
Metabolomics Insights into Gut Microbiota and Functional Constipation

 , , ,
, , ,  and
and
Abstract
1. Introduction
1.1. Diagnostic Advancements
1.2. Personalized Therapeutic Interventions
1.3. Health Management of Constipation
2. The Relationship Between Gut Microbiota and Constipation
2.1. Normal Microbiota and Constipation Balance
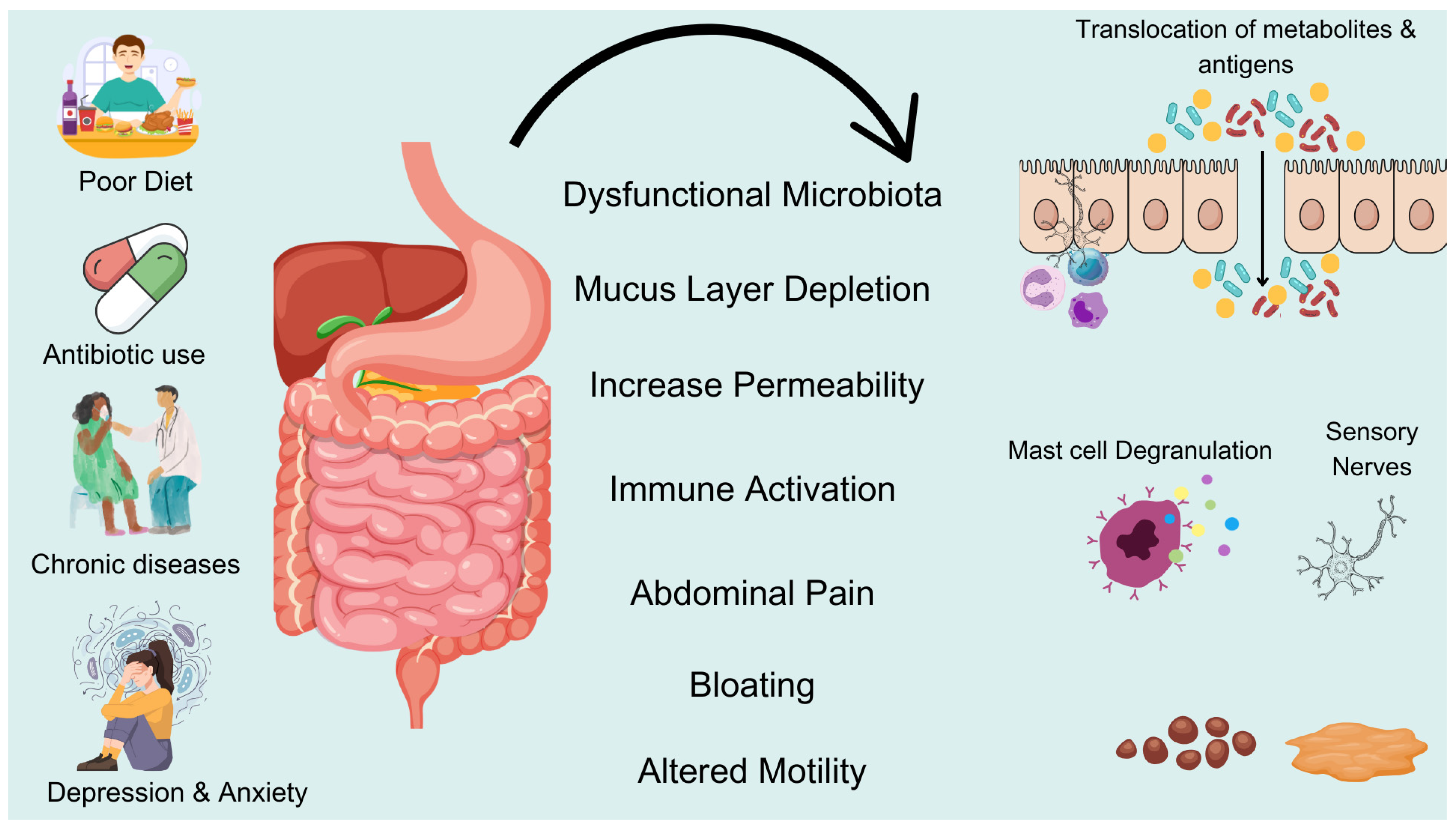
2.2. Mechanisms of Dysbiosis and the Occurrence of Constipation
2.3. Mechanisms by Which the Gut Microbiota Regulates Intestinal Motility and Functions
3. Exploring Metabolomics in Understanding Constipation Mechanisms
3.1. Metabolomic Characteristics of Patients with Constipation
3.2. The Impact of Gut Microbial Metabolites on Host Metabolism
3.3. Analysis of Potential Biomarkers of Constipation

{kind=link}
{kind=link}
| Population (n) | Intervention | Key Outcomes | Microbiota/Metabolite Changes | Reference |
|---|---|---|---|---|
| FC patients (n = 60) vs. Healthy (n = 60) | None (observational) | Butyrate (5.2 vs. 12.3 μM) correlated with slower transit (p < 0.01) | Roseburia decreased; Bacteroides increased | [11] |
| Adults with FC (n = 120) | Prebiotic UG1601 (8 weeks) | Stool frequency (3.1 increased to 5.2/week; p = 0.003); improved Bristol score | Bifidobacterium increased; fecal butyrate increased by 40% | [53] |
| FC patients (n = 45) | L. rhamnosus LRJ-1 (12 weeks) | Improved bowel movements (p = 0.02); reduced bloating | GABA-producing bacteria (Lactobacillus increased 2.5-fold) | [54] |
| Refractory FC (n = 30) | FMT (single dose) | 70% response rate (with increased stool frequency); effects lasted 6 months | Donor-like microbiota; SCFAs (butyrate) increased by 60% | [84] |
| Healthy adults (n = 40) | Arabinoxylan oligosaccharides | Improved transit time (p = 0.04); no adverse effects | Bifidobacterium; propionateincreased by 35% | [83] |
4. Mechanism of Gut Microbiota Metabolites and Constipation
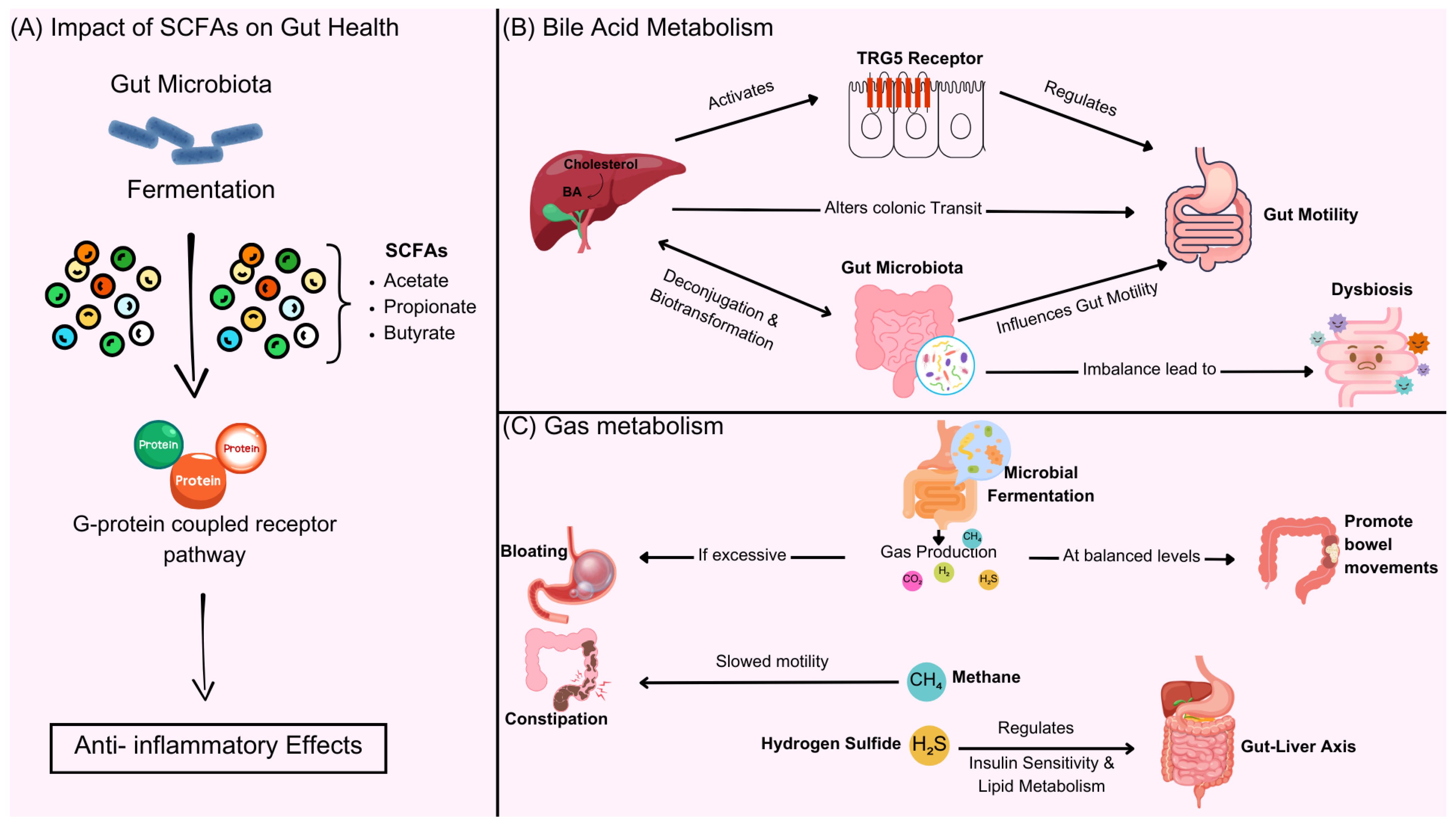
4.1. The Impact of SCFAs on Gut Health
4.2. BAs Metabolism and Intestinal Motility
4.3. Gas Metabolism and Regulation of Intestinal Function
5. The Potential Applications of Metabolomics in Personalized Treatment of Constipation
5.1. Innovative Approaches in Personalized Diagnosis of Constipation: Metabolomics-Driven Strategies
5.2. Metabolic Strategies for Treating Constipation by Regulating Gut Microbiota
5.3. Future Research Directions and Challenges
6. Concluding Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| VTE | Venous thromboembolism |
| BAs | Bile acids |
| SCFAs | Short-chain fatty acids |
| FC | Functional constipation |
| DCA | Deoxycholic acid |
| ENS | Enteric nervous system |
| CNS | Central nervous system |
| MMC | Migrating motor complex |
| NO | Nitric oxide |
| IBS | Irritable bowel syndrome |
| IBD | Inflammatory bowel disease |
| GC-MS | Gas chromatography–mass spectrometry |
| LC-MS | Liquid chromatography–mass spectrometry |
| NMR | nuclear magnetic resonance |
| GABA | Gamma-aminobutyric acid |
| XOS | Xylooligosaccharides |
| FMT | Fecal microbiota transplantation |
| GMIGH | Global Metabolomic Initiative for Gastrointestinal Health |
References
- Bharucha, A.E.; Lacy, B.E. Mechanisms, Evaluation, and Management of Chronic Constipation. Gastroenterology 2020, 158, 1232–1249. [Google Scholar] [CrossRef]
- Ford, A.C.; Moayyedi, P.; Lacy, B.E.; Lembo, A.J.; Saito, Y.A.; Schiller, L.R.; Soffer, E.E.; Spiegel, B.M.; Quigley, E.M.; FACG for the Task Force on the Management of Functional Bowel Disorders. American College of Gastroenterology monograph on the management of irritable bowel syndrome and chronic idiopathic constipation. Am. J. Gastroenterol. 2014, 109 (Suppl. S1), S2–S26, quiz S27. [Google Scholar] [CrossRef] [PubMed]
- Bytzer, P.; Howell, S.; Leemon, M.; Young, L.J.; Jones, M.P.; Talley, N.J. Low socioeconomic class is a risk factor for upper and lower gastrointestinal symptoms: A population based study in 15 000 Australian adults. Gut 2001, 49, 66–72. [Google Scholar] [CrossRef]
- Wald, A.; Scarpignato, C.; Kamm, M.A.; Mueller-Lissner, S.; Helfrich, I.; Schuijt, C.; Bubeck, J.; Limoni, C.; Petrini, O. The burden of constipation on quality of life: Results of a multinational survey. Aliment. Pharmacol. Ther. 2007, 26, 227–236. [Google Scholar] [CrossRef]
