
Changes in Serum Metabolome Following Low-Energy Diet-Induced Weight Loss in Women with Overweight and Prediabetes: A PREVIEW-New Zealand Sub-Study

 ,
,  , ,
, ,  , and
, and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Participants
2.2. Study Treatments
2.3. Clinical Measurements
2.4. Blood Collection
2.5. Metabolomics
2.5.1. Chemicals
2.5.2. Sample Preparation
2.5.3. NMR Spectroscopy
2.5.4. Data Processing and Metabolite Quantification
2.6. Statistical Analysis
3. Results
3.1. Baseline Characteristics of Study Participants
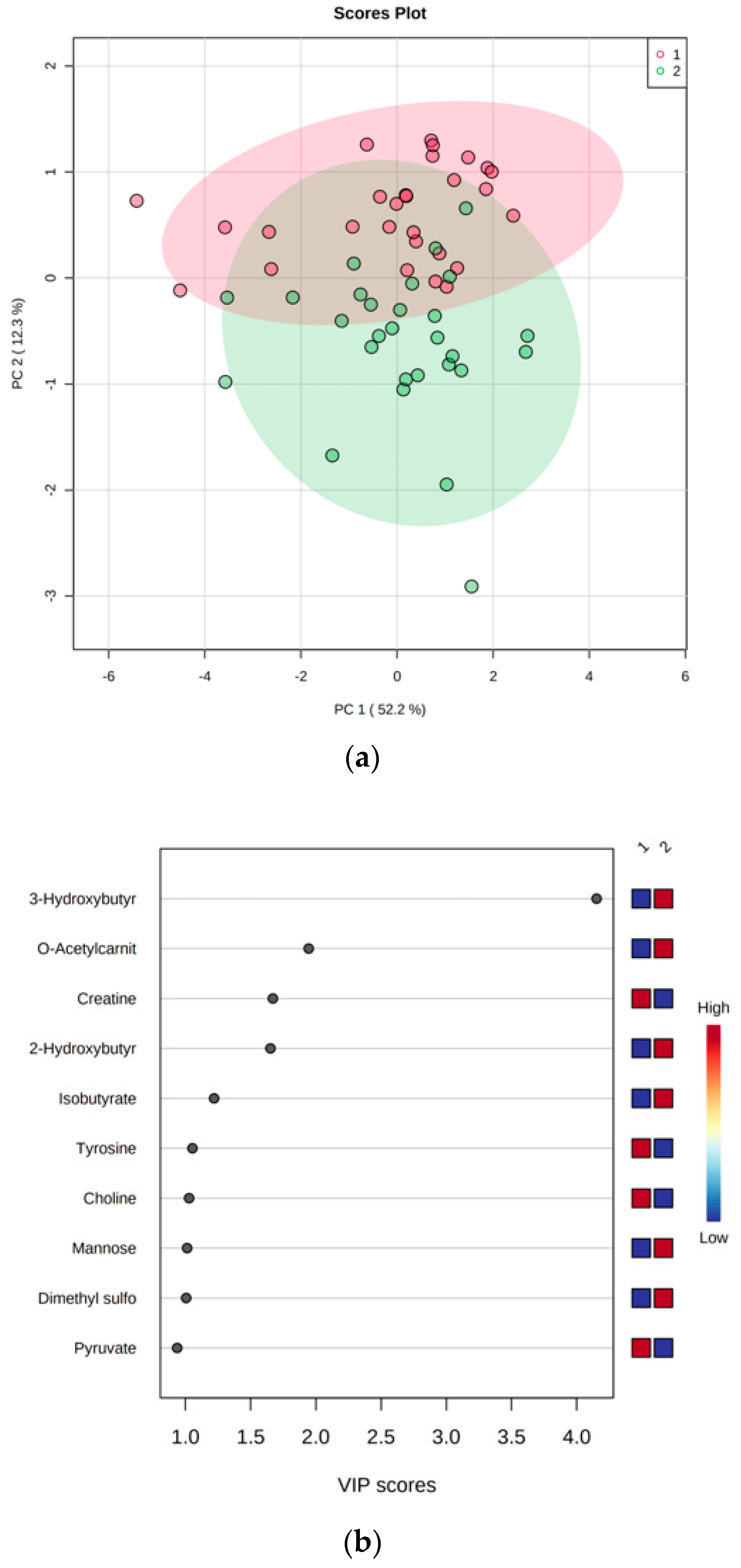
3.2. LED-Dependent Weight Loss Shifts Metabolism Towards Lipid Oxidation
3.3. Association between Changes in Anthropometric and Clinical Parameters and Changes in Metabolite Concentrations
3.4. Baseline Metabolites Do Not Predict Weight Loss
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nyberg, S.T.; Batty, G.D.; Pentti, J.; Virtanen, M.; Alfredsson, L.; Fransson, E.I.; Goldberg, M.; Heikkilä, K.; Jokela, M.; Knutsson, A.; et al. Obesity and Loss of Disease-Free Years Owing to Major Non-Communicable Diseases: A Multicohort Study. Lancet Public Health 2018, 3, e490–e497. [Google Scholar] [CrossRef] [PubMed]
- World Obesity Federation. World Obesity Atlas 2023; World Obesity Federation: London, UK, 2023; Available online: https://data.worldobesity.org/publications/?cat=19 (accessed on 27 February 2024).
