
Fermentative Production of the Diamine Putrescine: System Metabolic Engineering of Corynebacterium Glutamicum



Abstract
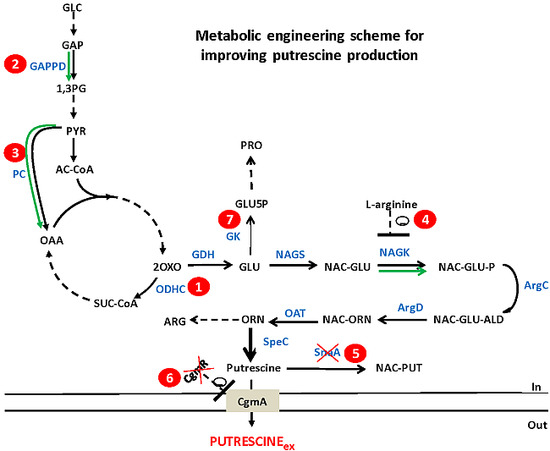
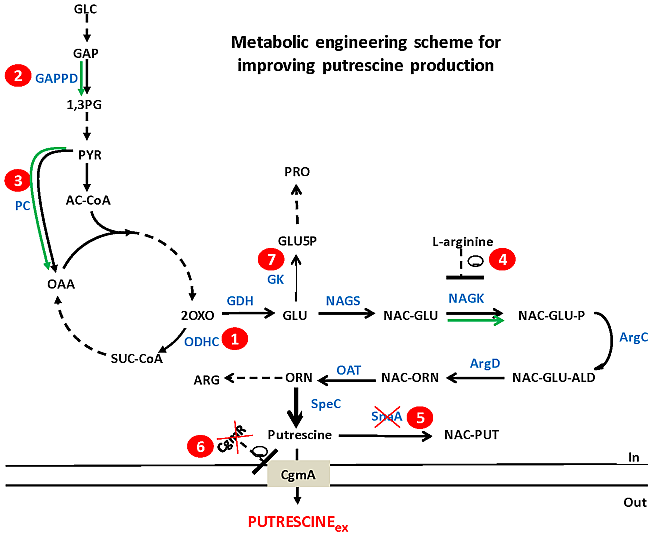
:
1. Introduction
2. Results and Discussion
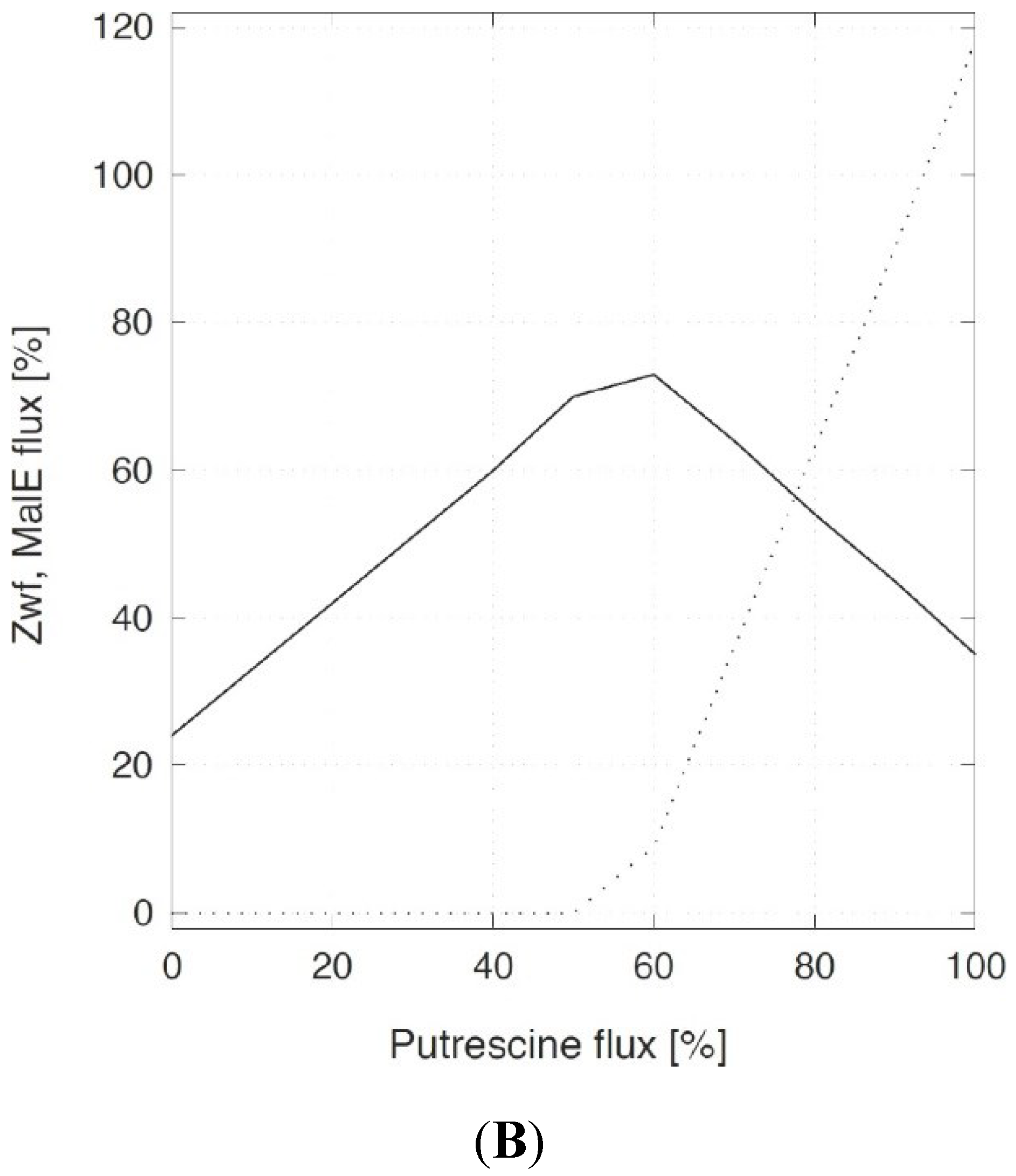
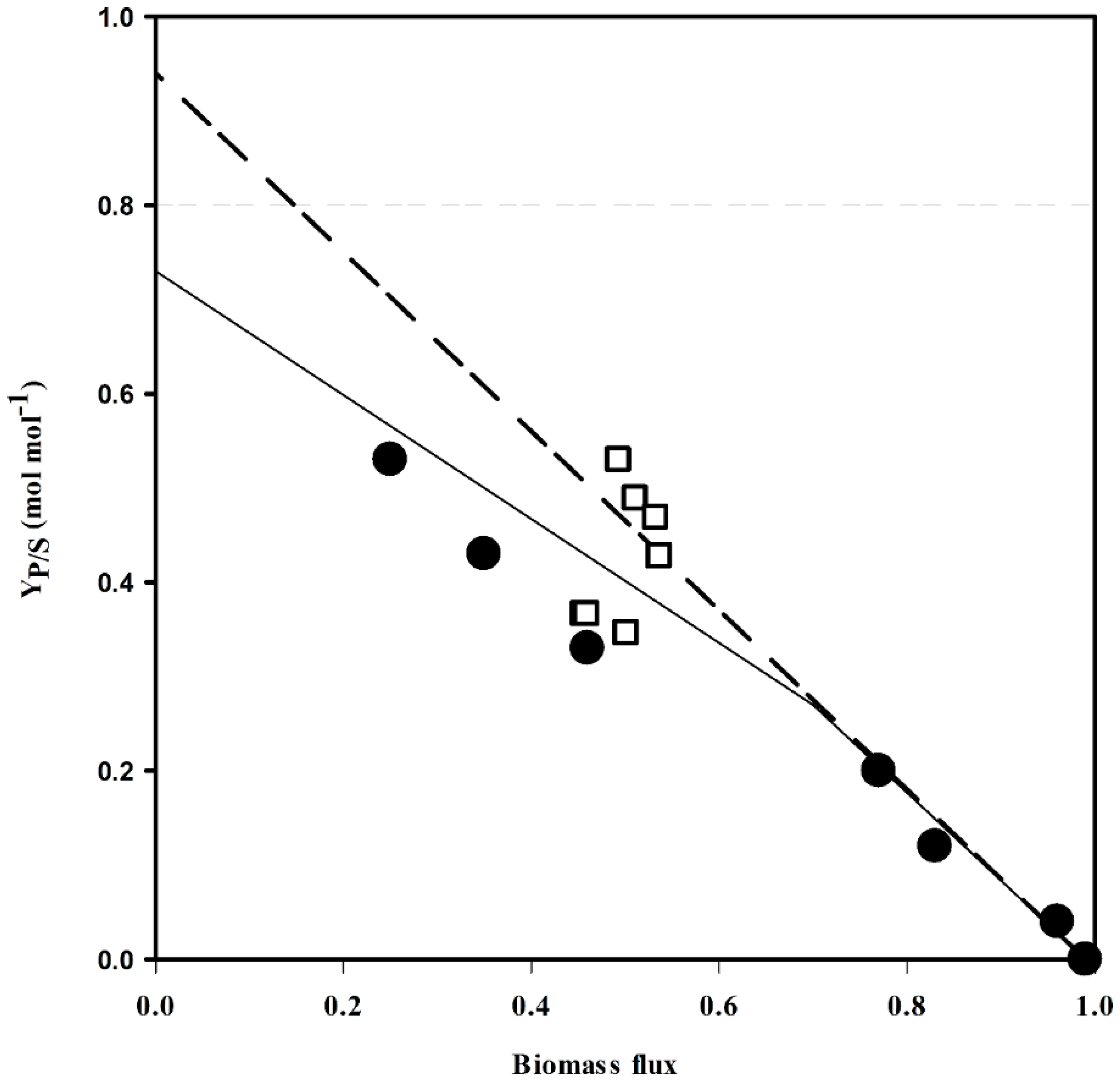
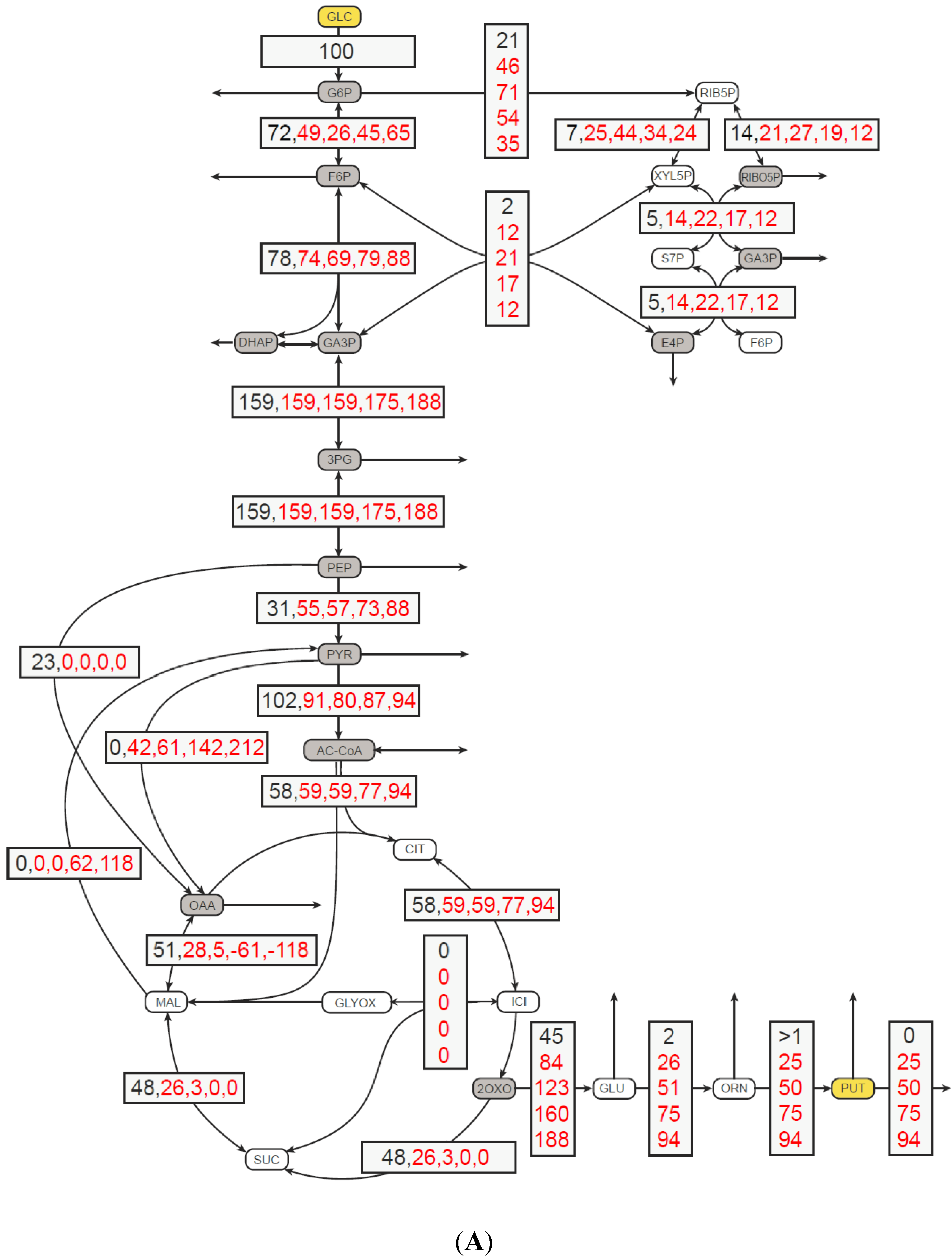
2.1. In Silico Characterization of Putrescine Production in C. glutamicum

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
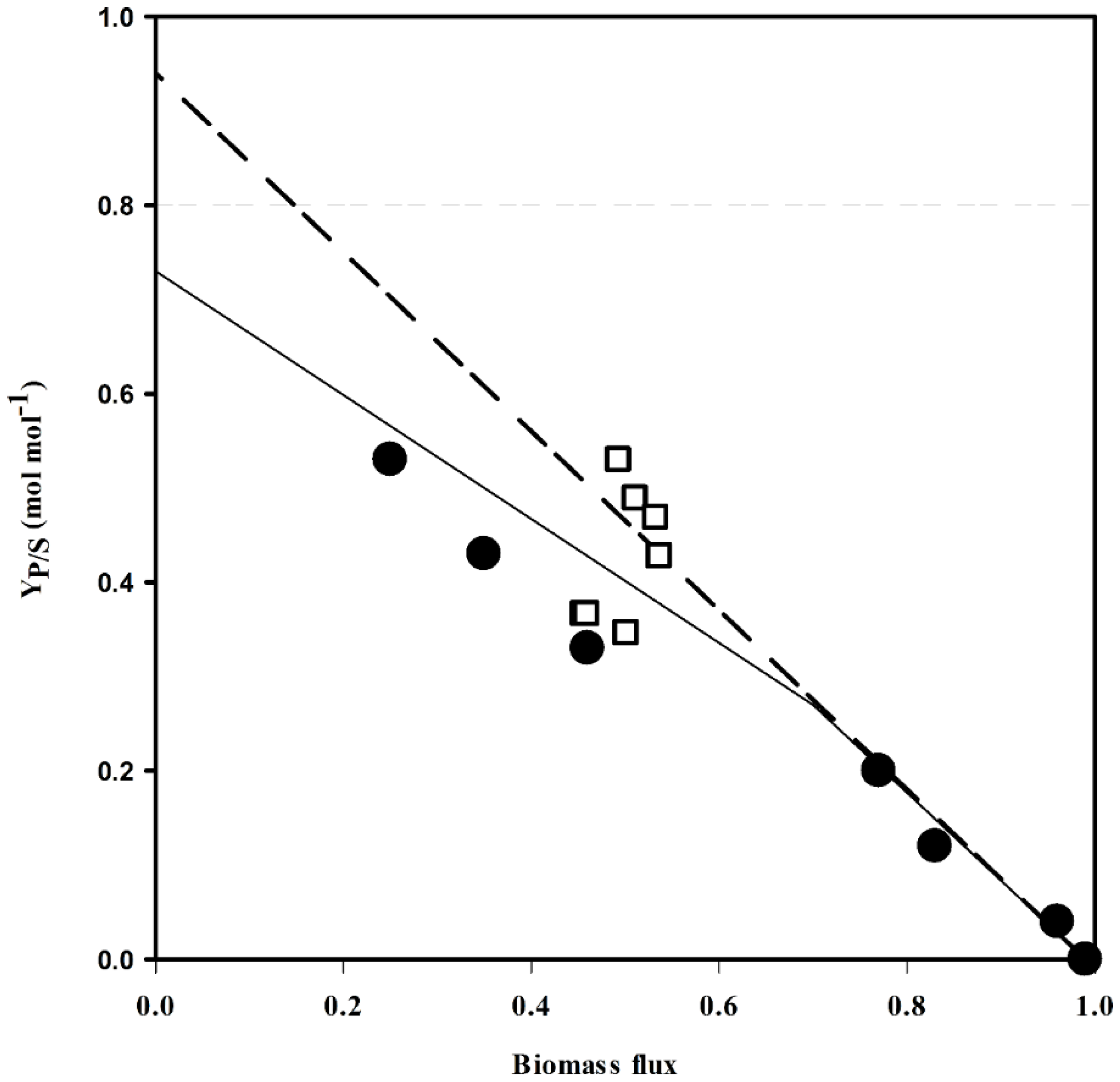
| Carbon Source | Κ | YP/S [mol-C·mol-C−1] |
|---|---|---|
| Glucose | 4.0 | 0.627 |
| Glycerol | 4.7 | 0.653 |
| Lactate | 4.0 | 0.511 |
| Acetate | 4.0 | 0.464 |

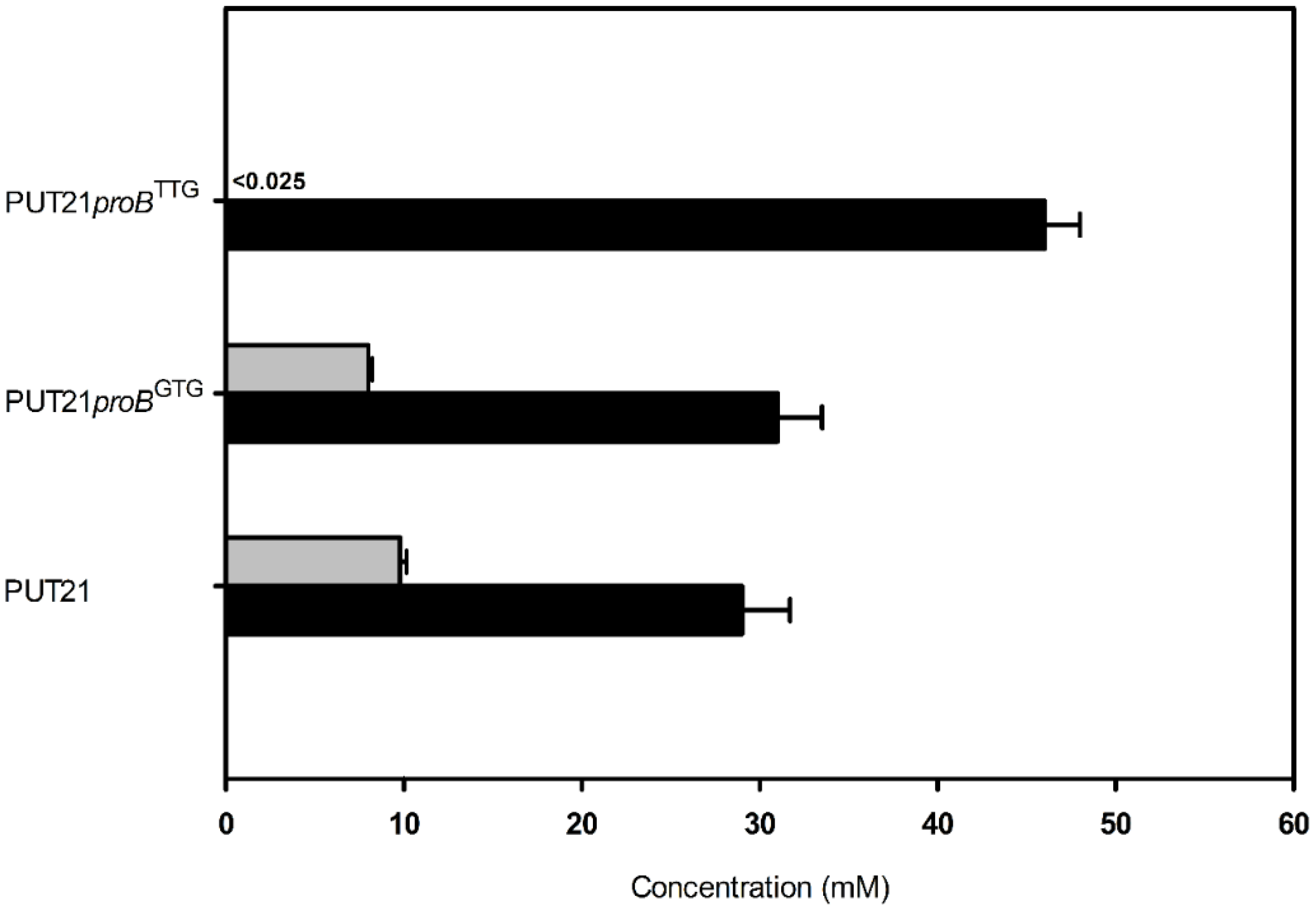
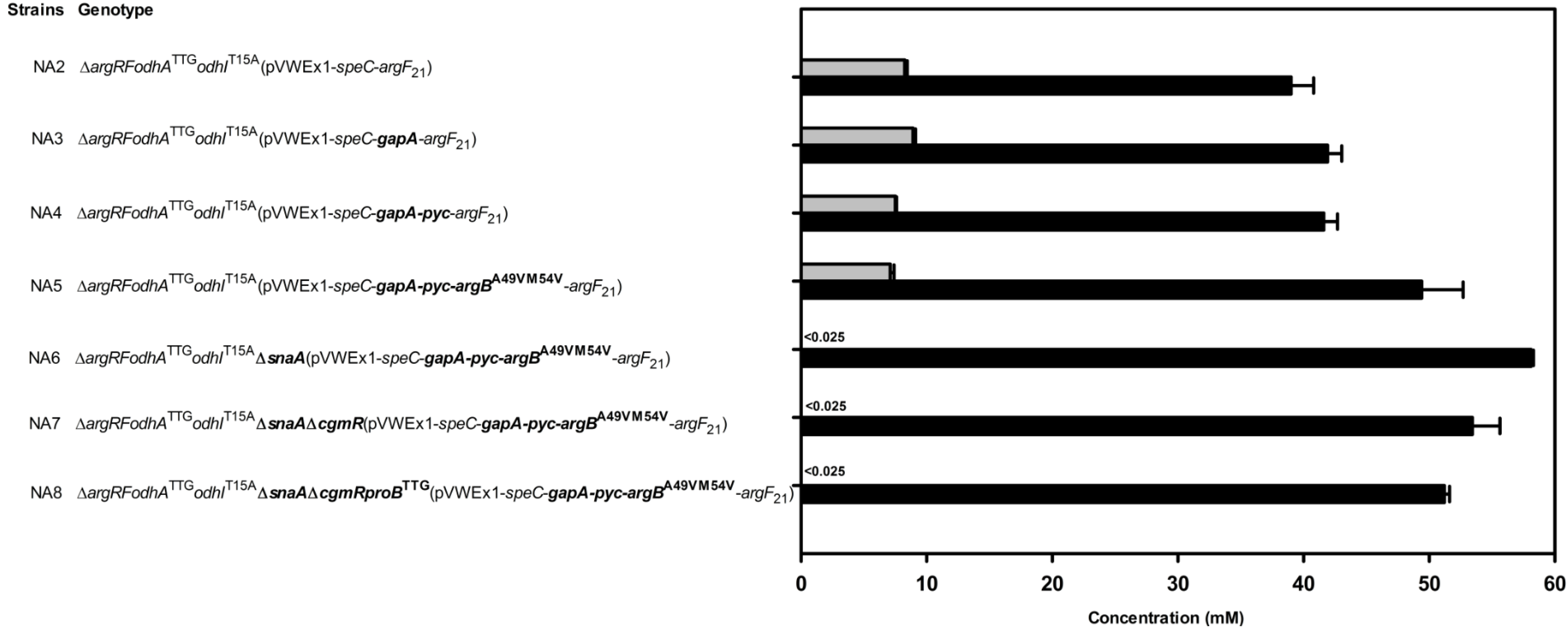
2.2. Reducing 2-Oxoglutarate Dehydrogenase Activity as a Target to Increase Putrescine Production
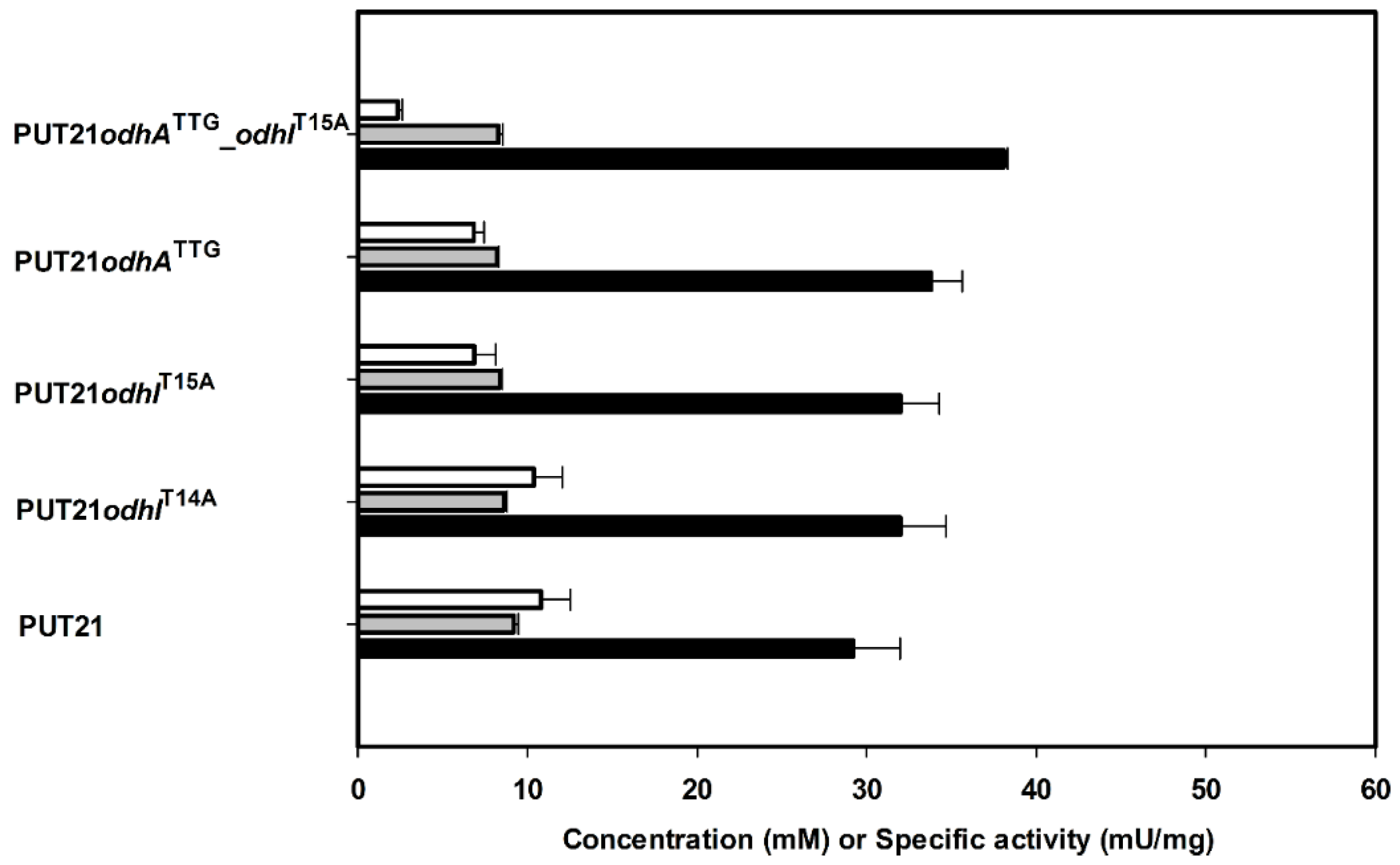
| Strain | Growth Rate µb (h−1) | Putrescine Accumulation (mM) | Product Yield YP/Sb (g·g−1) | Biomass Yield YX/S b (g·g−1) | Volumetric Productivity Qp c (g·L−1·h−1) | Specific Productivity qpc (g·g−1·h−1) |
|---|---|---|---|---|---|---|
| PUT21 | 0.19 | 29.2 ± 2.8 | 0.13 | 0.28 | 0.11 | 0.020 |