- Suares, N.C.; Ford, A.C. Prevalence of, and risk factors for, chronic idiopathic constipation in the community: Systematic review and meta-analysis. Am. J. Gastroenterol. 2011, 106, 1582–1591, quiz 1581, 1592. [Google Scholar] [CrossRef] [PubMed]
- Vazquez Roque, M.; Bouras, E.P. Epidemiology and management of chronic constipation in elderly patients. Clin. Interv. Aging 2015, 10, 919–930. [Google Scholar] [CrossRef] [PubMed]
- Talley, N.J.; Fleming, K.C.; Evans, J.M.; O‘Keefe, E.A.; Weaver, A.L.; Zinsmeister, A.R.; Melton, L.J., 3rd. Constipation in an elderly community: A study of prevalence and potential risk factors. Am. J. Gastroenterol. 1996, 91, 19–25. [Google Scholar]
- Talley, N.J.; O‘Keefe, E.A.; Zinsmeister, A.R.; Melton, L.J., 3rd. Prevalence of gastrointestinal symptoms in the elderly: A population-based study. Gastroenterology 1992, 102, 895–901. [Google Scholar] [CrossRef]
- Shin, A.; Ballou, S.; Camilleri, M.; Xu, H.; Lembo, A. Information- and Health-care Seeking Behaviors in Patients with Irritable Bowel Syndrome. Clin. Gastroenterol. Hepatol. 2020, 18, 2840–2842. [Google Scholar] [CrossRef]
- McCormick, D. Managing costs and care for chronic idiopathic constipation. Am. J. Manag. Care. 2019, 25, S63–S69. [Google Scholar]
- Min, L.; Chi, Y.; Dong, S. Gut microbiota health closely associates with PCB153-derived risk of host diseases. Ecotoxicol. Environ. Saf. 2020, 203, 111041. [Google Scholar] [CrossRef]
- Chi, L.; Tu, P.; Ru, H.; Lu, K. Studies of xenobiotic-induced gut microbiota dysbiosis: From correlation to mechanisms. Gut Microbes 2021, 13, 1921912. [Google Scholar] [CrossRef]
- Pavlo, P.; Kamyshna, I.; Kamyshnyi, A. Effects of metformin on the gut microbiota: A systematic review. Mol. Metab. 2023, 77, 101805. [Google Scholar] [CrossRef] [PubMed]
- Xiong, R.G.; Li, J.; Cheng, J.; Zhou, D.D.; Wu, S.X.; Huang, S.Y.; Saimaiti, A.; Yang, Z.J.; Gan, R.Y.; Li, H.B. The Role of Gut Microbiota in Anxiety, Depression, and Other Mental Disorders as Well as the Protective Effects of Dietary Components. Nutrients 2023, 15, 3258. [Google Scholar] [CrossRef]
- Serra, J.; Mascort-Roca, J.; Marzo-Castillejo, M.; Delgado Aros, S.; Ferrandiz Santos, J.; Rey Diaz Rubio, E.; Mearin Manrique, F. Clinical practice guidelines for the management of constipation in adults. Part 1: Definition, aetiology and clinical manifestations. Gastroenterol. Hepatol. 2017, 40, 132–141. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.; Rao, S. Constipation: Pathophysiology and Current Therapeutic Approaches. Handb. Exp. Pharmacol. 2017, 239, 59–74. [Google Scholar] [CrossRef] [PubMed]
- Chen, B.; Zeng, G.; Sun, L.; Jiang, C. When smoke meets gut: Deciphering the interactions between tobacco smoking and gut microbiota in disease development. Sci. China Life Sci. 2024, 67, 854–864. [Google Scholar] [CrossRef]
- Cheung, P.K.; Ma, M.H.; Tse, H.F.; Yeung, K.F.; Tsang, H.F.; Chu, M.K.M.; Kan, C.M.; Cho, W.C.S.; Ng, L.B.W.; Chan, L.W.C.; et al. The applications of metabolomics in the molecular diagnostics of cancer. Expert Rev. Mol. Diagn. 2019, 19, 785–793. [Google Scholar] [CrossRef]
- Wang, J.; Wang, L.; Yu, Q.; Tang, N.; Mei, C.; Zhang, H.; Wang, G.; Lu, J.; Chen, W. Characteristics of the Gut Microbiome and Serum Metabolome in Patients with Functional Constipation. Nutrients 2023, 15, 1779. [Google Scholar] [CrossRef]
- Amdanee, N.; Shao, M.; Hu, X.; Fang, X.; Zhou, C.; Chen, J.; Ridwan Chattun, M.; Wen, L.; Pan, X.; Zhang, X.; et al. Serum Metabolic Profile in Schizophrenia Patients with Antipsychotic-Induced Constipation and Its relationship with Gut Microbiome. Schizophr. Bull. 2023, 49, 646–658. [Google Scholar] [CrossRef]
- Pan, R.; Wang, L.; Xu, X.; Chen, Y.; Wang, H.; Wang, G.; Zhao, J.; Chen, W. Crosstalk between the Gut Microbiome and Colonic Motility in Chronic Constipation: Potential Mechanisms and Microbiota Modulation. Nutrients 2022, 14, 3704. [Google Scholar] [CrossRef] [PubMed]
- Avelar Rodriguez, D.; Popov, J.; Ratcliffe, E.M.; Toro Monjaraz, E.M. Functional Constipation and the Gut Microbiome in Children: Preclinical and Clinical Evidence. Front. Pediatr. 2020, 8, 595531. [Google Scholar] [CrossRef] [PubMed]
