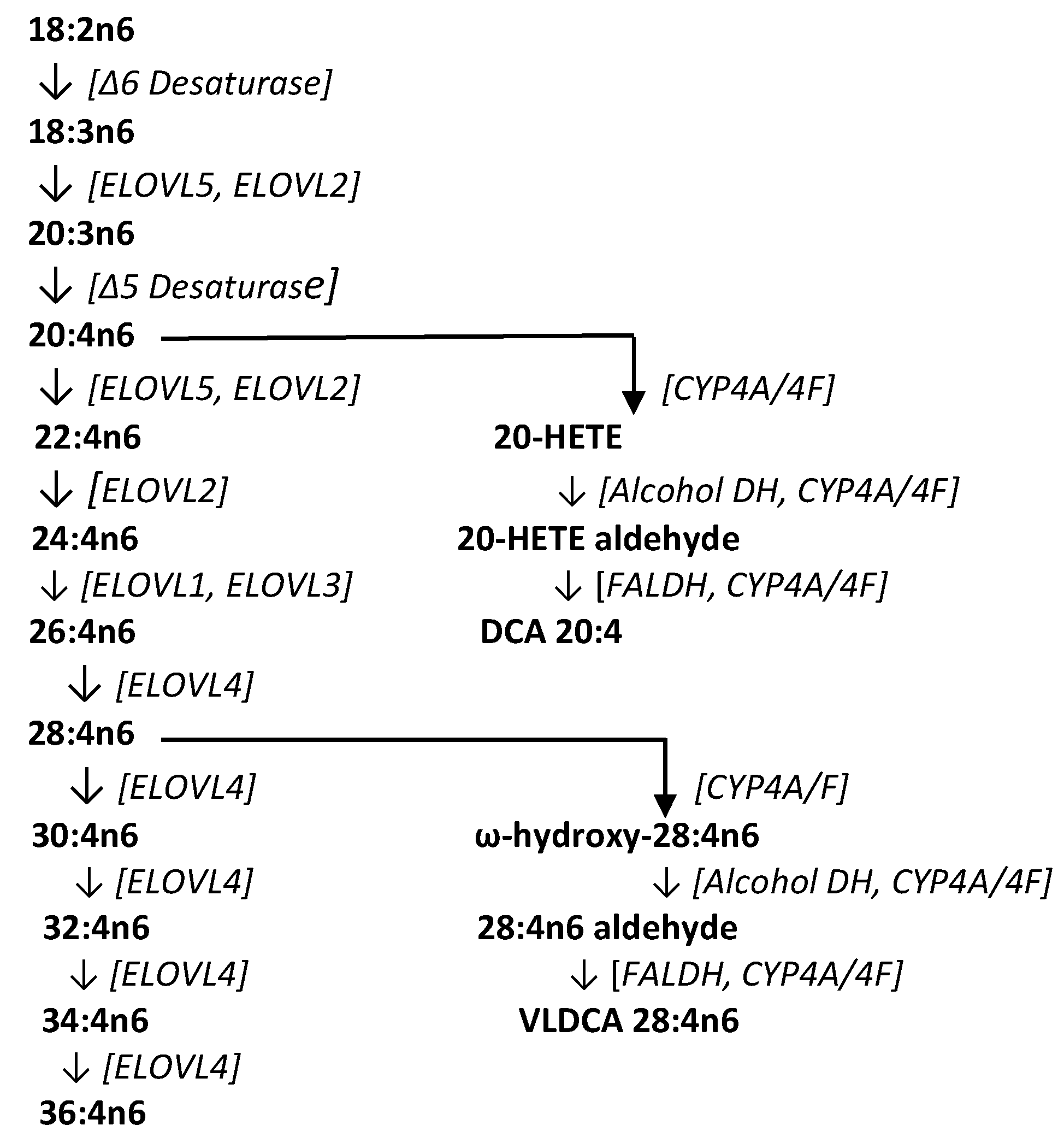
2.1. HR-MS Analyses of C28H46O4
2.2. Structural Validation of VLCDCA 28:4
2.3. VLCDCA 28:4 in Biofluids
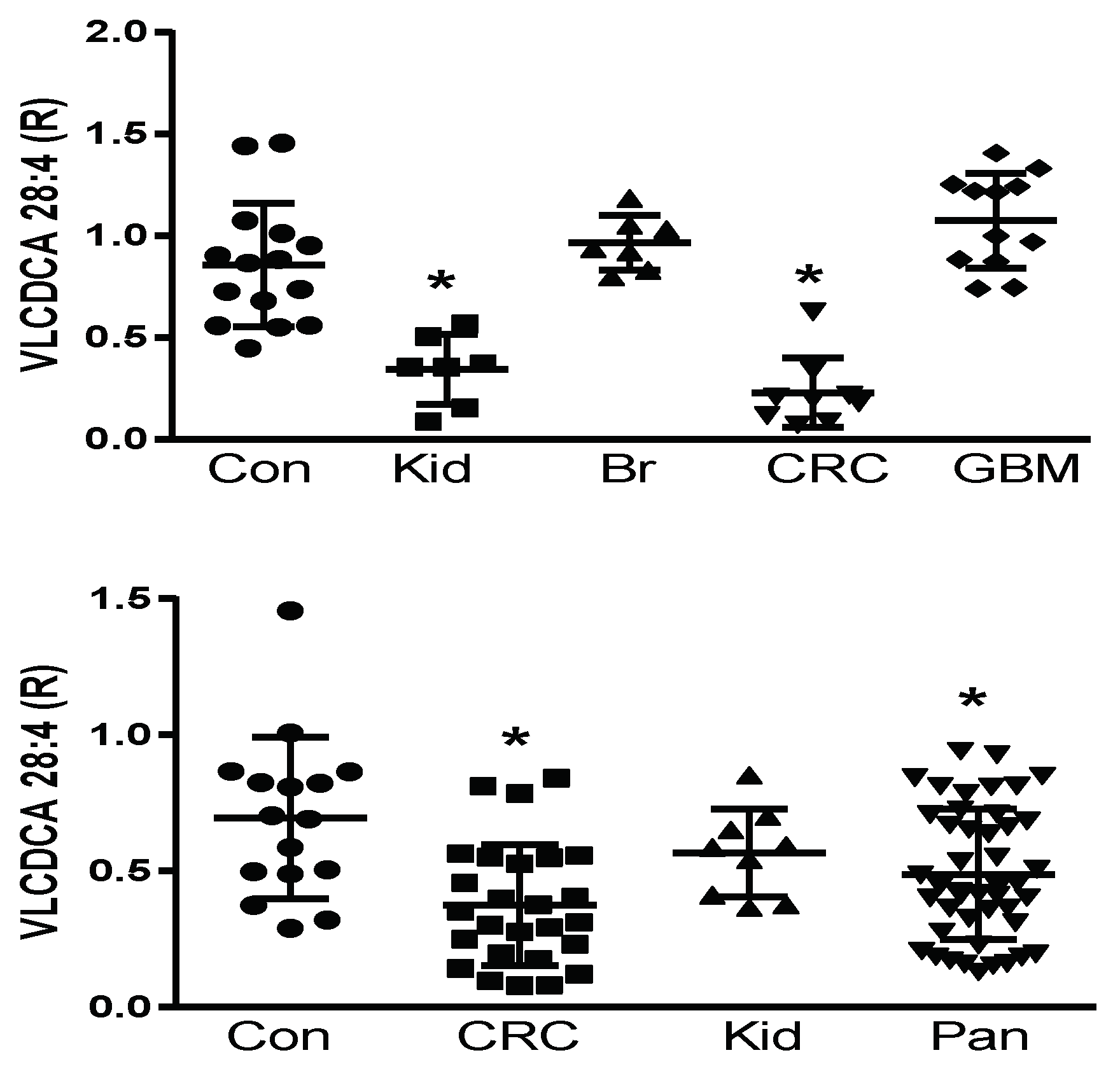
2.4. Plasma VLCDCA 28:4 in Cancer Patients
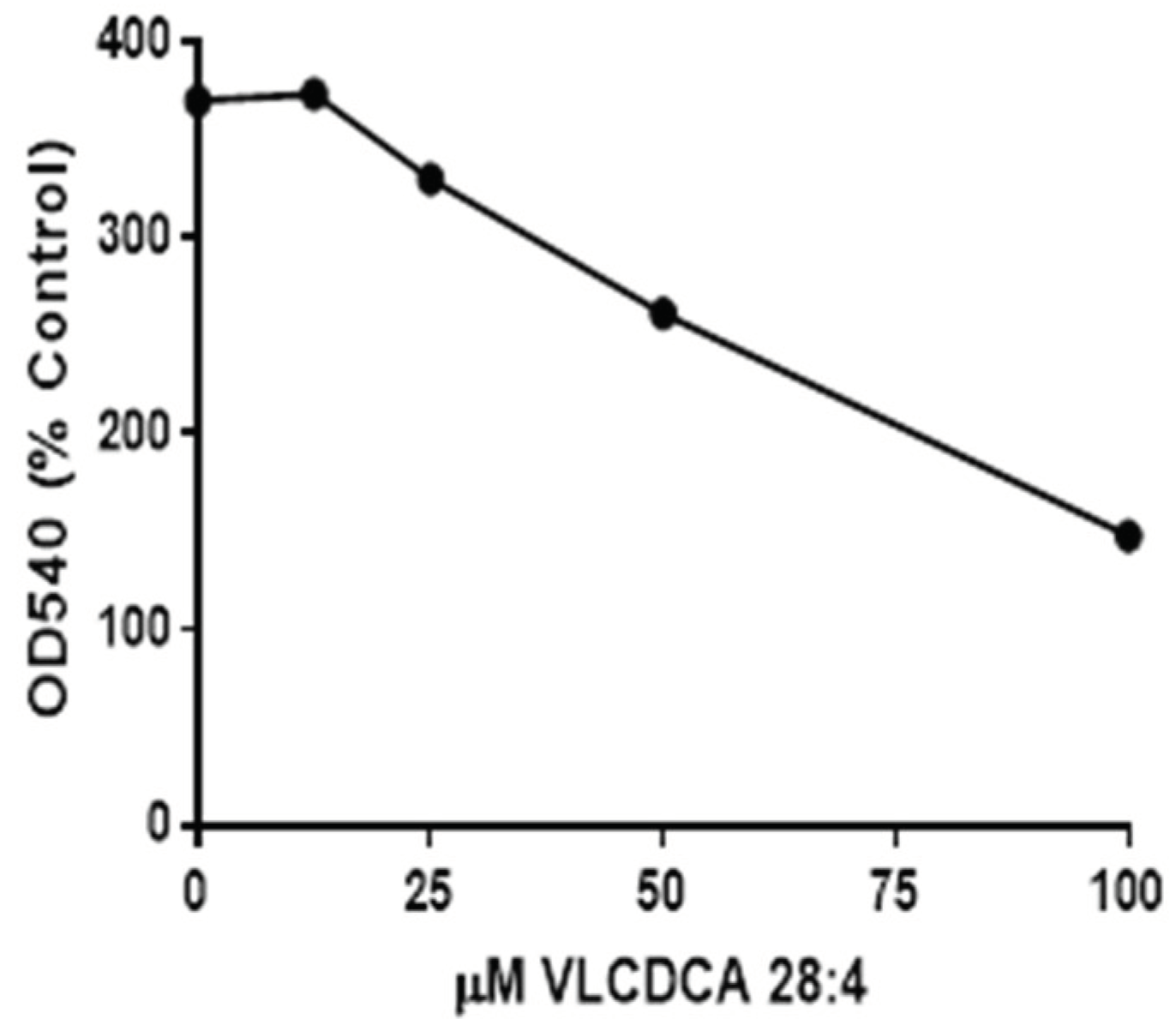
2.5. VLCDCA 28:4: Anti-Inflammatory Activity
3. Discussion
4. Materials and Methods
4.1. Clinical Samples
4.2. Sample Processing
4.3. Semipurification of Plasma VLCDCAs
4.4. Chemical Synthesis of VLCDCA 28:4

4.5. High-Resolution Mass Spectrometric Analyses
4.6. Derivatization: Picolylamine
4.7. Anti-Inflammatory Assay
5. Conclusions
6. Patents
Supplementary Materials
Funding
Acknowledgments
Conflicts of Interest
References
- Amiel, G.; Hoffman, A.; Half, E.E. Update on Screening for Urological Malignancies. Rambam Maimonides Med. J. 2017, 8, e0041. [Google Scholar]
- Diaz de Leon, A.; Pedrosa, I. Imaging and Screening of Kidney Cancer. Radiol. Clin. North Am. 2017, 55, 1235–1250. [Google Scholar] [CrossRef] [PubMed]
- Youn, S.Y.; Rha, S.E.; Jung, E.S.; Lee, I.S. Pancreas ductal adenocarcinoma with cystic features on cross-sectional imaging: Radiologic-pathologic correlation. Diagn. Interv. Radiol. 2018, 24, 5–11. [Google Scholar] [CrossRef] [PubMed]
- Horvat, N.; Ryan, D.E.; LaGratta, M.D.; Shah, P.M.; Do, R.K. Imaging for pancreatic ductal adenocarcinoma. Chin. Clin. Oncol. 2017, 6, 62. [Google Scholar] [CrossRef] [PubMed]
- Aziz, M.H.; Sideras, K.; Aziz, N.A.; Mauff, K.; Haen, R.; Roos, D.; Saida, L.; Suker, M.; van der Harst, E.; Mieog, J.S.; et al. The Systemic-Immune-Inflammation Index Independently Predicts Survival and Recurrence in Resectable Pancreatic Cancer and its Prognostic Value Depends on Bilirubin Levels: A Retrospective Multicenter Cohort Study. Ann. Surg. 2018. [Google Scholar] [CrossRef] [PubMed]