- Khan, S.S.; Ning, H.; Wilkins, J.T.; Allen, N.; Carnethon, M.; Berry, J.D.; Sweis, R.N.; Lloyd-Jones, D.M. Association of Body Mass Index With Lifetime Risk of Cardiovascular Disease and Compression of Morbidity. JAMA Cardiol. 2018, 3, 280–287. [Google Scholar] [CrossRef] [PubMed]
- Park, S.; Sadanala, K.C.; Kim, E.K. A Metabolomic Approach to Understanding the Metabolic Link between Obesity and Diabetes. Mol. Cells 2015, 38, 587–596. [Google Scholar] [CrossRef]
- González-Muniesa, P.; Mártinez-González, M.A.; Hu, F.B.; Després, J.P.; Matsuzawa, Y.; Loos, R.J.F.; Moreno, L.A.; Bray, G.A.; Martinez, J.A. Obesity. Nat. Rev. Dis. Primers 2017, 3, 17034. [Google Scholar] [CrossRef] [PubMed]
- Müller, M.J.; Geisler, C.; Heymsfield, S.B.; Bosy-Westphal, A. Recent Advances in Understanding Body Weight Homeostasis in Humans. F1000Research 2018, 7, 1025. [Google Scholar] [CrossRef]
- Yumuk, V.; Tsigos, C.; Fried, M.; Schindler, K.; Busetto, L.; Micic, D.; Toplak, H. European Guidelines for Obesity Management in Adults. Obes. Facts 2015, 8, 402–424. [Google Scholar] [CrossRef] [PubMed]
- Abu Bakar, M.H.; Sarmidi, M.R.; Cheng, K.K.; Ali Khan, A.; Suan, C.L.; Zaman Huri, H.; Yaakob, H. Metabolomics—The Complementary Field in Systems Biology: A Review on Obesity and Type 2 Diabetes. Mol. Biosyst. 2015, 11, 1742–1774. [Google Scholar] [CrossRef]
- Rauschert, S.; Uhl, O.; Koletzko, B.; Hellmuth, C. Metabolomic Biomarkers for Obesity in Humans: A Short Review. Ann. Nutr. Metab. 2014, 64, 314–324. [Google Scholar] [CrossRef]
- Cambiaghi, A.; Ferrario, M.; Masseroli, M. Analysis of Metabolomic Data: Tools, Current Strategies and Future Challenges for Omics Data Integration. Brief. Bioinform. 2017, 18, 498–510. [Google Scholar] [CrossRef]
- Kang, M.; Yoo, H.J.; Kim, M.; Kim, M.; Lee, J.H. Metabolomics Identifies Increases in the Acylcarnitine Profiles in the Plasma of Overweight Subjects in Response to Mild Weight Loss: A Randomized, Controlled Design Study. Lipids Health Dis. 2018, 17, 237. [Google Scholar] [CrossRef]
- Bihlmeyer, N.A.; Kwee, L.C.; Clish, C.B.; Deik, A.A.; Gerszten, R.E.; Pagidipati, N.J.; Laferrère, B.; Svetkey, L.P.; Newgard, C.B.; Kraus, W.E.; et al. Metabolomic Profiling Identifies Complex Lipid Species and Amino Acid Analogues Associated with Response to Weight Loss Interventions. PLoS ONE 2021, 16, e0240764. [Google Scholar] [CrossRef] [PubMed]
- Papandreou, C.; García-Gavilán, J.; Camacho-Barcia, L.; Toft Hansen, T.; Harrold, J.A.; Sjödin, A.; Halford, J.C.G.; Bulló, M. Changes in Circulating Metabolites During Weight Loss Are Associated with Adiposity Improvement, and Body Weight and Adiposity Regain During Weight Loss Maintenance: The SATIN Study. Mol. Nutr. Food Res. 2021, 65, 2001154. [Google Scholar] [CrossRef] [PubMed]
- Geidenstam, N.; Al-Majdoub, M.; Ekman, M.; Spégel, P.; Ridderstråle, M. Metabolite Profiling of Obese Individuals before and after a One Year Weight Loss Program. Int. J. Obes. 2017, 41, 1369–1378. [Google Scholar] [CrossRef]
- Palau-Rodriguez, M.; Garcia-Aloy, M.; Miñarro, A.; Bernal-Lopez, M.R.; Brunius, C.; Gómez-Huelgas, R.; Landberg, R.; Tinahones, F.J.; Andres-Lacueva, C. Effects of a Long-Term Lifestyle Intervention on Metabolically Healthy Women with Obesity: Metabolite Profiles According to Weight Loss Response. Clin. Nutr. 2020, 39, 215–224. [Google Scholar] [CrossRef] [PubMed]
- Stroeve, J.H.M.; Saccenti, E.; Bouwman, J.; Dane, A.; Strassburg, K.; Vervoort, J.; Hankemeier, T.; Astrup, A.; Smilde, A.K.; Van Ommen, B.; et al. Weight Loss Predictability by Plasma Metabolic Signatures in Adults with Obesity and Morbid Obesity of the DiOGenes Study. Obesity 2016, 24, 379–388. [Google Scholar] [CrossRef] [PubMed]