- Martin, F.P.; Cominetti, O.; Berger, B.; Combremont, S.; Marquis, J.; Xie, G.; Jia, W.; Pinto-Sanchez, M.I.; Bercik, P.; Bergonzelli, G. Metabolome-associated psychological comorbidities improvement in irritable bowel syndrome patients receiving a probiotic. Gut Microbes 2024, 16, 2347715. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Wang, Y.; Zhang, Y.; Li, W.; Jiang, S.; Qian, D.; Duan, J. Gut microbiota: A new avenue to reveal pathological mechanisms of constipation. Appl. Microbiol. Biotechnol. 2022, 106, 6899–6913. [Google Scholar] [CrossRef]
- Zhang, X.; Hu, B.; Sun, G.; Zheng, J.; Hu, H.; Yang, H.; Cheng, X.; Lin, A.; Liu, H. Plasma metabolomic pro fi les reveal regulatory effect of chitosan oligosaccharides on loperamide-induced constipation in mice. J. Pharm. Biomed. Anal. 2022, 211, 114590. [Google Scholar] [CrossRef]
- Szachta, P.; Bartnicka, A.; Galecka, M. Microbiota—A key to healing the gastrointestinal tract? Pomeranian J. Life Sci. 2016, 62, 21–24. [Google Scholar]
- Goya-Jorge, E.; Bondue, P.; Gonza, I.; Laforet, F.; Antoine, C.; Boutaleb, S.; Douny, C.; Scippo, M.L.; de Ribaucourt, J.C.; Crahay, F.; et al. Butyrogenic, bifidogenic and slight anti-inflammatory effects of a green kiwifruit powder (Kiwi FFG(R)) in a human gastrointestinal model simulating mild constipation. Food Res. Int. 2023, 173, 113348. [Google Scholar] [CrossRef]
- Langford, J.S.; Tokita, E.; Martindale, C.; Millsap, L.; Hemp, J.; Pace, L.A.; Cortez, M.M. Quantitative gastrointestinal function and corresponding symptom profiles in autonomic neuropathy. Front. Neurol. 2022, 13, 1027348. [Google Scholar] [CrossRef]
- Saad, R.J. The Wireless Motility Capsule: A One-Stop Shop for the Evaluation of GI Motility Disorders. Curr. Gastroenterol. Rep. 2016, 18, 14. [Google Scholar] [CrossRef]
- Dimidi, E.; Christodoulides, S.; Fragkos, K.C.; Scott, S.M.; Whelan, K. The effect of probiotics on functional constipation in adults: A systematic review and meta-analysis of randomized controlled trials. Am. J. Clin. Nutr. 2014, 100, 1075–1084. [Google Scholar] [CrossRef]
- Zhang, C.; Jiang, J.; Tian, F.; Zhao, J.; Zhang, H.; Zhai, Q.; Chen, W. Meta-analysis of randomized controlled trials of the effects of probiotics on functional constipation in adults. Clin. Nutr. 2020, 39, 2960–2969. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.C.; Zeng, B.S.; Liang, C.S.; Hsu, C.W.; Su, K.P.; Wu, Y.C.; Tu, Y.K.; Lin, P.Y.; Stubbs, B.; Chen, T.Y.; et al. Efficacy and acceptability of different probiotic products plus laxatives for pediatric functional constipation: A network meta-analysis of randomized controlled trials. Eur. J. Pediatr. 2024, 183, 3531–3541. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Zhu, S.; Liu, Z.; Wei, H.; Zhang, L.; He, M.; Pei, F.; Zhang, J.; Sun, Q.; Duan, L. Increased Expression of Colonic Mucosal Melatonin in Patients with Irritable Bowel Syndrome Correlated with Gut Dysbiosis. Genom. Proteom. Bioinform. 2020, 18, 708–720. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Lv, C.; Yin, M.; Zhu, M.; Wang, B.; Tian, J.; Hashimoto, K.; Yu, Y. Efficacy and Safety of Transcutaneous Auricular Vagus Nerve Stimulation in Patients with Constipation-Predominant Irritable Bowel Syndrome: A Single-Center, Single-Blind, Randomized Controlled Trial. Am. J. Gastroenterol. 2024. [Google Scholar] [CrossRef]
- Kim, M.C.; Lee, S.; Park, J.K.; Park, J.; Lee, D.; Park, J.; Kim, B.Y.; Cho, M.S.; Kim, T.Y.; Park, H.Y.; et al. Effects of ID-HWS1000 on the Perception of Bowel Activity and Microbiome in Subjects with Functional Constipation: A Randomized, Double-Blind Placebo-Controlled Study. J. Med. Food 2021, 24, 883–893. [Google Scholar] [CrossRef]
- Storz, M.A.; Rizzo, G.; Muller, A.; Lombardo, M. Bowel Health in U.S. Vegetarians: A 4-Year Data Report from the National Health and Nutrition Examination Survey (NHANES). Nutrients 2022, 14, 681. [Google Scholar] [CrossRef]
- van Wunnik, B.P.; Baeten, C.G.; Southwell, B.R. Neuromodulation for constipation: Sacral and transcutaneous stimulation. Best Pract. Res. Clin. Gastroenterol. 2011, 25, 181–191. [Google Scholar] [CrossRef]
- Sharma, A.; Liu, B.; Waudby, P.; Duthie, G.S. Sacral neuromodulation for the management of severe constipation: Development of a constipation treatment protocol. Int. J. Colorectal. Dis. 2011, 26, 1583–1587. [Google Scholar] [CrossRef]