| PUT21proBGTG | 0.22 | 31.2 ± 3.0 | 0.14 | 0.27 | 0.12 | 0.022 |
| PUT21proBTTG | 0.14 | 46.1 ± 2.8 | 0.20 | 0.29 | 0.14 | 0.024 |
| NA2 | 0.17 | 38.1 ± 0.2 | 0.17 | 0.25 | 0.14 | 0.028 |
| NA3 | 0.16 | 41.9 ± 1.1 | 0.18 | 0.22 | 0.14 | 0.032 |
| NA4 | 0.16 | 41.6 ± 1.1 | 0.18 | 0.26 | 0.14 | 0.027 |
| NA5 | 0.15 | 48.3 ± 3.3 | 0.21 | 0.23 | 0.19 | 0.041 |
| NA6 | 0.17 | 58.1 ± 0.2 | 0.26 | 0.23 | 0.21 | 0.045 |
| NA7 | 0.20 | 53.4 ± 2.2 | 0.24 | 0.24 | 0.20 | 0.042 |
| NA8 | 0.20 | 51.2 ± 0.5 | 0.23 | 0.24 | 0.20 | 0.041 |
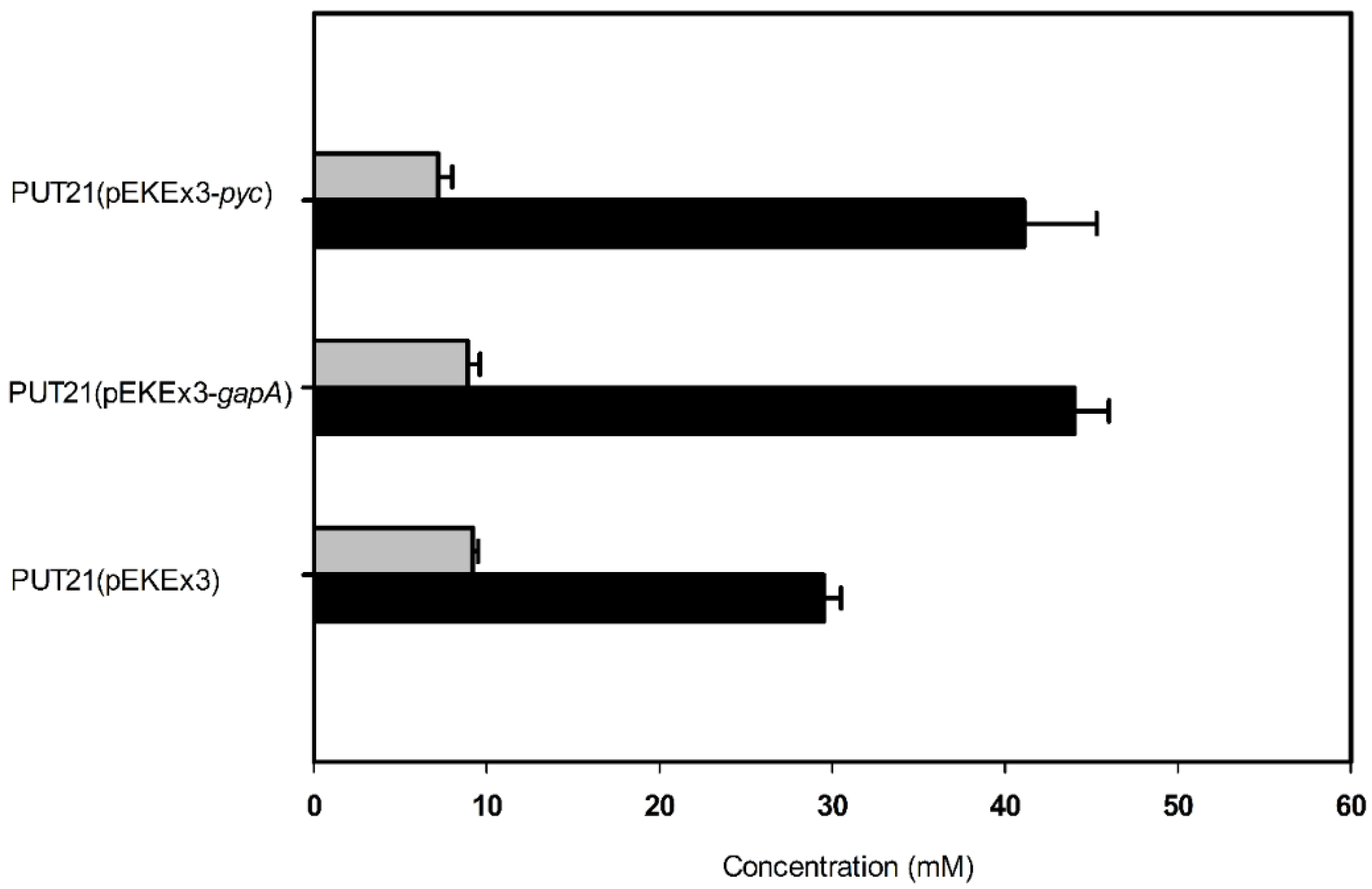
2.3. Increasing Precursor Supply Flux for Putrescine Production
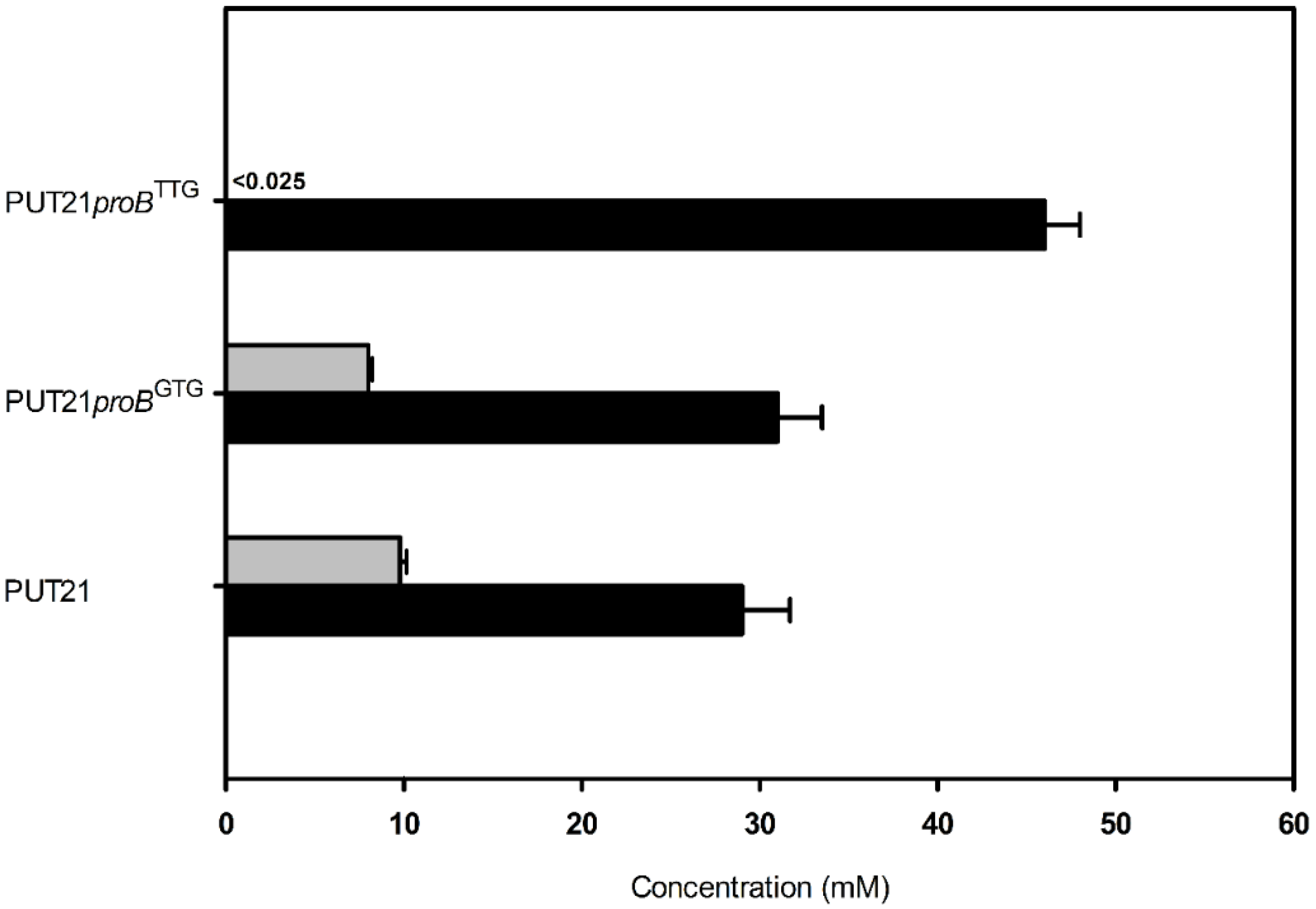
2.4. Decreasing Conversion of Glutamate to Proline
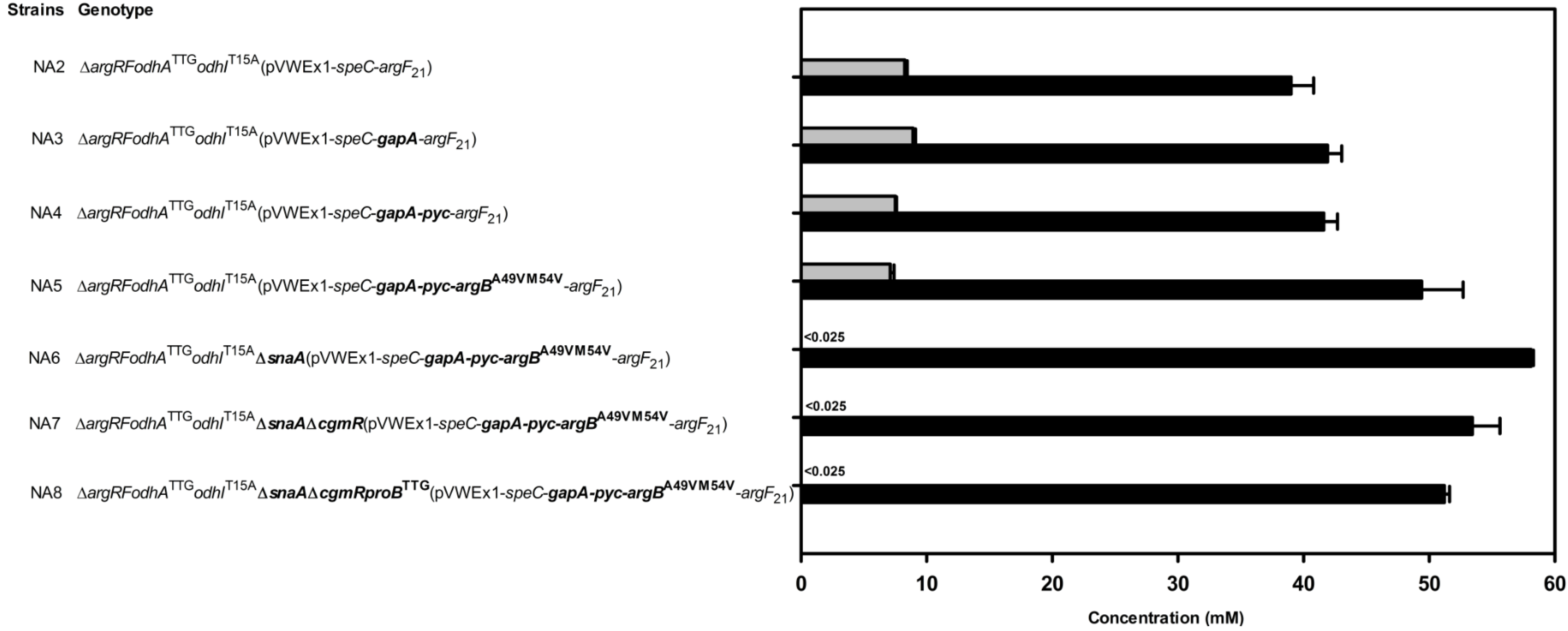
2.5. Combinatorial Construction of Putrescine Overproducing C. glutamicum Strains
3. Experimental Section
3.1. Bacterial Strains and Culture Conditions
| Name | Relevant Genotype/Information | Refs. |
|---|---|---|
| Strains | ||
| ORN1 | In-frame deletion of argR and argF, l-ornithine overproducing strain derived from C. glutamicum ATCC13032; auxotrophic for l-arginine | [5] |
| PUT21 | ORN1 carrying plasmid pVWEx1-speC-argF21 | [4] |