- Krah, N.M.; Murtaugh, L.C. Differentiation and Inflammation: ‘Best Enemies’ in Gastrointestinal Carcinogenesis. Trends Cancer 2016, 2, 723–735. [Google Scholar] [CrossRef] [PubMed]
- Khalafalla, F.G.; Khan, M.W. Inflammation and Epithelial-Mesenchymal Transition in Pancreatic Ductal Adenocarcinoma: Fighting Against Multiple Opponents. Cancer Growth Metastasis 2017, 10, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Aoki, T.; Narumiya, S. Prostaglandin E2-EP2 signaling as a node of chronic inflammation in the colon tumor microenvironment. Inflamm. Regen. 2017, 37, 4. [Google Scholar] [CrossRef] [PubMed]
- Long, A.G.; Lundsmith, E.T.; Hamilton, K.E. Inflammation and Colorectal Cancer. Curr. Colorectal Cancer Rep. 2017, 13, 341–351. [Google Scholar] [CrossRef] [PubMed]
- Kooman, J.P.; Dekker, M.J.; Usvyat, L.A.; Kotanko, P.; van der Sande, F.M.; Schalkwijk, C.G.; Shiels, P.G.; Stenvinkel, P. Inflammation and premature aging in advanced chronic kidney disease. Am. J. Physiol. Renal. Physiol. 2017, 313, F938–F950. [Google Scholar] [CrossRef] [PubMed]
- Komaki, Y.; Komaki, F.; Micic, D.; Ido, A.; Sakuraba, A. Risk of Colorectal Cancer in Chronic Kidney Disease: A Systematic Review and Meta-Analysis. J. Clin. Gastroenterol. 2017, 52, 796–804. [Google Scholar] [CrossRef] [PubMed]
- Ricciardiello, L.; Ahnen, D.J.; Lynch, P.M. Chemoprevention of hereditary colon cancers: Time for new strategies. Nat. Rev. Gastroenterol. Hepatol. 2016, 13, 352–361. [Google Scholar] [CrossRef] [PubMed]
- Gong, J.; Sachdev, E.; Robbins, L.A.; Lin, E.; Hendifar, A.E.; Mita, M.M. Statins and pancreatic cancer. Oncol. Lett. 2017, 13, 1035–1040. [Google Scholar] [CrossRef] [PubMed]
