
Effects of Maillard Reaction Products on Skeletal Muscle Cells: An In Vitro Study Using C2C12 Myotubes

 , , , , and
, , , , and
Abstract
1. Introduction
2. Materials and Methods
2.1. Preparation of Maillard Reaction Products (MRPs)
2.2. Evaluation of the Antioxidant Activity of MRPs
2.3. Cell Culture
2.4. Measurement of Intracellular Reactive Oxygen Species (ROS) Levels
2.5. Western Blotting
2.6. RNA Isolation and qPCR Analysis
2.7. Immunocytochemistry
2.8. Statistical Analysis
3. Results
3.1. Determination of Antioxidant Activity of MRPs
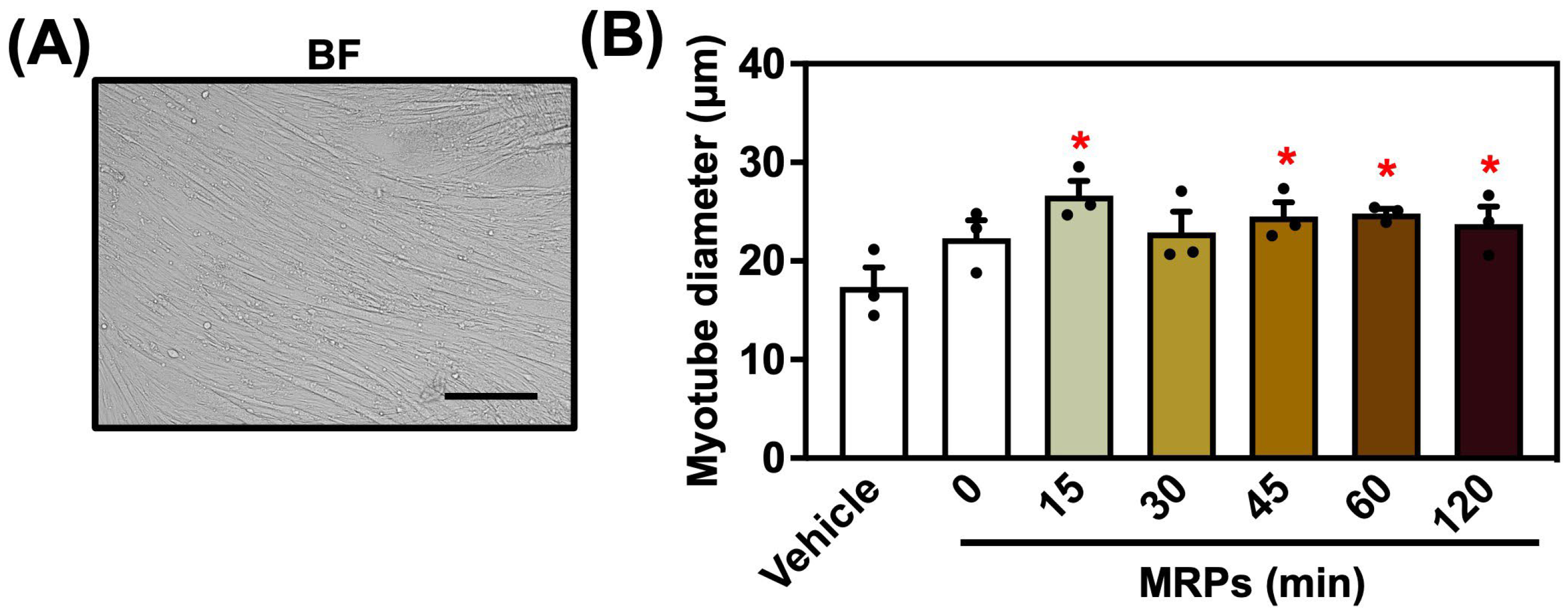
3.2. Effect of MRPs on C2C12 Myotube Diameter
3.3. Effect of MRPs on Protein Synthesis Signaling Pathway and Muscle Fiber Types in C2C12 Myotubes
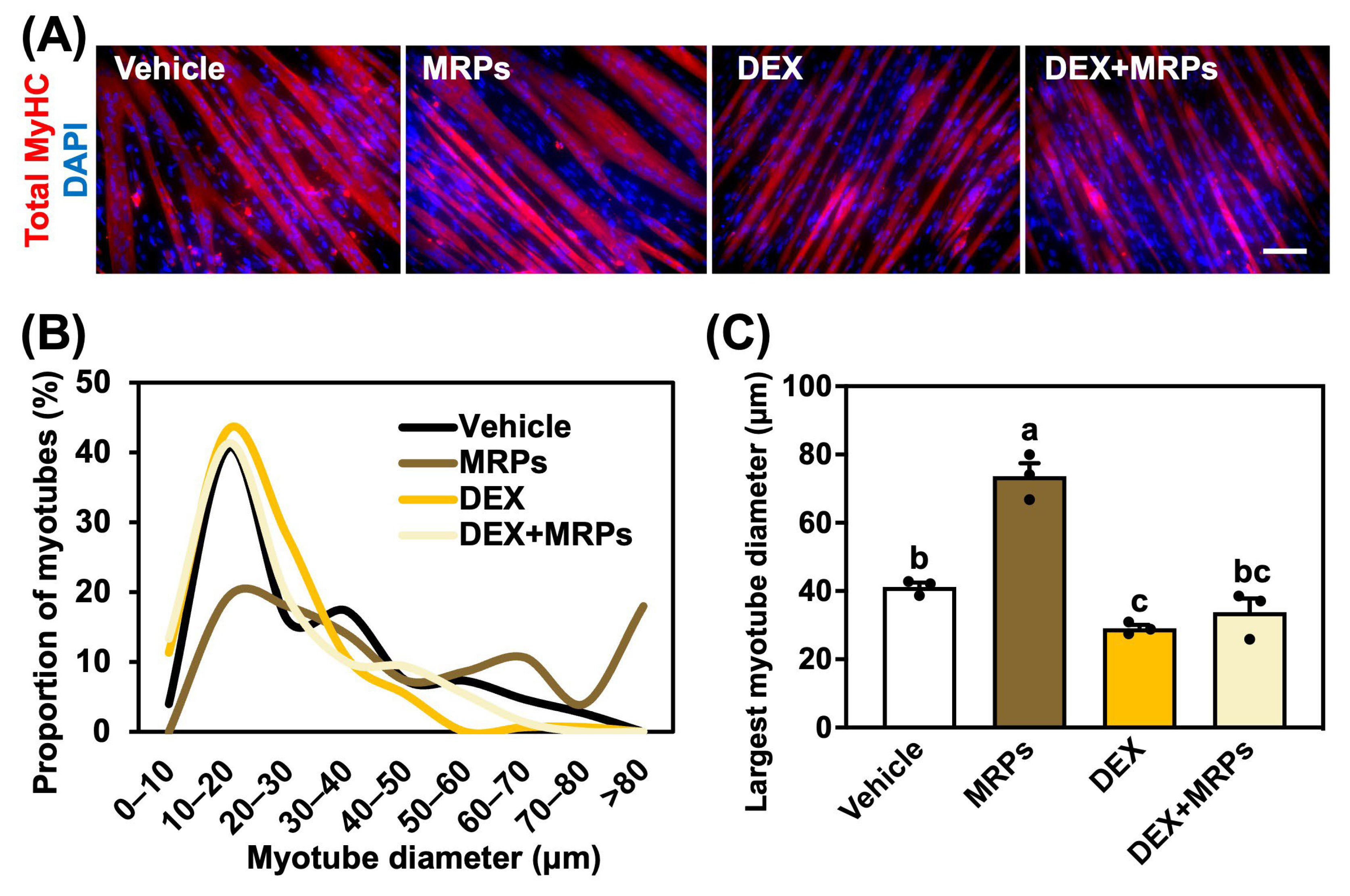
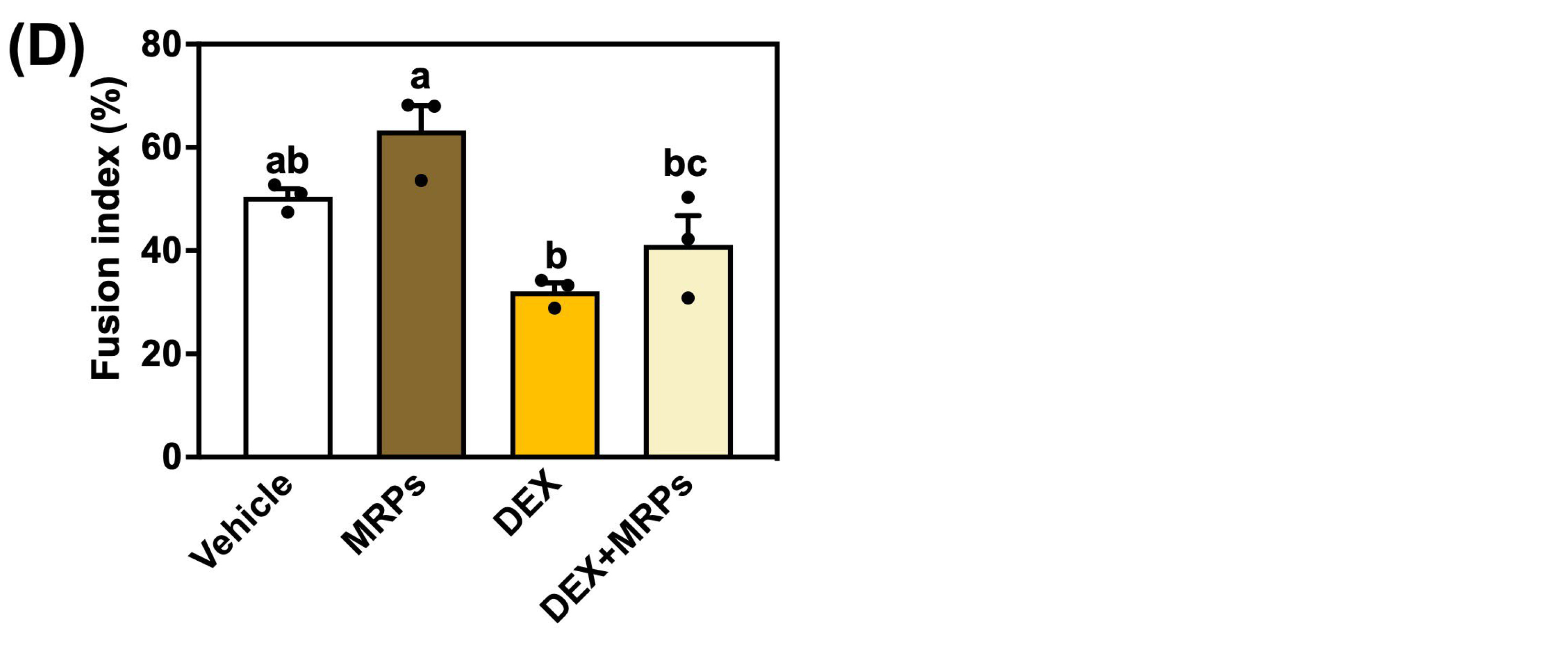
3.4. Effect of MRPs on Dexamethasone-Induced Muscle Atrophy in C2C12 Myotubes
3.5. Effect of MRPs on mRNA Expression in C2C12 Myotubes
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| BSA | Bovine serum albumin |
| EDTA | ethylenediaminetetraacetic acid |
| DAPI | 4′,6-diamidino-2-phenylindole |
| DEX | Dexamethasone |
| DMEM | Dulbecco’s modified Eagle’s medium |
| DPPH | 2,2-diphenyl-1-picrylhydrazylradical |
| DTT | DL-dithiothreitol |
| FBS | Fetal bovine serum |
| HS | Horse serum |
| H2DCFDA | 2′,7′-dichlorofluorescein diacetate |
| MRPs | Maillard reaction products |
| MyHC | Myosin heavy chain |
| PFA | Paraformaldehyde |
| ROS | Reactive oxygen species |
| SDS | Sodium dodecyl sulfate |
| TPBS | Phosphate buffered saline with Tween 20 |
| TTBS | Tris buffered saline with Tween 20 |
References
- Lieber, R.L.; Roberts, T.J.; Blemker, S.S.; Lee, S.S.M.; Herzog, W. Skeletal Muscle Mechanics, Energetics and Plasticity. J. Neuroeng. Rehabil. 2017, 14, 108. [Google Scholar] [CrossRef] [PubMed]
- Smith, J.A.B.; Murach, K.A.; Dyar, K.A.; Zierath, J.R. Exercise Metabolism and Adaptation in Skeletal Muscle. Nat. Rev. Mol. Cell Biol. 2023, 24, 607–632. [Google Scholar] [CrossRef] [PubMed]