| PUT21odhATTG | PUT21 with replacement of translational start codon GTG of chromosomal odhA of by TTG | This study |
| PUT21odhIT14A | PUT21 with replacement of threonine codon 14 of odhI by an alanine codon | This study |
| PUT21odhIT15A | PUT21 with replacement of threonine codon 15 of chromosomal odhI by an alanine codon | This study |
| PUT21proBGTG | PUT21 with replacement of translational start codon ATG of chromosomal proB by GTG | This study |
| PUT21proBTTG | PUT21 with replacement of translational start codon ATG of chromosomal proB by TTG | This study |
| NA2 | PUT21odhATTG with replacement of threonine codon 15 of chromosomal odhI by an alanine codon | This study |
| NA3 | NA2, but carrying plasmid pVWEx1-speC-gapA-argF21 instead of pVWEx1-speC-argF21 | This study |
| NA4 | NA2, but carrying plasmid pVWEx1-speC-gapA-pyc-argF21 instead of pVWEx1-speC-argF21 | This study |
| NA5 | NA2, but carrying plasmid pVWEx1-speC-gapA-pyc-argBA49V/M54V-argF21 instead of pVWEx1-speC-argF21 | This study |
| NA6 | NA5 with chromosomal deletion of snaA | This study |
| NA7 | NA6 with chromosomal deletion of cgmR | This study |