- Rangel-Huerta, O.D.; Pastor-Villaescusa, B.; Gil, A. Are We Close to Defining a Metabolomic Signature of Human Obesity? A Systematic Review of Metabolomics Studies. Metabolomics 2019, 15, 93. [Google Scholar] [CrossRef] [PubMed]
- Perez-Cornago, A.; Brennan, L.; Ibero-Baraibar, I.; Hermsdorff, H.H.M.; O’Gorman, A.; Zulet, M.A.; Martínez, J.A. Metabolomics Identifies Changes in Fatty Acid and Amino Acid Profiles in Serum of Overweight Older Adults Following a Weight Loss Intervention. J. Physiol. Biochem. 2014, 70, 593–602. [Google Scholar] [CrossRef] [PubMed]
- Gu, Y.; Zhao, A.; Huang, F.; Zhang, Y.; Liu, J.; Wang, C.; Jia, W.; Xie, G.; Jia, W. Very Low Carbohydrate Diet Significantly Alters the Serum Metabolic Profiles in Obese Subjects. J. Proteome Res. 2013, 12, 5801–5811. [Google Scholar] [CrossRef] [PubMed]
- International Diabetes Federation. IDF Diabetes Atlas, 10th ed.; International Diabetes Federation: Brussels, Belgium, 2021; ISBN 9782930229980. [Google Scholar]
- Tulipani, S.; Palau-Rodriguez, M.; Miñarro Alonso, A.; Cardona, F.; Marco-Ramell, A.; Zonja, B.; Lopez de Alda, M.; Muñoz-Garach, A.; Sanchez-Pla, A.; Tinahones, F.J.; et al. Biomarkers of Morbid Obesity and Prediabetes by Metabolomic Profiling of Human Discordant Phenotypes. Clin. Chim. Acta 2016, 463, 53–61. [Google Scholar] [CrossRef]
- Vijay, A.; Valdes, A.M. The Metabolomic Signatures of Weight Change. Metabolites 2019, 9, 67. [Google Scholar] [CrossRef]
- Byrne, N.M.; Sainsbury, A.; King, N.A.; Hills, A.P.; Wood, R.E. Intermittent Energy Restriction Improves Weight Loss Efficiency in Obese Men: The MATADOR Study. Int. J. Obes. 2018, 42, 129–138. [Google Scholar] [CrossRef] [PubMed]
- Christensen, P.; Meinert Larsen, T.; Westerterp-Plantenga, M.; Macdonald, I.; Martinez, J.A.; Handjiev, S.; Poppitt, S.; Hansen, S.; Ritz, C.; Astrup, A.; et al. Men and Women Respond Differently to Rapid Weight Loss: Metabolic Outcomes of a Multi-Centre Intervention Study after a Low-Energy Diet in 2500 Overweight, Individuals with Pre-Diabetes (PREVIEW). Diabetes Obes. Metab. 2018, 20, 2840–2851. [Google Scholar] [CrossRef] [PubMed]
- Fogelholm, M.; Larsen, T.M.; Westerterp-Planten, M.; Macdonald, I.; Alfredo Martinez, J.; Boyadjieva, N.; Poppitt, S.; Schlicht, W.; Stratton, G.; Sundvall, J.; et al. PREVIEW: Prevention of Diabetes through Lifestyle Intervention and Population Studies in Europe and around the World. Design, Methods, and Baseline Participant Description of an Adult Cohort Enrolled into a Three-Year Randomised Clinical Trial. Nutrients 2017, 9, 632. [Google Scholar] [CrossRef]
- Tremblay, A.; Fogelholm, M.; Jalo, E.; Westerterp-Plantenga, M.S.; Adam, T.C.; Huttunen-Lenz, M.; Stratton, G.; Lam, T.; Handjieva-Darlenska, T.; Handjiev, S.; et al. What Is the Profile of Overweight Individuals Who Are Unsuccessful Responders to a Low-Energy Diet? A PREVIEW Sub-Study. Front. Nutr. 2021, 8, 707682. [Google Scholar] [CrossRef] [PubMed]
- ADA. Diagnosis and Classification of Diabetes Mellitus. Diabetes Care 2011, 34 (Suppl. S1), S62–S69. [Google Scholar] [CrossRef] [PubMed]
- Raben, A.; Vestentoft, P.S.; Brand-Miller, J.; Jalo, E.; Drummen, M.; Simpson, L.; Martinez, J.A.; Handjieva-Darlenska, T.; Stratton, G.; Huttunen-Lenz, M.; et al. The PREVIEW Intervention Study: Results from a 3-Year Randomized 2 × 2 Factorial Multinational Trial Investigating the Role of Protein, Glycaemic Index and Physical Activity for Prevention of Type 2 Diabetes. Diabetes Obes. Metab. 2021, 23, 324–337. [Google Scholar] [CrossRef] [PubMed]
- Jensen, M.D.; Ryan, D.H.; Apovian, C.M.; Ard, J.D.; Comuzzie, A.G.; Donato, K.A.; Hu, F.B.; Hubbard, V.S.; Jakicic, J.M.; Kushner, R.F.; et al. 2013 AHA/ACC/TOS Guideline for the Management of Overweight and Obesity in Adults. Circulation 2014, 129, 2985–3023. [Google Scholar] [CrossRef] [PubMed]