- Pitocco, D.; Di Leo, M.; Tartaglione, L.; De Leva, F.; Petruzziello, C.; Saviano, A.; Pontecorvi, A.; Ojetti, V. The role of gut microbiota in mediating obesity and diabetes mellitus. Eur. Rev. Med. Pharmacol. Sci. 2020, 24, 1548–1562. [Google Scholar] [CrossRef]
- Farcas, R.A.; Grad, S.; Grad, C.; Dumitrascu, D.L. Microbiota and Digestive Metabolites Alterations in Functional Dyspepsia. J. Gastrointestin. Liver Dis. 2024, 33, 102–106. [Google Scholar] [CrossRef]
- Hu, R.; Liu, Z.; Geng, Y.; Huang, Y.; Li, F.; Dong, H.; Ma, W.; Song, K.; Zhang, M.; Song, Y. Gut Microbiota and Critical Metabolites: Potential Target in Preventing Gestational Diabetes Mellitus? Microorganisms 2023, 11, 1725. [Google Scholar] [CrossRef]
- Mann, E.R.; Lam, Y.K.; Uhlig, H.H. Short-chain fatty acids: Linking diet, the microbiome and immunity. Nat. Rev. Immunol. 2024, 24, 577–595. [Google Scholar] [CrossRef]
- Xu, Z.; Liu, T.; Zhou, Q.; Chen, J.; Yuan, J.; Yang, Z. Roles of Chinese Medicine and Gut Microbiota in Chronic Constipation. Evid.-Based Complement. Altern. Med. 2019, 2019, 9372563. [Google Scholar] [CrossRef]
- Cao, H.; Liu, X.; An, Y.; Zhou, G.; Liu, Y.; Xu, M.; Dong, W.; Wang, S.; Yan, F.; Jiang, K.; et al. Dysbiosis contributes to chronic constipation development via regulation of serotonin transporter in the intestine. Sci. Rep. 2017, 7, 10322. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.X.; Jin, L.; Jin, H.; Zheng, G.S. Influence of Dietary Components and Traditional Chinese Medicine on Hypertension: A Potential Role for Gut Microbiota. Evid.-Based Complement. Altern. Med. 2021, 2021, 5563073. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.K.; Yao, S.K. Roles of Gut Microbiota and Metabolites in Pathogenesis of Functional Constipation. Evid.-Based Complement. Altern. Med. 2021, 2021, 5560310. [Google Scholar] [CrossRef] [PubMed]
- Tan, J.K.; Macia, L.; Mackay, C.R. Dietary fiber and SCFAs in the regulation of mucosal immunity. J. Allergy Clin. Immun. 2023, 151, 361–370. [Google Scholar] [CrossRef]
- Xu, Y.S.; Wang, Y.H.; Liu, Y.; Sun, X.; Xu, J.S.; Song, Y.; Jiang, X.; Xiong, Z.F.; Tian, Z.B.; Zhang, C.P. Alteration of the faecal microbiota composition in patients with constipation: Evidence of American Gut Project. Benef. Microbes 2022, 13, 427–436. [Google Scholar] [CrossRef]
- Zhu, L.; Liu, W.; Alkhouri, R.; Baker, R.D.; Bard, J.E.; Quigley, E.M.; Baker, S.S. Structural changes in the gut microbiome of constipated patients. Physiol. Genomics 2014, 46, 679–686. [Google Scholar] [CrossRef]
- Zhou, J.; Yuan, X.; Liu, Y. The gut microbiota-constipation connection: Insights from a two sample bidirectional Mendelian randomization study. Microb. Pathog. 2024, 192, 106667. [Google Scholar] [CrossRef]
- Bai, J.; Cai, Y.; Huang, Z.; Gu, Y.; Huang, N.; Sun, R.; Zhang, G.; Liu, R. Shouhui Tongbian Capsule ameliorates constipation via gut microbiota-5-HT-intestinal motility axis. Biomed. Pharmacother. 2022, 154, 113627. [Google Scholar] [CrossRef] [PubMed]
- Ohkusa, T.; Koido, S.; Nishikawa, Y.; Sato, N. Gut Microbiota and Chronic Constipation: A Review and Update. Front. Med. 2019, 6, 19. [Google Scholar] [CrossRef]
- Luo, M.; Xie, P.; Deng, X.; Fan, J.; Xiong, L. Rifaximin Ameliorates Loperamide-Induced Constipation in Rats through the Regulation of Gut Microbiota and Serum Metabolites. Nutrients 2023, 15, 4502. [Google Scholar] [CrossRef]
- Zhou, L.; Zhang, W.; Hu, X.; Wang, D.; Tang, D. Metabolic Reprogramming of Cancer-Associated Fibroblast in the Tumor Microenvironment: From Basics to Clinic. Clin. Med. Insights Oncol. 2024, 18, 11795549241287058. [Google Scholar] [CrossRef]
- Avetisyan, M.; Schill, E.M.; Heuckeroth, R.O. Building a second brain in the bowel. J. Clin. Investig. 2015, 125, 899–907. [Google Scholar] [CrossRef] [PubMed]
- Furness, J.B. The enteric nervous system and neurogastroenterology. Nat. Rev. Gastroenterol. Hepatol. 2012, 9, 286–294. [Google Scholar] [CrossRef]
- Tiwari, P.; Dwivedi, R.; Bansal, M.; Tripathi, M.; Dada, R. Role of Gut Microbiota in Neurological Disorders and Its Therapeutic Significance. J. Clin. Med. 2023, 12, 1650. [Google Scholar] [CrossRef] [PubMed]
- Deloose, E.; Janssen, P.; Depoortere, I.; Tack, J. The migrating motor complex: Control mechanisms and its role in health and disease. Nat. Rev. Gastroenterol. Hepatol. 2012, 9, 271–285. [Google Scholar] [CrossRef]