| NA8 | NA7 with replacement of translational start codon ATG of chromosomal proB by TTG | This study |
| Plasmids | ||
| pEKEx3 | SpecR; Ptac, lacIq; pBL1, oriVC.g., oriVE.c. | [44] |
| pEKEx3-gapA | SpeCR, pEKEx3 overexpressing gapA from C. glutamicum ATCC13032 | [30] |
| pEKEx3-pyc | SpeCR, pEKEx3 overexpressing pyc from C. glutamicum ATCC13032 | [27] |
| pEKEx3-argBA49V/M54V | SpeCR, pEKEx3 overexpressing argBA49V/M54V from C. glutamicum ATCC13032 | [36] |
| pVWEx1-speC-argF21 | KanR , plasmid-based overexpressing speC from E. coli MG1655 and leaky expression of argF in pVWEx1 | [4] |
| pK19mobsacBodhATTG | KanR; mobilizable vector for the replacement of start codon of odhA from GTG to TTG | This study |
| pK19mobsacBodhIT14A | KanR; mobilizable vector for the replacement of threonine 14 in odhI by alanine | This study |
| pVWEx1-speC-gapA-pyc-argBA49V/M54V-argF21 | KanR, KanR, plasmid-based overexpressing argBA49V/M54V, pyc and gapA from C. glutamicum, speC from E. coli MG1655 and leaky expression of argF | This study |
| pK19mobsacB | KanR; mobilizable E. coli vector for the construction of deletion mutants in C. glutamicum (oriVE.c. , PT7, lacI) | [45] |