- Brown, A.; Leeds, A.R. Very Low-Energy and Low-Energy Formula Diets: Effects on Weight Loss, Obesity Co-Morbidities and Type 2 Diabetes Remission—An Update on the Evidence for Their Use in Clinical Practice. Nutr. Bull. 2019, 44, 7–24. [Google Scholar] [CrossRef]
- Tiziani, S.; Emwas, A.H.; Lodi, A.; Ludwig, C.; Bunce, C.M.; Viant, M.R.; Günther, U.L. Optimized Metabolite Extraction from Blood Serum for 1H Nuclear Magnetic Resonance Spectroscopy. Anal. Biochem. 2008, 377, 16–23. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2023. [Google Scholar]
- Martínez-Sánchez, M.A.; Balaguer-Román, A.; Fernández-Ruiz, V.E.; Almansa-Saura, S.; García-Zafra, V.; Ferrer-Gómez, M.; Frutos, M.D.; Queipo-Ortuño, M.I.; Ruiz-Alcaraz, A.J.; Núñez-Sánchez, M.Á.; et al. Plasma Short-Chain Fatty Acid Changes after Bariatric Surgery in Patients with Severe Obesity. Surg. Obes. Relat. Dis. 2023, 19, 727–734. [Google Scholar] [CrossRef]
- Moore, S.C.; Matthews, C.E.; Sampson, J.N.; Stolzenberg-Solomon, R.Z.; Zheng, W.; Cai, Q.; Tan, Y.T.; Chow, W.H.; Ji, B.T.; Liu, D.K.; et al. Human Metabolic Correlates of Body Mass Index. Metabolomics 2014, 10, 259–269. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Xu, Z.; Lu, X.; Yang, X.; Yin, P.; Kong, H.; Yu, Y.; Xu, G. Comprehensive Two-Dimensional Gas Chromatography/Time-of-Flight Mass Spectrometry for Metabonomics: Biomarker Discovery for Diabetes Mellitus. Anal. Chim. Acta 2009, 633, 257–262. [Google Scholar] [CrossRef] [PubMed]
- Gall, W.E.; Beebe, K.; Lawton, K.A.; Adam, K.-P.; Mitchell, M.W.; Nakhle, P.J.; Ryals, J.A.; Milburn, M.V.; Nannipieri, M.; Camastra, S.; et al. Alfa-Hydroxybutyrate Is an Early Biomarker of Insulin Resistance and Glucose Intolerance in a Nondiabetic Population. PLoS ONE 2010, 5, e10883. [Google Scholar] [CrossRef] [PubMed]
- Almanza-Aguilera, E.; Brunius, C.; Bernal-Lopez, M.R.; Garcia-Aloy, M.; Madrid-Gambin, F.; Tinahones, F.J.; Gómez-Huelgas, R.; Landberg, R.; Andres-Lacueva, C. Impact in Plasma Metabolome as Effect of Lifestyle Intervention for Weight-Loss Reveals Metabolic Benefits in Metabolically Healthy Obese Women. J. Proteome Res. 2018, 17, 2600–2610. [Google Scholar] [CrossRef] [PubMed]
- Newgard, C.B.; An, J.; Bain, J.R.; Muehlbauer, M.J.; Stevens, R.D.; Lien, L.F.; Haqq, A.M.; Shah, S.H.; Arlotto, M.; Slentz, C.A.; et al. A Branched-Chain Amino Acid-Related Metabolic Signature That Differentiates Obese and Lean Humans and Contributes to Insulin Resistance. Cell Metab. 2009, 9, 311–326. [Google Scholar] [CrossRef] [PubMed]
- Stephens, F.B.; Constantin-teodosiu, D.; Greenhaff, P.L. New Insights Concerning the Role of Carnitine in the Regulation of Fuel Metabolism in Skeletal Muscle. J. Physiol. 2007, 581, 431–444. [Google Scholar] [CrossRef] [PubMed]
- Mardinoglu, A.; Stančáková, A.; Lotta, L.A.; Kuusisto, J.; Boren, J.; Blüher, M.; Wareham, N.J.; Ferrannini, E.; Groop, P.H.; Laakso, M.; et al. Plasma Mannose Levels Are Associated with Incident Type 2 Diabetes and Cardiovascular Disease. Cell Metab. 2017, 26, 281–283. [Google Scholar] [CrossRef]
- Ferrannini, E.; Bokarewa, M.; Brembeck, P.; Baboota, R.; Hedjazifar, S.; Andersson, K.; Baldi, S.; Campi, B.; Muscelli, E.; Saba, A.; et al. Mannose Is an Insulin-Regulated Metabolite Reflecting Whole-Body Insulin Sensitivity in Man. Metabolism 2020, 102, 153974. [Google Scholar] [CrossRef] [PubMed]
- Food Database. 2-Methylpropanoic Acid. Available online: https://foodb.ca/compounds/FDB003277 (accessed on 21 October 2023).
- Li, N.; Huang, G.; Zhang, Y.; Zheng, N.; Zhao, S.; Wang, J. Diversity of Volatile Compounds in Raw Milk with Different N-6 to n-3 Fatty Acid Ratio. Animals 2022, 12, 252. [Google Scholar] [CrossRef]
- HMDB. Dimethyl Sulfone. Available online: https://hmdb.ca/metabolites/HMDB0004983#identification (accessed on 16 September 2022).