- Powers, S.K.; Kavazis, A.N.; DeRuisseau, K.C. Mechanisms of Disuse Muscle Atrophy: Role of Oxidative Stress. Am. J. Physiol.-Regul. Integr. Comp. Physiol. 2005, 288, R337–R344. [Google Scholar] [CrossRef]
- Yin, L.; Li, N.; Jia, W.; Wang, N.; Liang, M.; Yang, X.; Du, G. Skeletal Muscle Atrophy: From Mechanisms to Treatments. Pharmacol. Res. 2021, 172, 105807. [Google Scholar] [CrossRef]
- Biolo, G.; Cederholm, T.; Muscaritoli, M. Muscle Contractile and Metabolic Dysfunction Is a Common Feature of Sarcopenia of Aging and Chronic Diseases: From Sarcopenic Obesity to Cachexia. Clin. Nutr. 2014, 33, 737–748. [Google Scholar] [CrossRef]
- Yadav, A.; Dabur, R. Skeletal Muscle Atrophy after Sciatic Nerve Damage: Mechanistic Insights. Eur. J. Pharmacol. 2024, 970, 176506. [Google Scholar] [CrossRef]
- Bodine, S.; Stitt, T.; Gonzalez, M.; Kline, W.O.; Stover, G.L.; Bauerlein, R.; Zlotchenko, E.; Scrimgeour, A.; Lawrence, J.; Glass, D.; et al. Akt/MTOR Pathway Is a Crucial Regulator of Skeletal Muscle Hypertrophy and Can Prevent Muscle Atrophy In Vivo. Nat. Cell Biol. 2001, 3, 1014–1019. [Google Scholar] [CrossRef]
- Glass, D.J. Molecular Mechanisms Modulating Muscle Mass. Trends Mol. Med. 2003, 9, 344–350. [Google Scholar] [CrossRef]
- Glass, D.J. Skeletal Muscle Hypertrophy and Atrophy Signaling Pathways. Int. J. Biochem. Cell Biol. 2005, 37, 1974–1984. [Google Scholar] [CrossRef]
- Anthony, J.; Anthony, T.; Kimball, S.; Vary, T.; Jefferson, L. Orally Administered Leucine Stimulates Protein Synthesis in Skeletal Muscle of Postabsorptive Rats in Association with Increased EIF4F Formation. J. Nutr. 2000, 130, 139–145. [Google Scholar] [CrossRef]
- Crozier, S.; Kimball, S.; Emmert, S.W.; Anthony, J.; Jefferson, L. Oral Leucine Administration Stimulates Protein Synthesis in Rat Skeletal Muscle. J. Nutr. 2005, 135, 376–382. [Google Scholar] [CrossRef] [PubMed]