| pK19mobsacBodhATTG | KanR; mobilizable vector for the replacement of start codon of odhA from GTG to TTG | This study |
| pK19mobsacBodhIT14A | KanR; mobilizable vector for the replacement of threonine 14 in odhI by alanine | This study |
| pK19mobsacBodhIT15A | KanR; mobilizable vector for the replacement of threonine 15 in odhI by alanine | This study |
| pK19mobsacBproBGTG | KanR; mobilizable vector for replacement of native start codon ATG of proB by GTG | This study |
| pK19mobsacBproBTTG | KanR; mobilizable vector for replacement of native start codon ATG of proB by TTG | This study |
| pK19mobsacBΔsnaA | KanR; mobilizable vector for deletion of snaA | [39] |
| pK19mobsacBΔcgmR | KanR; mobilizable vector for deletion of cgmR | [39] |
3.2. Strain Construction
| Primer names | Sequence (5′-3′) |
|---|---|
| odhI141 | CGAATCCATTCACCTGC |
| odhI142 | ACTGAGGTGGCCTCGACCTG |
| odhI143 | CAGGTCGAGGCCACCTCAGT |
| odhI144 | GCAACCGCACTGTTTG |
| odhI152 | ACTGAGGCGGTCTCGACCTG |
| odhI153 | CAGGTCGAGACCGCCTCAGT |
| odhA1 | CCTGATGGTTTCAACCATCAAGTC |
| odhA2 | AGTACTAGCGCTGCTCAAGGCAGG |
| odhA3 | CTGCCTTGAGCAGCGCTAGTAC |
| odhA4 | CCATGGCGTAGCCAATGATG |
| gtg1 | AGCAGTTGGCTACCTGG |
| gtg2 | CACCGGCGCCACTTGGGTTG |
| gtg3 | CAACCCAAGTGGCGCCGGTG |