- He, X.; Slupsky, C.M. Metabolic Fingerprint of Dimethyl Sulfone (DMSO2) in Microbial-Mammalian Co-Metabolism. J. Proteome Res. 2014, 13, 5281–5292. [Google Scholar] [CrossRef]
- Zeisel, S.H.; Corbin, K.D. Choline. In Present Knowledge in Nutrition; Erdman, J.W., Macdonald, I.A., Zeisel, S.H., Eds.; Wiley-Blackwell: Hoboken, NJ, USA, 2012; pp. 405–418. [Google Scholar]
- Garcia, E.; Shalaurova, I.; Matyus, S.P.; Wolak-Dinsmore, J.; Oskardmay, D.N.; Connelly, M.A. Quantification of Choline in Serum and Plasma Using a Clinical Nuclear Magnetic Resonance Analyzer. Clin. Chim. Acta 2022, 524, 106–112. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.J.; Larson, M.G.; Vasan, R.S.; Cheng, S.; Rhee, E.P.; McCabe, E.; Lewis, G.D.; Fox, C.S.; Jacques, P.F.; Fernandez, C.; et al. Metabolite Profiles and the Risk of Developing Diabetes. Nat. Med. 2011, 17, 448–453. [Google Scholar] [CrossRef] [PubMed]
- Wyss, M.; Kaddurah-Daouk, R. Creatine and Creatinine Metabolism. Physiol. Rev. 2000, 80, 1107–1213. [Google Scholar] [CrossRef] [PubMed]
- Mittendorfer, B.; Magkos, F.; Fabbrini, E.; Mohammed, B.S.; Klein, S. Relationship between Body Fat Mass and Free Fatty Acid Kinetics in Men and Women. Obesity 2009, 17, 1872–1877. [Google Scholar] [CrossRef]
- Nelson, D.L.; Cox, M.M. Fatty Acid Catabolism. In Lehninger—Principles of Biochemistry; W. H. Freeman and Company: New York, NY, USA, 2008; pp. 652–668. [Google Scholar]

{kind=link}
| All | Weight Loss (<10%) | Weight Loss (≥10%) | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Week 0 | Week 8 | p-Value | Week 0 | Week 8 | p-Value | Week 0 | Week 8 | p-Value | |
| Participants, n (%) | 28 | 28 | 10 (35.7) | 10 (35.7) | 18 (64.3) | 18 (64.3) | |||
| Age (years) | 50.5 ± 6.1 | 50.6 ± 6.8 | 50.5 ± 5.9 | ||||||
| Height (m) | 1.65 ± 0.06 | 1.65 ± 0.06 | 1.65 ± 0.07 | ||||||
| Body weight (kg) | 100.5 ± 16.3 | 89.4 ± 14.9 | <0.001 | 101.6 ± 13.4 | 92.6 ± 12.3 | <0.001 | 99.9 ± 18.1 | 87.5 ± 16.3 | <0.001 |
| BMI (kg/m2) | 37.0 ± 5.5 | 31.7 (30.0–35.2) † | <0.001 | 37.3 ± 4.7 | 34.0 ± 4.3 | <0.001 | 36.8 ± 6.1 | 31.0 (28.3–33.4) † | <0.001 |
| Fat-free mass (kg) | 51.2 ± 6.1 | 48.7 ± 5.9 | <0.001 | 52.1 ± 5.5 | 50.3 ± 5.3 | 0.014 | 50.6 ± 6.5 | 47.8 ± 6.2 | <0.001 |
| Fat mass (kg) | 48.7 ± 11.1 | 40.9 ± 10.4 | <0.001 | 49.3 ± 9.8 | 42.6 ± 8.9 | <0.001 | 46.4 (39.6–51.5) † | 39.9 ± 11.2 | <0.001 |
| Fat mass (%) | 49.6 ± 4.5 | 46.5 ± 4.9 | <0.001 | 49.4 ± 4.5 | 46.8 ± 4.9 | <0.001 | 49.7 ± 4.6 | 46.3 ± 5.1 | <0.001 |
| Waist circumference (cm) | 108.5 ± 11.7 | 97.2 ± 10.6 | <0.001 | 105.6 ± 7.4 | 98.3 ± 7.7 | <0.001 | 110.2 ± 13.5 | 96.5 ± 12.0 | <0.001 |
| Hip circumference (cm) | 122.8 ± 10.1 | 114.2 ± 10.6 | <0.001 | 123.9 ± 10.8 | 115.5 ± 10.2 | <0.001 | 122.3 ± 10.0 | 113.4 ± 11.1 | <0.001 |
| Waist–hip ratio | 0.88 ± 0.06 | 0.8 (0.8–0.9) † | 0.001 | 0.86 ± 0.06 | 0.85 ± 0.07 | 0.887 | 0.90 ± 0.06 | 0.85 ± 0.04 | <0.001 |
| FPG (mmol/L) | 5.8 ± 0.7 | 5.8 ± 1.0 | 0.891 | 6.2 ± 0.6 | 6.2 ± 1.5 | 1.000 | 5.6 ± 0.6 | 5.6 ± 0.7 | 0.820 |
| Fasting insulin (mU/L) | 9.8 (7.9–10.7) † | 5.3 (4.1–7.9) † | <0.001 | 9.1 ± 2.6 | 5.3 (4.0–6.8) † | 0.003 | 10.3 ± 4.2 | 6.8 ± 3.6 | <0.001 |