- Zhao, L.; Lu, W. Defensins in innate immunity. Curr. Opin. Hematol. 2014, 21, 37–42. [Google Scholar] [CrossRef]
- Porrini, C.; Ramarao, N.; Tran, S.L. Dr. NO and Mr. Toxic—The versatile role of nitric oxide. Biol. Chem. 2020, 401, 547–572. [Google Scholar] [CrossRef]
- Leclerc, M.; Bedu-Ferrari, C.; Etienne-Mesmin, L.; Mariadassou, M.; Lebreuilly, L.; Tran, S.L.; Brazeau, L.; Mayeur, C.; Delmas, J.; Rue, O.; et al. Nitric Oxide Impacts Human Gut Microbiota Diversity and Functionalities. mSystems 2021, 6, e0055821. [Google Scholar] [CrossRef] [PubMed]
- Fu, Y.J.; Zhao, X.; Wang, L.Y.; Li, K.; Jiang, N.; Zhang, S.T.; Wang, R.K.; Zhao, Y.F.; Yang, W. A Gas Therapy Strategy for Intestinal Flora Regulation and Colitis Treatment by Nanogel-Based Multistage NO Delivery Microcapsules. Adv. Mater. 2024, 36, e2309972. [Google Scholar] [CrossRef] [PubMed]
- Ejtahed, H.S.; Mohtadi-Nia, J.; Homayouni-Rad, A.; Niafar, M.; Asghari-Jafarabadi, M.; Mofid, V. Probiotic yogurt improves antioxidant status in type 2 diabetic patients. Nutrition 2012, 28, 539–543. [Google Scholar] [CrossRef] [PubMed]
- Ren, Z.; Zhao, Z.; Wang, Y.; Huang, K. Preparation of selenium/zinc-enriched probiotics and their effect on blood selenium and zinc concentrations, antioxidant capacities, and intestinal microflora in canine. Biol. Trace Elem. Res. 2011, 141, 170–183. [Google Scholar] [CrossRef]
- Drommi, F.; Calabro, A.; Vento, G.; Pezzino, G.; Cavaliere, R.; Omero, F.; Muscolino, P.; Granata, B.; D‘Anna, F.; Silvestris, N.; et al. Crosstalk between ILC3s and Microbiota: Implications for Colon Cancer Development and Treatment with Immune Check Point Inhibitors. Cancers 2023, 15, 2893. [Google Scholar] [CrossRef]
- Chang, W.Y.; Yang, Y.T.; She, M.P.; Tu, C.H.; Lee, T.C.; Wu, M.S.; Sun, C.H.; Hsin, L.W.; Yu, L.C.H. 5-HT7 receptor-dependent intestinal neurite outgrowth contributes to visceral hypersensitivity in irritable bowel syndrome. Lab. Investig. 2022, 102, 1023–1037. [Google Scholar] [CrossRef]
- Michetti, F.; Clementi, M.E.; Di Liddo, R.; Valeriani, F.; Ria, F.; Rende, M.; Di Sante, G.; Romano Spica, V. The S100B Protein: A Multifaceted Pathogenic Factor More Than a Biomarker. Int. J. Mol. Sci. 2023, 24, 9605. [Google Scholar] [CrossRef]
- Furlani, I.L.; da Cruz Nunes, E.; Canuto, G.A.B.; Macedo, A.N.; Oliveira, R.V. Liquid Chromatography-Mass Spectrometry for Clinical Metabolomics: An Overview. Adv. Exp. Med. Biol. 2021, 1336, 179–213. [Google Scholar] [CrossRef]
- Zhong, A.B.; Muti, I.H.; Eyles, S.J.; Vachet, R.W.; Sikora, K.N.; Bobst, C.E.; Calligaris, D.; Stopka, S.A.; Agar, J.N.; Wu, C.L.; et al. Multiplatform Metabolomics Studies of Human Cancers with NMR and Mass Spectrometry Imaging. Front. Mol. Biosci. 2022, 9, 785232. [Google Scholar] [CrossRef]
- El-Sayed, A.; Aleya, L.; Kamel, M. Microbiota’s role in health and diseases. Environ. Sci. Pollut. Res. Int. 2021, 28, 36967–36983. [Google Scholar] [CrossRef]
- Kassaian, N.; Feizi, A.; Rostami, S.; Aminorroaya, A.; Yaran, M.; Amini, M. The effects of 6 mo of supplementation with probiotics and synbiotics on gut microbiota in the adults with prediabetes: A double blind randomized clinical trial. Nutrition 2020, 79–80, 110854. [Google Scholar] [CrossRef]
- Caricilli, A.M.; Saad, M.J. The role of gut microbiota on insulin resistance. Nutrients 2013, 5, 829–851. [Google Scholar] [CrossRef] [PubMed]
- Dalile, B.; Van Oudenhove, L.; Vervliet, B.; Verbeke, K. The role of short-chain fatty acids in microbiota-gut-brain communication. Nat. Rev. Gastroenterol. Hepatol. 2019, 16, 461–478. [Google Scholar] [CrossRef]
- Wang, Z.Q.; Shi, Y.L.; Zeng, S.Y.; Zheng, Y.P.; Wang, H.J.; Liao, H.H.; Song, J.; Zhang, X.Y.; Cao, J.; Li, C. Polysaccharides from Relieve Loperamide-Induced Constipation Symptoms in Mice. Int. J. Mol. Sci. 2023, 24, 2553. [Google Scholar] [CrossRef]
- Zhuang, M.; Shang, W.; Ma, Q.; Strappe, P.; Zhou, Z. Abundance of Probiotics and Butyrate-Production Microbiome Manages Constipation via Short-Chain Fatty Acids Production and Hormones Secretion. Mol. Nutr. Food Res. 2019, 63, e1801187. [Google Scholar] [CrossRef] [PubMed]
- Chu, J.R.; Kang, S.Y.; Kim, S.E.; Lee, S.J.; Lee, Y.C.; Sung, M.K. Prebiotic UG1601 mitigates constipation-related events in association with gut microbiota: A randomized placebo-controlled intervention study. World J. Gastroenterol. 2019, 25, 6129–6144. [Google Scholar] [CrossRef]