- Sakuma, K.; Yamaguchi, A. Novel Intriguing Strategies Attenuating to Sarcopenia. J. Aging Res. 2012, 2012, 251217. [Google Scholar] [CrossRef] [PubMed]
- Montesano, A.; Luzi, L.; Senesi, P.; Mazzocchi, N.; Terruzzi, I. Resveratrol Promotes Myogenesis and Hypertrophy in Murine Myoblasts. J. Transl. Med. 2013, 11, 310. [Google Scholar] [CrossRef] [PubMed]
- Losso, J.N. The Maillard Reaction Reconsidered: Cooking and Eating for Health; CRC Press: London, UK, 2016; ISBN 9781482248227. [Google Scholar]
- Fu, Y.; Zhang, Y.; Soladoye, O.P.; Aluko, R.E. Maillard Reaction Products Derived from Food Protein-Derived Peptides: Insights into Flavor and Bioactivity. Crit. Rev. Food Sci. Nutr. 2019, 60, 3429–3442. [Google Scholar] [CrossRef]
- Yokoyama, I.; Setoyama, O.; Urakawa, A.; Sugawara, M.; Jia, Y.; Komiya, Y.; Nagasao, J.; Arihara, K. Lysine-Glucose Maillard Reaction Products Promote Longevity and Stress Tolerance in Caenorhabditis Elegans via the Insulin/IGF-1 Signaling Pathway. J. Funct. Foods 2021, 87, 104750. [Google Scholar] [CrossRef]
- Ajandouz, E.H.; Tchiakpe, L.S.; Ore, F.D.; Benajiba, A.; Puigserver, A. Effects of PH on Caramelization and Maillard Reaction Kinetics in Fructose-lysine Model Systems. J. Food Sci. 2001, 66, 926–931. [Google Scholar] [CrossRef]
- Watanabe, N.; Komiya, Y.; Sato, Y.; Watanabe, Y.; Suzuki, T.; Arihara, K. Oleic Acid Up-Regulates Myosin Heavy Chain (MyHC) 1 Expression and Increases Mitochondrial Mass and Maximum Respiration in C2C12 Myoblasts. Biochem. Biophys. Res. Commun. 2020, 525, 406–411. [Google Scholar] [CrossRef]
- Komiya, Y.; Sakazaki, Y.; Goto, T.; Kawabata, F.; Suzuki, T.; Sato, Y.; Sawano, S.; Nakamura, M.; Tatsumi, R.; Ikeuchi, Y.; et al. Eicosapentaenoic Acid Increases Proportion of Type 1 Muscle Fibers through PPARδ and AMPK Pathways in Rats. iScience 2024, 27, 109816. [Google Scholar] [CrossRef]