| HOMA-IR | 2.5 ± 0.8 | 1.3 (1.0–2.2) † | <0.001 | 2.5 ± 0.7 | 1.7 ± 1.0 | 0.006 | 2.5 ± 0.9 | 1.3 (1.0–2.3) † | 0.001 |
| Systolic BP (mmHg) | 122.9 ± 15.7 | 113.7 ± 11.5 | 0.006 * | 122.7 ± 9.7 | 115.5 (111.0–118.6) † | 0.115 | 118.2 (109.5–136.4) † | 113.6 ± 12.1 | 0.020 * |
| Diastolic BP (mmHg) | 65.4 ± 8.4 | 65.1 ± 6.7 | 0.690 * | 68.3 (64.0–72.7) † | 66.6 ± 6.0 | 0.912 | 64.6 ± 7.8 | 64.3 ± 7.1 | 0.727 * |
| Total cholesterol (mmol/L) | 5.4 ± 0.9 | 4.6 ± 0.8 | <0.001 | 5.4 ± 0.7 | 5.0 ± 0.7 | 0.047 | 5.4 ± 1.0 | 4.4 ± 0.8 | <0.001 |
| HDL-C (mmol/L) | 1.4 ± 0.3 | 1.2 ± 0.3 | <0.001 | 1.4 ± 0.3 | 1.4 ± 0.3 | 0.540 | 1.4 ± 0.3 | 1.1 ± 0.2 | <0.001 |
| LDL-C (mmol/L) | 3.4 ± 0.8 | 2.9 ± 0.6 | <0.001 | 3.4 ± 0.6 | 3.1 ± 0.6 | 0.035 | 3.4 ± 0.9 | 2.8 ± 0.7 | <0.001 |
| Triglycerides (mmol/L) | 1.3 ± 0.5 | 1.0 ± 0.3 | 0.002 | 1.3 ± 0.4 | 1.2 ± 0.4 | 0.158 | 1.3 ± 0.5 | 1.0 ± 0.2 | 0.008 |
| All | Weight Loss Groups | Difference between Weight Loss Groups | ||
|---|---|---|---|---|
| <10% | ≥10% | p-Value | ||
| Δ Body weight (kg) | −11.16 ± 2.58 | −8.99 ± 1.18 | −12.37 ± 2.35 | <0.001 |
| Δ BMI (kg/m2) | −4.12 ± 0.97 | −3.30 ± 0.42 | −4.57 ± 0.89 | <0.001 |
| Δ Fat-free mass (kg) | −2.44 ± 1.78 | −1.80 ± 1.87 | −2.80 ± 1.68 | 0.16 |
| Δ Fat mass (kg) | −7.84 ± 2.23 | −6.75 ± 1.34 | −8.44 ± 2.42 | 0.057 |
| Δ Fat mass (%) | −3.06 ± 1.36 | −2.55 ± 1.52 | −3.34 ± 1.22 | 0.382 |
| Δ Waist circumference (cm) | −11.38 ± 5.68 | −7.33 ± 4.18 | −13.64 ± 5.19 | 0.002 |
| Δ Hip circumference (cm) | −8.63 ± 3.55 | −8.33 ± 3.16 | −8.81 ± 3.83 | 0.689 |
| Δ Waist–hip ratio | −0.03 ± 0.04 | 0.00 ± 0.03 | −0.05 ± 0.04 | 0.005 |
| Δ FPG (mmol/L) | −0.03 ± 1.09 | 0.00 ± 1.52 | −0.04 ± 0.82 | 0.944 |
| Δ Fasting insulin (mU/L) | −3.30 ± 2.79 | −3.01 ± 2.33 | −3.46 ± 3.07 | 0.724 |
| Δ HOMA-IR | −0.78 ± 0.75 | −0.79 ± 0.70 | −0.77 ± 0.79 | 0.832 |
| Δ Systolic BP (mmHg) | −3.33 (−18.0–1.33) † | −8.83 ± 16.04 | −4.67 (−16.83–0.08) † | 0.832 |
| Δ Diastolic BP (mmHg) | 0.33 (−1.58–3.58) † | −0.37 ± 10.26 | 0.33 (−1.42–3.33) † | 0.724 |
| Δ Total cholesterol (mmol/L) | −0.80 ± 0.69 | −0.46 ± 0.63 | −0.99 ± 0.65 | 0.051 |
| Δ HDL-C (mmol/L) | −0.17 ± 0.20 | −0.04 ± 0.21 | −0.25 ± 0.16 | 0.021 |
| Δ LDL-C (mmol/L) | −0.51 ± 0.51 | −0.34 ± 0.43 | −0.61 ± 0.53 | 0.245 |
| Δ Triglycerides (mmol/L) | −0.25 ± 0.40 | −0.17 ± 0.35 | −0.30 ± 0.42 | 0.555 |
| Metabolites (mM) | Week 0 | Week 8 | p-Value |
|---|---|---|---|
| 2-Hydroxybutyrate | 0.055 ± 0.018 | 0.070 (0.058–0.092) † | 0.002 * |
| 3-Hydroxybutyrate | 0.045 (0.033–0.070) † | 0.192 (0.106–0.300) † | <0.001 * |
| Acetate | 0.089 ± 0.023 | 0.095 ± 0.021 | 0.181 |
| Acetone | 0.036 (0.023–0.049) † | 0.035 (0.026–0.054) † | 0.966 |
| Alanine | 0.259 ± 0.078 | 0.233 ± 0.067 | 0.082 |
| Betaine | 0.032 ± 0.012 | 0.033 ± 0.012 | 0.617 |
| Carnitine | 0.034 ± 0.010 | 0.030 ± 0.010 | 0.060 |
| Choline | 0.012 ± 0.004 | 0.009 ± 0.002 | 0.008 |
| Creatine | 0.034 ± 0.016 | 0.021 ± 0.011 | <0.001 |
| Creatinine | 0.065 ± 0.016 | 0.070 ± 0.018 | 0.137 |