- Xia, T.; Huang, F.; Yun, F.; Liu, Y.; Wang, T.; Wang, S.; Jin, S.; Ma, X.; Wang, W.; He, J.; et al. Lacticaseibacillus rhamnosus LRJ-1 alleviates constipation through promoting gut Bacteroides-derived gamma-aminobutyric acid production. Curr. Res. Food Sci. 2024, 9, 100924. [Google Scholar] [CrossRef]
- Yang, C.; Hu, T.; Xue, X.; Su, X.; Zhang, X.; Fan, Y.; Shen, X.; Dong, X. Multi-omics analysis of fecal microbiota transplantation’s impact on functional constipation and comorbid depression and anxiety. BMC Microbiol. 2023, 23, 389. [Google Scholar] [CrossRef]
- Liu, P.; Jin, M.; Hu, P.; Sun, W.; Tang, Y.; Wu, J.; Zhang, D.; Yang, L.; He, H.; Xu, X. Gut microbiota and bile acids: Metabolic interactions and impacts on diabetic kidney disease. Curr. Res. Microb. Sci. 2024, 7, 100315. [Google Scholar] [CrossRef]
- Koh, A.; De Vadder, F.; Kovatcheva-Datchary, P.; Backhed, F. From Dietary Fiber to Host Physiology: Short-Chain Fatty Acids as Key Bacterial Metabolites. Cell 2016, 165, 1332–1345. [Google Scholar] [CrossRef]
- Wang, H.; Zhao, D.; Wang, S.; Liu, H.; Zhao, S.; Li, Z.; Qin, X.; Liu, X. Gastrointestinal Characteristics of Constipation from the Perspectives of Microbiome and Metabolome. Dig. Dis. Sci. 2024, 69, 1318–1335. [Google Scholar] [CrossRef]
- Fraser, K.; James, S.C.; Young, W.; Gearry, R.B.; Heenan, P.E.; Keenan, J.I.; Talley, N.J.; McNabb, W.C.; Roy, N.C. Characterisation of the Plasma and Faecal Metabolomes in Participants with Functional Gastrointestinal Disorders. Int. J. Mol. Sci. 2024, 25, 13465. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Lin, Z.; Lin, L.; Wang, M.; Zhang, H. Functional constipation: Implications for nursing interventions. J. Clin. Nurs. 2010, 19, 1838–1843. [Google Scholar] [CrossRef]
- Karasawa, Y.; Nozawa, K.; Nomoto, K.; Fukudo, S. Constipation among workers with depression/anxiety: A retrospective study using a claims database and survey data in Japan. BMJ Open 2024, 14, e083668. [Google Scholar] [CrossRef]
- Liang, J.; Zhao, Y.; Xi, Y.; Xiang, C.; Yong, C.; Huo, J.; Zou, H.; Hou, Y.; Pan, Y.; Wu, M.; et al. Association between Depression, Anxiety Symptoms and Gut Microbiota in Chinese Elderly with Functional Constipation. Nutrients 2022, 14, 5013. [Google Scholar] [CrossRef]
- Tian, D.; Zhang, W.; Lu, L.; Yu, Y.; Yu, Y.; Zhang, X.; Li, W.; Shi, W.; Liu, G. Enrofloxacin exposure undermines gut health and disrupts neurotransmitters along the microbiota-gut-brain axis in zebrafish. Chemosphere 2024, 356, 141971. [Google Scholar] [CrossRef] [PubMed]
- Hu, W.; Shang, R.; Yang, J.; Chen, C.; Liu, Z.; Liang, G.; He, W.; Luo, G. Skin γδ T Cells and Their Function in Wound Healing. Front. Immunol. 2022, 13, 875076. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Sun, X.; Li, Z.; Zhao, R.; Li, M.; Hu, T. Machine learning-based integration develops biomarkers initial the crosstalk between inflammation and immune in acute myocardial infarction patients. Front. Cardiovasc. Med. 2022, 9, 1059543. [Google Scholar] [CrossRef]
- Song, H.; Guo, R.; Sun, X.; Kou, Y.; Ma, X.; Chen, Y.; Song, L.; Wu, Y. Integrated metabolomics and transcriptomics revealed the anti-constipation mechanisms of xylooligosaccharides from corn cobs. Food Funct. 2024, 15, 894–905. [Google Scholar] [CrossRef]
- Chen, Y.; Wu, T.; Lu, W.; Yuan, W.; Pan, M.; Lee, Y.K.; Zhao, J.; Zhang, H.; Chen, W.; Zhu, J.; et al. Predicting the Role of the Human Gut Microbiome in Constipation Using Machine-Learning Methods: A Meta-Analysis. Microorganisms 2021, 9, 2149. [Google Scholar] [CrossRef]
- Devkota, S.; Chang, E.B. Interactions between Diet, Bile Acid Metabolism, Gut Microbiota, and Inflammatory Bowel Diseases. Dig. Dis. 2015, 33, 351–356. [Google Scholar] [CrossRef] [PubMed]
- Silva, Y.P.; Bernardi, A.; Frozza, R.L. The Role of Short-Chain Fatty Acids from Gut Microbiota in Gut-Brain Communication. Front. Endocrinol. 2020, 11, 25. [Google Scholar] [CrossRef] [PubMed]
- Xiao, L.; Liu, Q.; Luo, M.; Xiong, L. Gut Microbiota-Derived Metabolites in Irritable Bowel Syndrome. Front. Cell. Infect. Microbiol. 2021, 11, 729346. [Google Scholar] [CrossRef]