- Nabavi, S.F.; Bilotto, S.; Russo, G.L.; Orhan, I.E.; Habtemariam, S.; Daglia, M.; Devi, K.P.; Loizzo, M.R.; Tundis, R.; Nabavi, S.M. Omega-3 polyunsaturated fatty acids and cancer: Lessons learned from clinical trials. Cancer Metastasis Rev. 2015, 34, 359–380. [Google Scholar] [CrossRef] [PubMed]
- Arakaki, A.K.; Mezencev, R.; Bowen, N.J.; Huang, Y.; McDonald, J.F.; Skolnick, J. Identification of metabolites with anticancer properties by computational metabolomics. Mol. Cancer 2008, 7, 57. [Google Scholar] [CrossRef] [PubMed]
- Parolin, C.; Calonghi, N.; Presta, E.; Boga, C.; Caruana, P.; Naldi, M.; Andrisano, V.; Masotti, L.; Sartor, G. Mechanism and stereoselectivity of HDAC I inhibition by (R)-9-hydroxystearic acid in coloncancer. Biochim. Biophys. Acta 2012, 1821, 1334–1340. [Google Scholar] [CrossRef] [PubMed]
- Ritchie, S.A.; Jayasinghe, D.; Davies, G.F.; Ahiahonu, P.; Ma, H.; Goodenowe, D.B. Human serum-derived hydroxy long-chain fatty acids exhibit anti-inflammatory and anti-proliferative activity. J. Exp. Clin. Cancer Res. 2011, 30, 59. [Google Scholar] [CrossRef] [PubMed]
- Ritchie, S.A.; Heath, D.; Yamazaki, Y.; Grimmalt, B.; Kavianpour, A.; Krenitsky, K.; Elshoni, H.; Takemasa, I.; Miyake, M.; Sekimoto, M.; et al. Reduction of novel circulating long-chain fatty acids in colorectal cancer patients is independent of tumor burden and correlates with age. BMC Gastroenterol. 2010, 10, 140. [Google Scholar] [CrossRef] [PubMed]
- Ritchie, S.A.; Ahiahonu, P.W.; Jayasinghe, D.; Heath, D.; Liu, J.; Lu, Y.; Jin, W.; Kavianpour, A.; Yamazaki, Y.; Khan, A.M.; et al. Reduced levels of hydroxylated, polyunsaturated ultra long-chain fatty acids in the serum of colorectal cancer patients: Implications for early screening and detection. BMC Med. 2010, 8, 13. [Google Scholar] [CrossRef] [PubMed]