- Sawano, S.; Baba, K.; Sonoda, Y.; Wakamatsu, J.-I.; Tomonaga, S.; Furuse, M.; Sato, Y.; Tatsumi, R.; Ikeuchi, Y.; Mizunoya, W. Beef Extract Supplementation Promotes Myoblast Proliferation and Myotube Growth in C2C12 Cells. Eur. J. Nutr. 2020, 59, 3735–3743. [Google Scholar] [CrossRef]
- Sawano, S.; Kobayashi, Y.; Maesawa, S.; Mizunoya, W. Egg Components Reverse the Atrophy Induced by Injury in Skeletal Muscles. Genes Cells 2022, 27, 138–144. [Google Scholar] [CrossRef]
- Stitt, T.N.; Drujan, D.; Clarke, B.A.; Panaro, F.; Timofeyva, Y.; Kline, W.O.; Gonzalez, M.; Yancopoulos, G.D.; Glass, D.J. The IGF-1/PI3K/Akt Pathway Prevents Short Article Expression of Muscle Atrophy-Induced Ubiquitin Ligases by Inhibiting FOXO Transcription Factors. Mol. Cell 2004, 14, 395–403. [Google Scholar] [CrossRef] [PubMed]
- Morales, F.J.; Somoza, V.; Fogliano, V. Physiological Relevance of Dietary Melanoidins. Amino Acids 2010, 42, 1097–1109. [Google Scholar] [CrossRef] [PubMed]
- Powers, S.K.; Morton, A.B.; Ahn, B.; Smuder, A.J. Redox Control of Skeletal Muscle Atrophy. Free Radic. Biol. Med. 2016, 98, 208–217. [Google Scholar] [CrossRef] [PubMed]
- Damiano, S.; Muscariello, E.; La Rosa, G.; Di Maro, M.; Mondola, P.; Santillo, M. Dual Role of Reactive Oxygen Species in Muscle Function: Can Antioxidant Dietary Supplements Counteract Age-Related Sarcopenia? Int. J. Mol. Sci. 2019, 20, 3815. [Google Scholar] [CrossRef]
- Sies, H.; Berndt, C.; Jones, D.P. Oxidative Stress. Annu. Rev. Biochem. 2017, 86, 715–748. [Google Scholar] [CrossRef]
- Imlay, J.A. Cellular Defenses against Superoxide and Hydrogen Peroxide. Annu. Rev. Biochem. 2008, 77, 755–776. [Google Scholar] [CrossRef]
- Pisoschi, A.M.; Pop, A. The Role of Antioxidants in the Chemistry of Oxidative Stress: A Review. Eur. J. Med. Chem. 2015, 97, 55–74. [Google Scholar] [CrossRef]
- Liou, G.-Y.; Storz, P. Reactive Oxygen Species in Cancer. Free Radic. Res. 2010, 44, 479–496. [Google Scholar] [CrossRef]
- Barrera, G. Oxidative Stress and Lipid Peroxidation Products in Cancer Progression and Therapy. Int. Sch. Res. Not. 2012, 2012, 137289. [Google Scholar] [CrossRef]