| Dimethyl sulfone | 0.006 ± 0.003 | 0.008 ± 0.004 | 0.007 |
| Dimethylamine | 0.005 (0.003–0.007) † | 0.005 ± 0.002 | 0.614 |
| Ethanol | 0.180 ± 0.017 | 0.181 ± 0.015 | 0.661 |
| Formate | 0.416 ± 0.085 | 0.417 ± 0.076 | 0.956 |
| Glycerol | 0.112 ± 0.045 | 0.114 ± 0.048 | 0.897 |
| Histidine | 0.046 ± 0.013 | 0.042 (0.040–0.057) † | 0.994 |
| Isobutyrate | 0.008 ± 0.002 | 0.010 ± 0.003 | 0.006 * |
| Isoleucine | 0.048 ± 0.014 | 0.051 ± 0.016 | 0.609 |
| Lactate | 2.297 ± 0.965 | 1.982 ± 0.659 | 0.066 |
| Leucine | 0.102 ± 0.028 | 0.102 ± 0.027 | 0.990 |
| Lysine | 0.055 ± 0.017 | 0.052 ± 0.013 | 0.357 |
| Mannose | 0.047 ± 0.013 | 0.056 ± 0.015 | 0.003 |
| Methanol | 0.275 ± 0.020 | 0.268 (0.259–0.276) † | 0.690 |
| Methionine | 0.021 ± 0.006 | 0.021 ± 0.006 | 0.695 |
| Methylamine | 0.018 ± 0.005 | 0.021 ± 0.006 | 0.111 |
| O-Acetylcarnitine | 0.007 ± 0.002 | 0.010 (0.009–0.014) † | <0.001 * |
| O-Phosphocholine | 0.003 (0.001–0.004) † | 0.003 ± 0.002 | 0.289 |
| Phenylalanine | 0.045 ± 0.014 | 0.041 ± 0.010 | 0.174 |
| Pyruvate | 0.047 ± 0.019 | 0.039 ± 0.018 | 0.053 |
| Threonine | 0.111 ± 0.043 | 0.106 ± 0.030 | 0.646 |
| Trimethylamine N-oxide | 0.005 (0.003–0.007) † | 0.005 ± 0.002 | 0.531 * |
| Tryptophan | 0.046 ± 0.012 | 0.045 ± 0.011 | 0.868 |
| Tyrosine | 0.058 ± 0.016 | 0.048 ± 0.013 | 0.005 |
| Valine | 0.184 ± 0.053 | 0.180 ± 0.040 | 0.716 |
| Δ 2-Hydroxybutyrate | p-Value | Δ 3-Hydroxybutyrate | p-Value | Δ Choline | p-Value | Δ Creatine | p-Value | Δ Dimethyl Sulfone | p-Value | |
| Δ BMI (kg/m2) | −0.05 (−16.62; 16.52) | 0.995 | −35.99 (−101.53; 29.54) | 0.282 | 0.26 (−2.16; 2.68) | 0.833 | 0.63 (−6.02; 7.28) | 0.853 | 0.79 (−0.62; 2.20) | 0.271 |
| Δ FM (%) | −6.10 (−11.62; −0.59) | 0.030 | −36.66 (−70.28; −3.05) | 0.033 | −1.60 (−3.79; 0.60) | 0.154 | −1.97 (−6.61; 2.68) | 0.406 | −0.52 (−1.62; 0.58) | 0.355 |
| Δ Fasting insulin (mU/L) | −3.00 (−6.08; 0.07) | 0.055 | −23.19 (−35.53; −10.85) | < 0.001 | −0.35 (−1.18; 0.47) | 0.399 | 0.08 (−1.91; 2.06) | 0.941 | −0.13 (−0.56; 0.30) | 0.550 |
| Δ HDL-C (mmol/L) | −1.97 (−81.26; 77.32) | 0.961 | −90.85 (−500.61; 318.90) | 0.664 | 1.67 (−16.10; 19.45) | 0.854 | 15.38 (−23.23; 53.99) | 0.435 | −1.35 (−12.64; 9.94) | 0.815 |
| Δ TG (mmol/L) | −34.27 (−65.98; −2.56) | 0.034 | −160.16 (−366.38; 46.06) | 0.128 | −4.80 (−8.61; −0.99) | 0.014 | −7.06 (−24.63; 10.51) | 0.431 | −2.13 (−5.35; 1.08) | 0.193 |
| Δ Waist circumference (cm) | −0.39 (−2.73; 1.95) | 0.746 | −4.25 (−21.26; 12.77) | 0.625 | 0.09 (−0.37; 0.56) | 0.692 | 0.54 (−0.93; 2.01) | 0.472 | 0.15 (−0.09; 0.39) | 0.223 |
| Δ Isobutyrate | p-Value | Δ Mannose | p-Value | Δ O-Acetylcarnitine | p-Value | Δ Tyrosine | p-Value | |||
| Δ BMI (kg/m2) | 0.06 (−1.12; 1.23) | 0.924 | −5.41 (−13.19; 2.38) | 0.173 | −0.46 (−2.44; 1.53) | 0.652 | 3.23 (−2.71; 9.16) | 0.287 | ||
| Δ FM (%) | −0.69 (−1.66; 0.27) | 0.158 | −4.13 (−8.06; −0.19) | 0.040 | −1.23 (−2.16; −0.30) | 0.010 | −3.40 (−7.69; 0.89) | 0.120 | ||
| Δ Fasting insulin (mU/L) | 0.18 (−0.17; 0.53) | 0.316 | 0.60 (−1.42; 2.63) | 0.558 | −0.23 (−0.73; 0.26) | 0.358 | 0.74 (−1.42; 2.89) | 0.504 | ||