- Ritchie, S.A.; Tonita, J.; Alvi, R.; Lehotay, D.; Elshoni, H.; Myat, S.; McHattie, J.; Goodenowe, D.B. Low-serum GTA-446 anti-inflammatory fatty acid levels as a new risk factor for colon cancer. Int. J. Cancer 2013, 132, 355–362. [Google Scholar] [CrossRef] [PubMed]
- Ritchie, S.A.; Akita, H.; Takemasa, I.; Eguchi, H.; Pastural, E.; Nagano, H.; Monden, M.; Doki, Y.; Mori, M.; Jin, W.; et al. Metabolic system alterations in pancreatic cancer patient serum: Potential for early detection. BMC Cancer 2013, 13, 416. [Google Scholar] [CrossRef] [PubMed]
- Ritchie, S.A.; Chitou, B.; Zheng, Q.; Jayasinghe, D.; Jin, W.; Mochizuki, A.; Goodenowe, D.B. Pancreatic cancer serum biomarker PC-594: Diagnostic performance and comparison to CA19-9. World J. Gastroenterol. 2015, 21, 6604–6612. [Google Scholar] [CrossRef] [PubMed]
- Senanayake, V.K.; Jin, W.; Mochizuki, A.; Chitou, B.; Goodenowe, D.B. Metabolic dysfunctions in multiple sclerosis: Implications as to causation, early detection, and treatment, a case control study. BMC Neurol. 2015, 15, 154. [Google Scholar] [CrossRef] [PubMed]
- Ritchie, S.A.; Goodenowe, D.B. Methods for the diagnosis of colorectal cancer and ovarian cancer health states. International Patent Application No. PCT/CA2006/001502, 10 May 2017. [Google Scholar]
- Liu, A.; Chang, J.; Lin, Y.; Shen, Z.; Bernstein, P.S. Long-chain and very long-chain polyunsaturated fatty acids in ocular aging and age-related macular degeneration. J. Lipid Res. 2010, 51, 3217–3729. [Google Scholar] [CrossRef] [PubMed]
- Poulos, A.; Sharp, P.; Johnson, D.; White, I.; Fellenberg, A. The occurrence of polyenoic fatty acids with greater than 22 carbon atoms in mammalian spermatozoa. Biochem. J. 1986, 240, 891–895. [Google Scholar] [CrossRef] [PubMed]