- Jin, K.; Yu, Y.; Zeng, H.; Liu, Z.; You, R.; Zhang, H.; Liu, C.; Su, X.; Yan, S.; Chang, Y.; et al. CD103+CD8+ tissue-resident memory T cell infiltration predicts clinical outcome and adjuvant therapeutic benefit in muscle-invasive bladder cancer. Br. J. Cancer 2022, 126, 1581–1588. [Google Scholar] [CrossRef]
- Park, S.; Sadanala, K.C.; Kim, E.K. A Metabolomic Approach to Understanding the Metabolic Link between Obesity and Diabetes. Mol. Cells 2015, 38, 587–596. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Liu, C.; Huang, S.; Wang, X.; Cao, M.; Gu, T.; Ou, X.; Pan, S.; Lin, Z.; Wang, X.; et al. Multi-omics analyses demonstrate the modulating role of gut microbiota on the associations of unbalanced dietary intake with gastrointestinal symptoms in children with autism spectrum disorder. Gut Microbes 2023, 15, 2281350. [Google Scholar] [CrossRef]
- Zhang, S.; Wang, R.; Li, D.; Zhao, L.; Zhu, L. Role of gut microbiota in functional constipation. Gastroenterol. Rep. 2021, 9, 392–401. [Google Scholar] [CrossRef]
- Yang, C.; Bai, X.; Hu, T.; Xue, X.; Su, X.; Zhang, X.; Wu, T.; Zhang, M.; Shen, X.; Dong, X. Integrated metagenomics and targeted-metabolomics analysis of the effects of phenylalanine on loperamide-induced constipation in rats. Front. Microbiol. 2022, 13, 1018008. [Google Scholar] [CrossRef]
- Serra, J. Management of bloating. Neurogastroenterol. Motil. 2022, 34, e14333. [Google Scholar] [CrossRef]
- Linlawan, S.; Patcharatrakul, T.; Somlaw, N.; Gonlachanvit, S. Effect of Rice, Wheat, and Mung Bean Ingestion on Intestinal Gas Production and Postprandial Gastrointestinal Symptoms in Non-Constipation Irritable Bowel Syndrome Patients. Nutrients 2019, 11, 2061. [Google Scholar] [CrossRef]
- Farre, R.; Fiorani, M.; Abdu Rahiman, S.; Matteoli, G. Intestinal Permeability, Inflammation and the Role of Nutrients. Nutrients 2020, 12, 1185. [Google Scholar] [CrossRef] [PubMed]
- Triantafyllou, K.; Chang, C.; Pimentel, M. Methanogens, methane and gastrointestinal motility. J. Neurogastroenterol. Motil. 2014, 20, 31–40. [Google Scholar] [CrossRef] [PubMed]
- Cai, L.; Wang, X.; Zhu, X.; Xu, Y.; Qin, W.; Ren, J.; Jiang, Q.; Yan, X. Lactobacillus-derived protoporphyrin IX and SCFAs regulate the fiber size via glucose metabolism in the skeletal muscle of chickens. mSystems 2024, 9, e0021424. [Google Scholar] [CrossRef] [PubMed]
- Borewicz, K.; Zhao, Y.; Zhu, Y. Daily intake of a dairy-based nutritional supplement improved self-reported gastrointestinal symptoms and modulated microbiota in adult Chinese volunteers. Sci. Rep. 2024, 14, 28651. [Google Scholar] [CrossRef]
- Rahayu, E.S.; Mariyatun, M.; Putri Manurung, N.E.; Hasan, P.N.; Therdtatha, P.; Mishima, R.; Komalasari, H.; Mahfuzah, N.A.; Pamungkaningtyas, F.H.; Yoga, W.K.; et al. Effect of probiotic Lactobacillus plantarum Dad-13 powder consumption on the gut microbiota and intestinal health of overweight adults. World J. Gastroenterol. 2021, 27, 107–128. [Google Scholar] [CrossRef]
- Francois, I.E.; Lescroart, O.; Veraverbeke, W.S.; Marzorati, M.; Possemiers, S.; Evenepoel, P.; Hamer, H.; Houben, E.; Windey, K.; Welling, G.W.; et al. Effects of a wheat bran extract containing arabinoxylan oligosaccharides on gastrointestinal health parameters in healthy adult human volunteers: A double-blind, randomised, placebo-controlled, cross-over trial. Br. J. Nutr. 2012, 108, 2229–2242. [Google Scholar] [CrossRef]
- Tian, Y.; Zuo, L.; Guo, Q.; Li, J.; Hu, Z.; Zhao, K.; Li, C.; Li, X.; Zhou, J.; Zhou, Y.; et al. Potential role of fecal microbiota in patients with constipation. Therap. Adv. Gastroenterol. 2020, 13, 1756284820968423. [Google Scholar] [CrossRef]
- Cochran, D.; Takis, P.G.; Alexander, J.L.; Mullish, B.H.; Powell, N.; Marchesi, J.R.; Powers, R. Evaluating protocols for reproducible targeted metabolomics by NMR. Analyst 2024, 149, 5423–5432. [Google Scholar] [CrossRef]
- Barnes, S. Overview of Experimental Methods and Study Design in Metabolomics, and Statistical and Pathway Considerations. Methods. Mol. Biol. 2020, 2104, 1–10. [Google Scholar] [CrossRef]
- Yin, X.; Subramanian, S.; Willinger, C.M.; Chen, G.; Juhasz, P.; Courchesne, P.; Chen, B.H.; Li, X.; Hwang, S.J.; Fox, C.S.; et al. Metabolite Signatures of Metabolic Risk Factors and their Longitudinal Changes. J. Clin. Endocrinol. Metab. 2016, 101, 1779–1789. [Google Scholar] [CrossRef]