- Kunsch, C.; Medford, R.M. Oxidative Stress as a Regulator of Gene Expression in the Vasculature. Circ. Res. 1999, 85, 753–766. [Google Scholar] [CrossRef]
- Bindoli, A.; Rigobello, M.P. Principles in Redox Signaling: From Chemistry to Functional Significance. Antioxid. Redox Signal. 2013, 18, 1557–1593. [Google Scholar] [CrossRef] [PubMed]
- Tan, P.L.; Shavlakadze, T.; Grounds, M.D.; Arthur, P.G. Differential Thiol Oxidation of the Signaling Proteins Akt, PTEN or PP2A Determines Whether Akt Phosphorylation Is Enhanced or Inhibited by Oxidative Stress in C2C12 Myotubes Derived from Skeletal Muscle. Int. J. Biochem. Cell Biol. 2015, 62, 72–79. [Google Scholar] [CrossRef] [PubMed]
- Ditch, S.; Paull, T.T. The ATM Protein Kinase and Cellular Redox Signaling: Beyond the DNA Damage Response. Trends Biochem. Sci. 2012, 37, 15–22. [Google Scholar] [CrossRef] [PubMed]
- Saha, S.; Buttari, B.; Panieri, E.; Profumo, E.; Saso, L. An Overview of Nrf2 Signaling Pathway and Its Role in Inflammation. Molecules 2020, 25, 5474. [Google Scholar] [CrossRef]
- Shelar, S.B.; Narasimhan, M.; Shanmugam, G.; Litovsky, S.H.; Gounder, S.S.; Karan, G.; Arulvasu, C.; Kensler, T.W.; Hoidal, J.R.; Darley-Usmar, V.M.; et al. Disruption of Nuclear Factor (Erythroid-Derived-2)-like 2 Antioxidant Signaling: A Mechanism for Impaired Activation of Stem Cells and Delayed Regeneration of Skeletal Muscle. FASEB J. 2016, 30, 1865–1879. [Google Scholar] [CrossRef]
- Hayashi, T.; Kudo, T.; Fujita, R.; Fujita, S.-I.; Tsubouchi, H.; Fuseya, S.; Suzuki, R.; Hamada, M.; Okada, R.; Muratani, M.; et al. Nuclear Factor E2-Related Factor 2 (NRF2) Deficiency Accelerates Fast Fibre Type Transition in Soleus Muscle during Space Flight. Commun. Biol. 2021, 4, 787. [Google Scholar] [CrossRef]
- Bodine, S.C.; Baehr, L.M. Skeletal Muscle Atrophy and the E3 Ubiquitin Ligases MuRF1 and MAFbx/Atrogin-1. Am. J. Physiol.-Endocrinol. Metab. 2014, 307, E469–E484. [Google Scholar] [CrossRef]