- Sandhoff, R. Very long chain sphingolipids: Tissue expression, function and synthesis. FEBS Lett. 2010, 584, 1907–1913. [Google Scholar] [CrossRef] [PubMed]
- Sanders, R.J.; Ofman, R.; Dacremont, G.; Wanders, R.J.; Kemp, S. Characterization of the human omega-oxidation pathway for omega-hydroxy-very-long-chain fatty acids. FASEB J. 2008, 22, 2064–2071. [Google Scholar] [CrossRef] [PubMed]
- Kundu, R.K.; Getz, G.S.; Tonsgard, J.H. Induction of (omega-1)-oxidation of monocarboxylic acids by acetylsalicylic acid. J. Lipid Res. 1993, 34, 1187–1199. [Google Scholar] [CrossRef] [PubMed]
- Johnson, A.L.; Edson, K.Z.; Totah, R.A.; Rettie, A.E. Cytochrome P450 ω-Hydroxylases in Inflammation and Cancer. Adv. Pharmacol. 2015, 74, 223–262. [Google Scholar] [PubMed]
- Hsu, M.H.; Savas, U.; Griffin, K.J.; Johnson, E.F. Human cytochrome p450 family 4 enzymes: Function, genetic variation and regulation. Drug Metab. Rev. 2007, 39, 515–538. [Google Scholar] [CrossRef] [PubMed]
- Alexanian, A.; Sorokin, A. Targeting 20-HETE producing enzymes in cancer—rationale, pharmacology, and clinical potential. Oncol. Targets Ther. 2013, 6, 243–255. [Google Scholar]
- Kumarakulasingham, M.; Rooney, P.H.; Dundas, S.R.; Telfer, C.; Melvin, W.T.; Curran, S.; Murray, G.I. Cytochrome p450 profile of colorectal cancer: Identification of markers of prognosis. Clin. Cancer Res. 2005, 11, 3758–3765. [Google Scholar] [CrossRef] [PubMed]
- Moon, J.W.; Lee, S.K.; Lee, J.O.; Kim, N.; Lee, Y.W.; Kim, S.J.; Kang, H.J.; Kim, J.; Kim, H.S.; Park, S.H. Identification of novel hypermethylated genes and demethylating effect of vincristine in colorectal cancer. J. Exp. Clin. Cancer Res. 2014, 33, 4. [Google Scholar] [CrossRef] [PubMed]