| gtg4 | GGCAAAAGAACGTCCCC |
| ttg2 | ACCGGCGCCAATTGGGTTGG |
| ttg3 | CCAACCCAATTGGCGCCGGT |
| gapA-cgl-fw | AAGGAGATATAGATATGACCATTCGTGTTGGTATTAAC |
| gapA-cgl-rv | TTAGAGCTTGGAAGCTACGAGCTC |
| ACBF-gapA1 | TTGTACGGTTATGTGTTGAAGTAAGGATCCGAAAGGAGGCCCTTCAGATGACCATTCGTGTTGGTATTA |
| ACBF-gapA2 | ATCTGAAGGGCCTCCTTTCACATGTTTAGAGCTTGGAAGCTACGAG |
| ACBF-gapA3 | AGTGAATTCGAGCTCGGTACCCGGGCATATGTTAGAGCTTGGAAGCTACGAG |
| ACBF-pyc1 | CTCGTAGCTTCCAAGCTCTAAACATGTGAAAGGAGGCCCTTCAGATGTCGACTCACACATCTTC |
| ACBF-pyc2 | GGCCTCCTTTCGCGGCCGCTTAGGAAACGACGACGATCA |
| ACBF-pyc3 | AGTGAATTCGAGCTCGGTACCCGGGCATATGTTAGGAAACGACGACGATCA |
| ACBF-argB1 | TGATCGTCGTCGTTTCCTAAGCGGCCGCGAAAGGAGGCCCTTCAGTTG |
| ACBF-argB2 | AGTGAATTCGAGCTCGGTACCCGGGCATATGTTACAGTTCCCCATCCTTGTC |
3.3. Flux Balance Analysis
3.4. Enzyme Assays
3.5. Quantification of Putrescine
4. Conclusions
Supplementary Files
Supplementary File 1Acknowledgments
Author Contributions
Conflicts of Interest
References
- Schaffer, S.; Haas, T. Biocatalytic and fermentative production of alpha,omega-bifunctional polymer precursors. Org. Process. Res. Dev. 2014, 18, pp. 752–766. Available online: http://dx.doi.org/10.1021/op5000418 (accessed on 11 February 2015).