- Schakman, O.; Gilson, H.; Kalista, S.; Thissen, J.P. Mechanisms of Muscle Atrophy Induced by Glucocorticoids. Horm. Res. 2009, 72, 36–41. [Google Scholar] [CrossRef]
- Wang, D.-T.; Yin, Y.; Yang, Y.-J.; Lv, P.-J.; Shi, Y.; Lu, L.; Wei, L.-B. Resveratrol Prevents TNF-α-Induced Muscle Atrophy via Regulation of Akt/MTOR/FoxO1 Signaling in C2C12 Myotubes. Int. Immunopharmacol. 2014, 19, 206–213. [Google Scholar] [CrossRef]
- Bennett, B.T.; Mohamed, J.S.; Alway, S.E. Effects of Resveratrol on the Recovery of Muscle Mass Following Disuse in the Plantaris Muscle of Aged Rats. PLoS ONE 2013, 8, e83518. [Google Scholar] [CrossRef]
- Mukai, R.; Nakao, R.; Yamamoto, H.; Nikawa, T.; Takeda, E.; Terao, J. Quercetin Prevents Unloading-Derived Disused Muscle Atrophy by Attenuating the Induction of Ubiquitin Ligases in Tail-Suspension Mice. J. Nat. Prod. 2010, 73, 1708–1710. [Google Scholar] [CrossRef]
- Le, N.H.; Kim, C.-S.; Park, T.; Park, J.H.Y.; Sung, M.-K.; Lee, D.G.; Hong, S.-M.; Choe, S.-Y.; Goto, T.; Kawada, T.; et al. Quercetin Protects against Obesity-Induced Skeletal Muscle Inflammation and Atrophy. Mediat. Inflamm. 2014, 2014, 834294. [Google Scholar] [CrossRef]
- Vargas-Mendoza, N.; Madrigal-Santillán, E.; Álvarez-González, I.; Madrigal-Bujaidar, E.; Anguiano-Robledo, L.; Aguilar-Faisal, J.L.; Morales-Martínez, M.; Delgado-Olivares, L.; Rodríguez-Negrete, E.V.; Morales-González, Á.; et al. Phytochemicals in Skeletal Muscle Health: Effects of Curcumin (from Curcuma longa Linn) and Sulforaphane (from Brassicaceae) on Muscle Function, Recovery and Therapy of Muscle Atrophy. Plants 2022, 11, 2517. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Zheng, J.; Chen, H.; Li, X.; Ye, C.; Zhang, F.; Zhang, Z.; Yao, Q.; Guo, Y. Curcumin Targeting NF-ΚB/Ubiquitin-Proteasome-System Axis Ameliorates Muscle Atrophy in Triple-Negative Breast Cancer Cachexia Mice. Mediat. Inflamm. 2022, 2022, 2567150. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, S.; Hayashi, T.; Egawa, T. Advanced Glycation End Products Inhibit Proliferation and Primary Cilia Formation of Myoblasts through Receptor for Advanced Glycation End Products Pathway. Biochem. Biophys. Res. Commun. 2023, 684, 149141. [Google Scholar] [CrossRef] [PubMed]
- Egawa, T.; Tsuda, S.; Goto, A.; Ohno, Y.; Yokoyama, S.; Goto, K.; Hayashi, T. Potential Involvement of Dietary Advanced Glycation End Products in Impairment of Skeletal Muscle Growth and Muscle Contractile Function in Mice. Br. J. Nutr. 2017, 117, 21–29. [Google Scholar] [CrossRef]
- Suzuki, S.; Hayashi, T.; Egawa, T. Advanced Glycation End Products Promote ROS Production via PKC/P47 Phox Axis in Skeletal Muscle Cells. J. Physiol. Sci. 2024, 74, 51. [Google Scholar] [CrossRef]
- Kong, X.; Wang, G.-D.; Ma, M.-Z.; Deng, R.-Y.; Guo, L.-Q.; Zhang, J.-X.; Yang, J.-R.; Su, Q. Sesamin Ameliorates Advanced Glycation End Products-Induced Pancreatic β-Cell Dysfunction and Apoptosis. Nutrients 2015, 7, 4689–4704. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Dependent Variable | Measurement Instruments/Methods |
|---|---|
| Maillard reaction products | Reaction of 0.1 M L-lysine and D-glucose in 0.25% (w/v) sodium carbonate buffer (pH 11.0) |
| Average myotube diameter | Microscopy and ImageJ analysis; measured the diameters of 50 individual myotubes |
| Largest myotube diameter | Microscopy and ImageJ analysis; top 25 largest myotubes selected from 50 measured |
| Fusion index | Immunostaining with anti-MyHC antibody; percentage of myonuclei in MyHC-positive myotubes (≥2 nuclei) |
| Akt phosphorylation | Western blotting |
| mRNA expression | Quantitative real-time PCR |
| Antioxidant activity | DPPH radical scavenging assay; H2DCFDA staining |
| Gene | Forward (5′-3′) | Reverse (5′-3′) |
|---|---|---|
| Cat | CCTTCAAGTTGGTTAATGCAGA | CAAGTTTTTGATGCCCTGGT |
| Foxo1 | GTGGGGCAACCTGTCGTA | TTCTCGGCTGAGCTCTCG |