- Singh Ranger, G. The role of aspirin in colorectal cancer chemoprevention. Crit. Rev. Oncol. Hematol. 2016, 104, 87–90. [Google Scholar] [CrossRef] [PubMed]
- Höög, J.O.; Ostberg, L.J. Mammalian alcohol dehydrogenases-a comparative investigation at gene and protein levels. Chem. Biol. Interact. 2011, 191, 2–7. [Google Scholar] [CrossRef] [PubMed]
- Pors, K.; Moreb, J.S. Aldehyde dehydrogenases in cancer: An opportunity for biomarker and drug development? Drug Discov. Today 2014, 19, 1953–1963. [Google Scholar] [CrossRef] [PubMed]
- Chen, B.; Hu, K.W.; Zhang, J.W.; Wei, Z.J.; Meng, X.L.; Xiong, M.M. A critical analysis of the relationship between aldehyde dehydrogenases-2 Glu487Lys polymorphism and colorectal cancer susceptibility. Pathol. Oncol. Res. 2015, 21, 727–733. [Google Scholar] [CrossRef] [PubMed]
- Wood, P.L. Non-targeted lipidomics utilizing constant infusion high resolution ESI mass spectrometry. In Lipidomics; Wood, P.L., Ed.; Humana Press: New York, NY, USA, 2017; Volume 125, pp. 13–19. [Google Scholar]
- Wood, P.L.; Locke, V.A.; Herling, P.; Passaro, A.; Vigna, G.B.; Volpato, S.; Valacchi, G.; Cervellati, C.; Zuliani, G. Targeted lipidomics distinguishes patient subgroups in mild cognitive impairment (MCI) and late onset Alzheimer’s disease (LOAD). BBA Clin. 2015, 5, 25–28. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Xu, W.; Vadivel, S.K.; Fan, P.; Makriyannis, A. High affinity electrophilic and photoactivatable covalent endocannabinoid probes for the CB1 receptor. J. Med. Chem. 2005, 48, 6423–6429. [Google Scholar] [CrossRef] [PubMed]