- Rydz, J.; Sikorska, W.; Kyulavska, M.; Christova, D. Polyester-based (bio)degradable polymers as environmentally friendly materials for sustainable development. Int. J. Mol. Sci. 2014, 16, 564–596. [Google Scholar] [CrossRef] [PubMed]
- Kroschwitz, J.I.; Seidel, A. Kirk-Othmer Encyclopedia of Chemical Technology, 5th ed.; Wiley-Interscienc: Hoboken, NJ, USA, 2004. [Google Scholar]
- Schneider, J.; Eberhardt, D.; Wendisch, V.F. Improving putrescine production by Corynebacterium glutamicum by fine-tuning ornithine transcarbamoylase activity using a plasmid addiction system. Appl. Microbiol. Biotechnol. 2012, 95, 169–178. [Google Scholar] [CrossRef] [PubMed]
- Schneider, J.; Wendisch, V.F. Putrescine production by engineered Corynebacterium glutamicum. Appl. Microbiol. Biotechnol. 2010, 88, 859–868. [Google Scholar] [CrossRef] [PubMed]
- Qian, Z.G.; Xia, X.X.; Lee, S.Y. Metabolic engineering of Escherichia coli for the production of putrescine: A four carbon diamine. Biotechnol. Bioeng. 2009, 104, 651–662. [Google Scholar] [PubMed]
- Meiswinkel, T.M.; Rittmann, D.; Lindner, S.N.; Wendisch, V.F. Crude glycerol-based production of amino acids and putrescine by Corynebacterium glutamicum. Bioresour. Technol. 2013, 145, 254–258. [Google Scholar] [CrossRef] [PubMed]
- Meiswinkel, T.M.; Gopinath, V.; Lindner, S.N.; Nampoothiri, K.M.; Wendisch, V.F. Accelerated pentose utilization by Corynebacterium glutamicum for accelerated production of lysine, glutamate, ornithine and putrescine. Microb. Biotechnol. 2013, 6, 131–140. [Google Scholar] [CrossRef] [PubMed]
- Uhde, A.; Youn, J.W.; Maeda, T.; Clermont, L.; Matano, C.; Kramer, R.; Wendisch, V.F.; Seibold, G.M.; Marin, K. Glucosamine as carbon source for amino acid-producing Corynebacterium glutamicum. Appl. Microbiol. Biotechnol. 2013, 97, 1679–1687. [Google Scholar] [CrossRef] [PubMed]
- Meiswinkel, T.M.; Lindner, S.N.; Wendisch, V.F. Thick juice-based production of amino acids and putrescine by Corynebacterium glutamicum. J. Biotechnol. Biomater. 2014. [Google Scholar] [CrossRef]
- Peters-Wendisch, P.; Gotker, S.; Heider, S.A.; Komati Reddy, G.; Nguyen, A.Q.; Stansen, K.C.; Wendisch, V.F. Engineering biotin prototrophic Corynebacterium glutamicum strains for amino acid, diamine and carotenoid production. J. Biotechnol. 2014, 192, 346–354. [Google Scholar] [CrossRef] [PubMed]
- Shinfuku, Y.; Sorpitiporn, N.; Sono, M.; Furusawa, C.; Hirasawa, T.; Shimizu, H. Development and experimental verification of a genome-scale metabolic model for Corynebacterium glutamicum. Microb. Cell. Fact. 2009. [Google Scholar] [CrossRef]
- Blombach, B.; Riester, T.; Wieschalka, S.; Ziert, C.; Youn, J.W.; Wendisch, V.F.; Eikmanns, B.J. Corynebacterium glutamicum tailored for efficient isobutanol production. Appl. Environ. Microbiol. 2011, 77, 3300–3310. [Google Scholar] [CrossRef] [PubMed]
- Rittmann, D.; Lindner, S.N.; Wendisch, V.F. Engineering of a glycerol utilization pathway for amino acid production by Corynebacterium glutamicum. Appl. Environ. Microbiol. 2008, 74, 6216–6222. [Google Scholar] [CrossRef] [PubMed]
- Hadiati, A.; Krahn, I.; Lindner, S.N.; Wendisch, V.F. Engineering of Corynebacterium glutamicum for growth and production of l-ornithine, l-lysine, and lycopene from hexuronic acids. Bioresour. Bioprocess. 2014. [Google Scholar] [CrossRef]
- Usuda, Y.; Tujimoto, N.; Abe, C.; Asakura, Y.; Kimura, E.; Kawahara, Y.; Kurahashi, O.; Matsui, H. Molecular cloning of the Corynebacterium glutamicum (“Brevibacterium lactofermentum” AJ12036) odhA gene encoding a novel type of 2-oxoglutarate dehydrogenase. Microbiology 1996, 142, 3347–3354. [Google Scholar] [CrossRef] [PubMed]
- Schwinde, J.W.; Hertz, P.F.; Sahm, H.; Eikmanns, B.J.; Guyonvarch, A. Lipoamide dehydrogenase from Corynebacterium glutamicum: Molecular and physiological analysis of the lpd gene and characterization of the enzyme. Microbiology 2001, 147, 2223–2231. [Google Scholar] [PubMed]
- Niebisch, A.; Kabus, A.; Schultz, C.; Weil, B.; Bott, M. Corynebacterial protein kinase G controls 2-oxoglutarate dehydrogenase activity via the phosphorylation status of the OdhI protein. J. Biol. Chem. 2006, 281, 12300–12307. [Google Scholar] [CrossRef] [PubMed]
- Kawahara, Y.; Takahashi-Fuke, K.; Shimizu, E.; Nakamatsu, T.; Nakamori, S. Relationship between the glutamate production and the activity of 2-oxoglutarate dehydrogenase in Brevibacterium lactofermentum. Biosci. Biotechnol. Biochem. 1997, 61, pp. 1109–1112. Available online: http://dx.doi.org/10.1271/bbb.61.1109 (accessed on 24 July 2014).
- Asakura, Y.; Kimura, E.; Usuda, Y.; Kawahara, Y.; Matsui, K.; Osumi, T.; Nakamatsu, T. Altered metabolic flux due to deletion of odhA causes l-glutamate overproduction in Corynebacterium glutamicum. Appl. Environ. Microbiol. 2006, 73, 1308–1319. [Google Scholar] [CrossRef] [PubMed]
- Raasch, K.; Bocola, M.; Labahn, J.; Leitner, A.; Eggeling, L.; Bott, M. Interaction of 2-oxoglutarate dehydrogenase OdhA with its inhibitor OdhI in Corynebacterium glutamicum: Mutants and a model. J. Biotechnol. 2014, 191, 99–105. [Google Scholar] [CrossRef] [PubMed]
- Krawczyk, S.; Raasch, K.; Schultz, C.; Hoffelder, M.; Eggeling, L.; Bott, M. The FHA domain of OdhI interacts with the carboxyterminal 2-oxoglutarate dehydrogenase domain of OdhA in Corynebacterium glutamicum. FEBS Lett. 2010, 584, 1463–1468. [Google Scholar] [CrossRef] [PubMed]
- Schultz, C.; Niebisch, A.; Schwaiger, A.; Viets, U.; Metzger, S.; Bramkamp, M.; Bott, M. Genetic and biochemical analysis of the serine/threonine protein kinases PknA, PknB, PknG and PknL of Corynebacterium glutamicum: Evidence for non-essentiality and for phosphorylation of OdhI and FtsZ by multiple kinases. Mol. Microbiol. 2009, 74, 724–741. [Google Scholar] [CrossRef] [PubMed]
- Schultz, C.; Niebisch, A.; Gebel, L.; Bott, M. Glutamate production by Corynebacterium glutamicum: Dependence on the oxoglutarate dehydrogenase inhibitor protein OdhI and protein kinase PknG. Appl. Microbiol. Biotechnol. 2007, 76, 691–700. [Google Scholar] [CrossRef] [PubMed]
- Yukawa, H.; Inui, M.; Vertes, A.A. Genomes and genome-level engineering of amino acid-producing bacteria. In Amino Acid Biosynthesis—Pathways, Regulation and Metabolic Engineering; Wendisch, V.F., Ed.; Springer: Heidelberg, Germany, 2007; Volume 5, pp. 349–401. [Google Scholar] [CrossRef]
- Peters-Wendisch, P.G.; Wendisch, V.F.; Paul, S.; Eikmanns, B.J.; Sahm, H. Pyruvate carboxylase as an anaplerotic enzyme in Corynebacterium glutamicum. Microbiology 1997, 143, 1095–1103. [Google Scholar] [CrossRef]
- Peters-Wendisch, P.G.; Schiel, B.; Wendisch, V.F.; Katsoulidis, E.; Mockel, B.; Sahm, H.; Eikmanns, B.J. Pyruvate carboxylase is a major bottleneck for glutamate and lysine production by Corynebacterium glutamicum. J. Mol. Microbiol. Biotechnol. 2001, 3, 295–300. [Google Scholar] [PubMed]
- Jojima, T.; Fujii, M.; Mori, E.; Inui, M.; Yukawa, H. Engineering of sugar metabolism of Corynebacterium glutamicum for production of amino acid l-alanine under oxygen deprivation. Appl. Microbiol. Biotechnol. 2010, 87, 159–165. [Google Scholar] [CrossRef] [PubMed]