| Gclc | AGATGATAGAACACGGGAGGAG | TGATCCTAAAGCGATTGTTCTTC |
| Igf1 | GCAGTTCTAACACCAGCCCA | CCCACTCGATCGTACCTTCTG |
| Myh1 | TCGCTGG CTTTGAGATCTTT | CGAACATGTGGTGGTTGAAG |
| Myh2 | AAAGCTCCAAGGACCCTCTT | AGCTCATGACTGCTGAACTCAC |
| Myh4 | GTCACCAAAGGCCAGACG | ACATCTTCTCATACATGGACTTGG |
| Myh7 | GAGCAGCAGGTGGATGATCT | GCTTGGCTCGCTCTAGGTC |
| Myod1 | AGCACTACAGTGGCGACTCA | GGCCGCTGTAATCCATCAT |
| Myog | CCTTGCTCAGCTCCCTCA | TGGGAGTTGCATTCACTGG |
| Nfe2l2 | CATGATGGACTTGGAGTTGC | CCTCCAAAGGATGTCAATCAA |
| Nqo1 | AGCGTTCGGTATTACGATCC | AGTACAATCAGGGCTCTTCTCG |
| Tbp | GGGGAGCTGTGATGTGAAGT | CCAGGAAATAATTCTGGCTCAT |
| Trim63 | TGACATCTACAAGCAGGAGTGC | TCGTCTTCGTGTTCCTTGC |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Miyaki, M.; Komiya, Y.; Sumiya, I.; Yamaguchi, R.; Kuno, M.; Kojima, C.; Makino, R.; Suzuki, T.; Suzuki, Y.; Yokoyama, I.; et al. Effects of Maillard Reaction Products on Skeletal Muscle Cells: An In Vitro Study Using C2C12 Myotubes. Metabolites 2025, 15, 316. https://doi.org/10.3390/metabo15050316
Miyaki M, Komiya Y, Sumiya I, Yamaguchi R, Kuno M, Kojima C, Makino R, Suzuki T, Suzuki Y, Yokoyama I, et al. Effects of Maillard Reaction Products on Skeletal Muscle Cells: An In Vitro Study Using C2C12 Myotubes. Metabolites. 2025; 15(5):316. https://doi.org/10.3390/metabo15050316
Chicago/Turabian StyleMiyaki, Marina, Yusuke Komiya, Itsuki Sumiya, Rina Yamaguchi, Moeka Kuno, Chika Kojima, Ryosuke Makino, Takahiro Suzuki, Yoshihiro Suzuki, Issei Yokoyama, and et al. 2025. "Effects of Maillard Reaction Products on Skeletal Muscle Cells: An In Vitro Study Using C2C12 Myotubes" Metabolites 15, no. 5: 316. https://doi.org/10.3390/metabo15050316
APA StyleMiyaki, M., Komiya, Y., Sumiya, I., Yamaguchi, R., Kuno, M., Kojima, C., Makino, R., Suzuki, T., Suzuki, Y., Yokoyama, I., & Arihara, K. (2025). Effects of Maillard Reaction Products on Skeletal Muscle Cells: An In Vitro Study Using C2C12 Myotubes. Metabolites, 15(5), 316. https://doi.org/10.3390/metabo15050316