- Higashi, T.; Ichikawa, T.; Inagaki, S.; Min, J.Z.; Fukushima, T.; Toyo’oka, T. Simple and practical derivatization procedure for enhanced detection of carboxylic acids in liquid chromatography-electrospray ionization-tandem mass spectrometry. Pharm. Biomed. Anal. 2010, 52, 809–818. [Google Scholar] [CrossRef]
- Wood, P.L.; Choksi, S.; Bocchini, V. Inducible microglial nitric oxide synthase: A large membrane pool. Neuroreport 1994, 5, 977–980. [Google Scholar] [CrossRef] [PubMed]
- Schnittert, J.; Heinrich, M.A.; Kuninty, P.R.; Storm, G.; Prakash, J. Reprogramming tumor stroma using an endogenous lipid lipoxin A4 to treat pancreatic cancer. Cancer Lett. 2018, 420, 247–258. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| VLCDCA | Molecular Formula | Published Nomenclature [17,18,19,20,21,22] | Exact Mass | [M − H]− | R | ppm Mass Error |
|---|---|---|---|---|---|---|
| [2H28] DCA 16:0 | Internal std. | 314.39016 | 313.3829 | 1.60 | ||
| 28:0 | C28H54O4 | 454.4022 | 453.3949 | 0.067 | 1.56 | |
| 28:1 | C28H52O4 | GTA-452 | 452.3866 | 451.3793 | 0.276 | 1.30 |
| 28:2 | C28H50O4 | GTA-450 | 450.3709 | 449.3636 | 0.773 | 1.73 |
| 28:3 | C28H48O4 | GTA-448 | 448.3553 | 447.3480 | 1.084 | 1.24 |
| 28:4 | C28H46O4 | GTA-446 | 446.3396 | 445.3323 | 0.778 | 1.18 |
| 28:5 | C28H44O4 | 444.3240 | 443.3167 | 0.047 | 0.22 |
 | ||||
|---|---|---|---|---|
| Fragment | Formula | Calculated Anion | ppm Std. | ppm Extract |
| VLCDCA 28:4 | C28H46O4 | 445.3323 | 0.62 | 1.07 |
| H2O Loss | C28H44O3 | 427.3217 | 0.39 | 0.30 |
| CO2 and H2O Loss | C27H44O | 383.3319 | 0.05 | 0.44 |
| CO2 Loss | C27H46O2 | 401.3425 | 0.07 | 0.36 |
| ω-1 | C6H10O2 | 113.0608 | 0.09 | 0.14 |
| ω-2 | C9H14O2 | 153.0921 | 0.07 | 0.30 |
| ω-2-H2O | C9H12O | 135.0815 | 0.14 | 0.89 |
| α-1 | C13H24O2 | 211.1703 | 0.14 | 0.66 |
| α-1-H2O | C13H22O | 193.1597 | 0.31 | 0.92 |
| α-2 | C16H28O2 | 251.2016 | 0.34 | 0.48 |