- Shirai, T.; Fujimura, K.; Furusawa, C.; Nagahisa, K.; Shioya, S.; Shimizu, H. Study on roles of anaplerotic pathways in glutamate overproduction of Corynebacterium glutamicum by metabolic flux analysis. Microb. Cell. Fact. 2007. [Google Scholar] [CrossRef]
- Siedler, S.; Lindner, S.N.; Bringer, S.; Wendisch, V.F.; Bott, M. Reductive whole-cell biotransformation with Corynebacterium glutamicum: Improvement of NADPH generation from glucose by a cyclized pentose phosphate pathway using pfkA and gapA deletion mutants. Appl. Microbiol. Biotechnol. 2013, 97, 143–152. [Google Scholar] [CrossRef] [PubMed]
- Jiang, L.Y.; Chen, S.G.; Zhang, Y.Y.; Liu, J.Z. Metabolic evolution of Corynebacterium glutamicum for increased production of l-ornithine. BMC Biotechnol. 2013. [Google Scholar] [CrossRef]
- Kim, S.Y.; Lee, J.; Lee, S.Y. Metabolic engineering of Corynebacterium glutamicum for the production of l-ornithine. Biotechnol. Bioeng. 2015, 112, 416–421. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.Y.; Shin, H.S.; Park, J.S.; Kim, Y.H.; Min, J. Proline reduces the binding of transcriptional regulator ArgR to upstream of argB in Corynebacterium glutamicum. Appl. Microbiol. Biotechnol. 2010, 86, 235–242. [Google Scholar] [CrossRef] [PubMed]
- Ronsch, H.; Kramer, R.; Morbach, S. Impact of osmotic stress on volume regulation, cytoplasmic solute composition and lysine production in Corynebacterium glutamicum MH20-22B. J. Biotechnol. 2003, 104, 87–97. [Google Scholar] [CrossRef] [PubMed]
- Weinand, M.; Kramer, R.; Morbach, S. Characterization of compatible solute transporter multiplicity in Corynebacterium glutamicum. Appl. Microbiol. Biotechnol. 2007, 76, 701–708. [Google Scholar] [CrossRef] [PubMed]
- Schneider, J.; Niermann, K.; Wendisch, V.F. Production of the amino acids l-glutamate, l-lysine, l-ornithine and l-arginine from arabinose by recombinant Corynebacterium glutamicum. J. Biotechnol. 2011, 154, 191–198. [Google Scholar] [CrossRef] [PubMed]
- Xu, M.; Rao, Z.; Dou, W.; Jin, J.; Xu, Z. Site-directed mutagenesis studies on the l-arginine-binding sites of feedback inhibition in N-acetyl-l-glutamate kinase (NAGK) from Corynebacterium glutamicum. Curr. Microbiol. 2012, 64, 164–172. [Google Scholar] [CrossRef] [PubMed]
- Schendzielorz, G.; Dippong, M.; Grunberger, A.; Kohlheyer, D.; Yoshida, A.; Binder, S.; Nishiyama, C.; Nishiyama, M.; Bott, M.; Eggeling, L. Taking control over control: Use of product sensing in single cells to remove flux control at key enzymes in biosynthesis pathways. ACS Synth. Biol. 2014, 3, 21–29. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, A.Q.; Schneider, J.; Wendisch, V.F. Elimination of polyamine N-acetylation and regulatory engineering improved putrescine production by Corynebacterium glutamicum. J. Biotechnol. 2015, 201, 75–85. [Google Scholar] [CrossRef] [PubMed]
- Itou, H.; Watanabe, N.; Yao, M.; Shirakihara, Y.; Tanaka, I. Crystal structures of the multidrug binding repressor Corynebacterium glutamicum CgmR in complex with inducers and with an operator. J. Mol. Biol. 2010, 403, 174–184. [Google Scholar] [CrossRef] [PubMed]
- Kind, S.; Kreye, S.; Wittmann, C. Metabolic engineering of cellular transport for overproduction of the platform chemical 1,5-diaminopentane in Corynebacterium glutamicum. Metab. Eng. 2011, 13, 617–627. [Google Scholar] [CrossRef] [PubMed]
- Kind, S.; Jeong, W.K.; Schroder, H.; Zelder, O.; Wittmann, C. Identification and elimination of the competing N-acetyldiaminopentane pathway for improved production of diaminopentane by Corynebacterium glutamicum. Appl. Environ. Microbiol. 2010, 76, 5175–5180. [Google Scholar] [CrossRef] [PubMed]
- Eggeling, L.; Bott, M. Handbook of Corynebacterium Glutamicum; CRC Press: Boca Raton, FL, USA, 2005. [Google Scholar]
- Stansen, C.; Uy, D.; Delaunay, S.; Eggeling, L.; Goergen, J.L.; Wendisch, V.F. Characterization of a Corynebacterium glutamicum lactate utilization operon induced during temperature-triggered glutamate production. Appl. Environ. Microbiol. 2005, 71, 5920–5928. [Google Scholar] [CrossRef] [PubMed]
- Schäfer, A.; Tauch, A.; Jäger, W.; Kalinowski, J.; Thierbach, G.; Puhler, A. Small mobilizable multi-purpose cloning vectors derived from the Escherichia coli plasmids pK18 and pK19: selection of defined deletions in the chromosome of Corynebacterium glutamicum. Gene 1994, 145, 69–73. [Google Scholar] [CrossRef] [PubMed]
- Hanahan, D. Studies on transformation of Escherichia coli with plasmids. J. Mol. Biol. 1983, 166, 557–580. [Google Scholar] [CrossRef] [PubMed]
- Gibson, D.G.; Young, L.; Chuang, R.Y.; Venter, J.C.; Hutchison, C.A., 3rd; Smith, H.O. Enzymatic assembly of DNA molecules up to several hundred kilobases. Nat. Methods 2009, 6, 343–345. [Google Scholar] [CrossRef] [PubMed]
- Schellenberger, J.; Que, R.; Fleming, R.M.; Thiele, I.; Orth, J.D.; Feist, A.M.; Zielinski, D.C.; Bordbar, A.; Lewis, N.E.; Rahmanian, S.; et al. Quantitative prediction of cellular metabolism with constraint-based models: the COBRA Toolbox v2.0. Nat. Protoc. 2011, 6, 1290–1307. [Google Scholar] [CrossRef] [PubMed]
- Shen, L.; Haufe, J.; Patel, M.K. Product overview and market projection of emerging bio-based plastics (Utrecht University). 2009. Available online: http://en.european-bioplastics.org (accessed on 9 February 2015).
- Hoffelder, M.; Raasch, K.; van Ooyen, J.; Eggeling, L. The E2 domain of OdhA of Corynebacterium glutamicum has succinyltransferase activity dependent on lipoyl residues of the acetyltransferase AceF. J. Bacteriol. 2010, 192, 5203–5211. [Google Scholar] [CrossRef] [PubMed]
- Omumasaba, C.A.; Okai, N.; Inui, M.; Yukawa, H. Corynebacterium glutamicum glyceraldehyde-3-phosphate dehydrogenase isoforms with opposite, ATP-dependent regulation. J. Mol. Microbiol. Biotechnol. 2004, 8, 91–103. [Google Scholar] [CrossRef] [PubMed]
- Haas, D.; Leisinger, T. N-acetylglutamate 5-phosphotransferase of Pseudomonas aeruginosa. Purification and ligand-directed association-dissociation. Eur. J. Biochem. 1975, 52, 365–375. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nguyen, A.Q.D.; Schneider, J.; Reddy, G.K.; Wendisch, V.F. Fermentative Production of the Diamine Putrescine: System Metabolic Engineering of Corynebacterium Glutamicum. Metabolites 2015, 5, 211-231. https://doi.org/10.3390/metabo5020211
Nguyen AQD, Schneider J, Reddy GK, Wendisch VF. Fermentative Production of the Diamine Putrescine: System Metabolic Engineering of Corynebacterium Glutamicum. Metabolites. 2015; 5(2):211-231. https://doi.org/10.3390/metabo5020211
Chicago/Turabian StyleNguyen, Anh Q. D., Jens Schneider, Gajendar Komati Reddy, and Volker F. Wendisch. 2015. "Fermentative Production of the Diamine Putrescine: System Metabolic Engineering of Corynebacterium Glutamicum" Metabolites 5, no. 2: 211-231. https://doi.org/10.3390/metabo5020211
APA StyleNguyen, A. Q. D., Schneider, J., Reddy, G. K., & Wendisch, V. F. (2015). Fermentative Production of the Diamine Putrescine: System Metabolic Engineering of Corynebacterium Glutamicum. Metabolites, 5(2), 211-231. https://doi.org/10.3390/metabo5020211