| Mode of Action | Key Mediators | Molecular Mechanisms | Functional Impacts |
|---|---|---|---|
| Metabolite signaling | SCFAs (butyric acid/propionic acid) | Activate the FFAR2/3 receptor of ENS neurons | Enhances intestinal contraction frequency (+35%) |
| Neurotransmitter precursor supply | Tryptophan → 5-HT | Activation of TPH1 enzyme in enterochromaffin cells | Modulates peristaltic rhythm |
| Immune-mediated regulation | IL-6/IL-1β | Activation of TLR4/NF-K B pathway in glial cells | Induces ENS hypersensitivity |
| Epigenetic modifications | Histone deacetylase inhibition | Inhibition of HDAC2/3 activity in ENS neurons | Promotes neuroplasticity-associated gene expression |
| Target | Change in FC | Levels in FC vs. Healthy | Function | Clinical Correlation | Therapeutic Response | Detection Method | Reference |
|---|---|---|---|---|---|---|---|
| Butyrate | 50–60% decreased | 5.2 vs. 12.3 μM (fecal) | Energy for colonocytes, motility | Correlates with stool frequency (r = 0.58) | Probiotics levels increased by 40% | GC-MS | [51] |
| Propionate | 40% decreased | 8.1 vs. 13.5 μM (fecal) | Anti-inflammatory, barrier | Links to visceral hypersensitivity | Prebiotics restore in 8 weeks | LC-MS | [82] |
| Acetate | 35% decreased | 15.2 vs. 23.4 μM (fecal) | pH regulation, mucus production | Associated with hard stools (p < 0.05) | FMT normalizes in 4 weeks | NMR | [83] |
| Deoxycholic acid (DCA) | 40% dercreased | 1.1 vs. 1.8 mM (fecal) | TGR5 activation → motility | Predicts slow transit (AUC = 0.71) | Rifaximin increased 2-fold | LC-MS | [72] |
| Lithocholic acid (LCA) | 30% decreased | 0.8 vs. 1.2 mM (fecal) | Anti-inflammatory | Correlates with bloating severity | Ursodeoxycholic acid improves | LC-MS/MS | [73] |
| Tryptophan | 30% decreased | 5.9 vs. 8.4 μg/mL (serum) | Serotonin precursor | Links to depression (OR = 2.1) | Probiotics restore to 90% normal | HPLC | [54] |
| GABA | 45% decreased | 2.1 vs. 3.8 μg/mL (serum) | ENS modulation | Correlates with anxiety (r = 0.42) | L. rhamnosus increased 2.5-fold | ELISA | [63] |
| Roseburia spp. | 70% decreased | 3.1% vs. 10.4% (rel. abundance) | Butyrate production | Predicts fiber response (p = 0.02) | XOS abundance increased 2.5-fold | 16S rRNA | [66] |
| Akkermansia muciniphila | 80% decreased | 0.5% vs. 2.5% (rel. abundance) | Mucus layer maintenance | Links to IBS overlap (p = 0.03) | L. plantarum increased 60% | qPCR | [82] |
| Bifidobacterium spp. | 50% decreased | 4.2% vs. 8.4% (rel. abundance) | Acetate production | Correlates with stool consistency | Probiotic cocktails increased 3-fold | Metagenomics | [81] |
| Methane (CH4) | 2-fold increased | ≥3 ppm vs. ≤1 ppm (breath) | Slows motility | OR = 3.2 for bloating | Low-FODMAP diet decreased 50% | Breath test | [79] |
| Hydrogen sulfide (H2S) | 3-fold increased | 2.4 vs. 0.8 mM (fecal) | Inhibits smooth muscle | Links to pain severity (VAS increased 20%) | Bismuth subsalicylate reduces | GC | [78] |
| Butyrate + Tryptophan | Combined decreased | N/A | FC vs. IBS-C differentiation | Sensitivity 85%, Specificity 78% | Probiotics improve panel accuracy | LC-MS/NMR | [81] |
| 2-Oxoglutarate | 60% decreased | 0.4 vs. 1.0 μM (serum) | Arginine metabolism | Predicts fiber response (p = 0.01) | Dietary arginine supplementation | GC-MS | [75] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zheng, F.; Yang, Y.; Lu, G.; Tan, J.S.; Mageswary, U.; Zhan, Y.; Ayad, M.E.; Lee, Y.-Y.; Xie, D. Metabolomics Insights into Gut Microbiota and Functional Constipation. Metabolites 2025, 15, 269. https://doi.org/10.3390/metabo15040269
Zheng F, Yang Y, Lu G, Tan JS, Mageswary U, Zhan Y, Ayad ME, Lee Y-Y, Xie D. Metabolomics Insights into Gut Microbiota and Functional Constipation. Metabolites. 2025; 15(4):269. https://doi.org/10.3390/metabo15040269
Chicago/Turabian StyleZheng, Fan, Yong Yang, Guanting Lu, Joo Shun Tan, Uma Mageswary, Yu Zhan, Mina Ehab Ayad, Yeong-Yeh Lee, and Daoyuan Xie. 2025. "Metabolomics Insights into Gut Microbiota and Functional Constipation" Metabolites 15, no. 4: 269. https://doi.org/10.3390/metabo15040269
APA StyleZheng, F., Yang, Y., Lu, G., Tan, J. S., Mageswary, U., Zhan, Y., Ayad, M. E., Lee, Y.-Y., & Xie, D. (2025). Metabolomics Insights into Gut Microbiota and Functional Constipation. Metabolites, 15(4), 269. https://doi.org/10.3390/metabo15040269