| α-2-H2O | C16H26O | 233.1910 | 0.13 | 0.13 |
| Biofluid | R (N) |
|---|---|
| Adult human plasma | 0.79 (12) |
| Human synovial fluid | 0.50 (3) |
| Human pleural fluid | 0.14 (3) |
| Human cerebrospinal fluid | 0.045 (12) |
| Human umbilical cord plasma | 0.85 (6) |
| Human aqueous humor | 0.024 (2) |
| Chimpanzee plasma | 0.061 (2) |
| Cynomologous monkey plasma | 0.0022 (2) |
| Rhesus macaque plasma | ND (1) |
| Dog plasma | ND (6) |
| Rat plasma | ND (6) |
| Cow plasma | ND (6) |
| Cat plasma | ND (6) |
| Group | Age (Yr. ±SD) | N | Number of Females |
|---|---|---|---|
| Pilot-Controls | 54.5 ± 8.9 | 15 | 9 |
| Pilot-Kidney | 65.8 ± 12.0 | 7 | 1 |
| Pilot-Breast | 59.4 ± 7.4 | 7 | 7 |
| Pilot-GBM | 54.6 ± 5.6 | 12 | 5 |
| Pilot-CRC | 69.9 ± 6.9 | 9 | 5 |
| Validation-Controls | 57.3 ±5.8 | 23 | 14 |
| Validation-Pancreatic | 69.1 ± 9.3 | 18 | 11 |
| Validation-CRC | 57.3 ± 8.1 | 28 | 22 |
| Validation-Kidney | 55.7 ± 8.9 | 9 | 2 |
© 2018 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wood, P.L. Endogenous Anti-Inflammatory Very-Long-Chain Dicarboxylic Acids: Potential Chemopreventive Lipids. Metabolites 2018, 8, 76. https://doi.org/10.3390/metabo8040076
Wood PL. Endogenous Anti-Inflammatory Very-Long-Chain Dicarboxylic Acids: Potential Chemopreventive Lipids. Metabolites. 2018; 8(4):76. https://doi.org/10.3390/metabo8040076
Chicago/Turabian StyleWood, Paul L. 2018. "Endogenous Anti-Inflammatory Very-Long-Chain Dicarboxylic Acids: Potential Chemopreventive Lipids" Metabolites 8, no. 4: 76. https://doi.org/10.3390/metabo8040076
APA StyleWood, P. L. (2018). Endogenous Anti-Inflammatory Very-Long-Chain Dicarboxylic Acids: Potential Chemopreventive Lipids. Metabolites, 8(4), 76. https://doi.org/10.3390/metabo8040076