| Δ HDL-C (mmol/L) | −0.95 (−8.33; 6.43) | 0.801 | −2.76 (−25.97; 20.44) | 0.815 | 2.45 (−10.05; 14.95) | 0.700 | 14.06 (−30.29; 58.42) | 0.534 | ||
| Δ TG (mmol/L) | −1.31 (−5.31; 2.69) | 0.521 | −13.46 (−34.78; 7.86) | 0.216 | −7.19 (−13.08; −1.30) | 0.017 | −14.96 (−58.72; 28.81) | 0.503 | ||
| Δ Waist circumference (cm) | −0.04 (−0.27; 0.19) | 0.735 | 0.10 (−0.84; 1.04) | 0.832 | 0.03 (−0.39; 0.44) | 0.899 | 0.38 (−1.83; 2.59) | 0.736 | ||
| Model 1 | Model 2 | |||
|---|---|---|---|---|
| β (95% CI) | p-Value | β (95% CI) | p-Value | |
| 2-Hydroxybutyrate | 1.5 (−41.19; 44.09) | 0.947 | 2.3 (−47.18; 51.80) | 0.927 |
| 3-Hydroxybutyrate | −8.6 (−29.24; 12.04) | 0.414 | −10.8 (−43.33; 21.66) | 0.513 |
| Acetate | 25.5 (−12.75; 63.76) | 0.191 | 27.0 (−15.63; 69.65) | 0.214 |
| Acetone | 16.3 (−21.68; 54.43) | 0.399 | 23.5 (−25.25; 72.21) | 0.345 |
| Alanine | 3.4 (−6.93; 13.67) | 0.521 | 5.3 (−6.30; 16.91) | 0.370 |
| Betaine | 24.8 (−44.22; 93.72) | 0.482 | 40.5 (−54.28; 135.24) | 0.402 |
| Carnitine | −2.3 (−78.06; 73.42) | 0.952 | 27.1 (−64.75; 118.86) | 0.564 |
| Choline | −16.8 (−207.99; 174.33) | 0.863 | 15.4 (−207.20; 238.04) | 0.892 |
| Creatine | 10.3 (−40.45; 60.97) | 0.692 | 0.9 (−62.08; 63.93) | 0.977 |
| Creatinine | 14.8 (−34.75; 64.41) | 0.558 | 30.2 (−28.80; 89.25) | 0.316 |
| Dimethyl sulfone | 62.9 (−205.97; 331.80) | 0.647 | 91.1 (−260.56; 442.73) | 0.612 |
| Dimethylamine | 157.6 (−157.10; 472.30) | 0.326 | 244.4 (−175.07; 663.80) | 0.254 |
| Ethanol | 7.1 (−40.01; 54.10) | 0.769 | −17.9 (−74.55; 38.72) | 0.535 |
| Formate | 3.5 (−6.02; 13.07) | 0.470 | 5.5 (−5.53; 16.49) | 0.329 |
| Glycerol | −15.7 (−35.13; 3.80) | 0.115 | −14.3 (−37.63; 8.97) | 0.228 |
| Histidine | 30.3 (−31.61; 92.11) | 0.338 | 32.1 (−38.38; 102.48) | 0.372 |
| Isobutyrate | 61.0 (−255.69; 377.70) | 0.706 | 58.0 (−307.51; 423.47) | 0.756 |
| Isoleucine | 29.7 (−29.49; 88.94) | 0.325 | 33.8 (−30.92; 98.53) | 0.306 |
| Lactate | 0.4 (−0.53; 1.23) | 0.432 | 0.4 (−0.54; 1.36) | 0.393 |
| Leucine | 14.7 (−13.51; 42.82) | 0.308 | 14.9 (−16.44; 46.15) | 0.352 |
| Lysine | 15.1 (−32.74; 62.89) | 0.537 | 10.9 (−40.25; 61.98) | 0.677 |
| Mannose | −17.8 (−81.19; 45.51) | 0.581 | −24.0 (−92.75; 44.78) | 0.494 |
| Methanol | 12.4 (−29.84; 54.66) | 0.565 | 17.9 (−33.45; 69.31) | 0.494 |
| Methionine | 72.4 (−65.33; 210.03) | 0.303 | 100.0 (−68.22; 267.26) | 0.245 |
| Methylamine | 100.2 (−65.18; 265.50) | 0.235 | 65.7 (−111.27; 242.66) | 0.467 |
| O-Acetylcarnitine | −82.1 (−402.74; 238.49) | 0.616 | −68.4 (−460.72; 323.84) | 0.732 |
| O-Phosphocholine | −44.4 (−372.31; 283.45) | 0.791 | −123.1 (−502.88; 256.63) | 0.525 |
| Phenylalanine | 16.8 (−41.12; 74.74) | 0.570 | 21.0 (−44.84; 86.74) | 0.533 |
| Pyruvate | 23.7 (−22.06; 69.46) | 0.310 | 14.9 (−32.15; 62.00) | 0.534 |
| Threonine | 6.5 (−12.40; 25.30) | 0.502 | 8.6 (−17.45; 34.61) | 0.518 |
| Trimethylamine N-oxide | 30.0 (−112.81; 172.77) | 0.681 | 0.7 (−196.31; 197.61) | 0.995 |
| Tryptophan | 9.0 (−54.52; 72.51) | 0.781 | 19.1 (−52.69; 90.79) | 0.603 |
| Tyrosine | 27.8 (−22.42; 77.92) | 0.278 | 26.6 (−28.70; 81.98) | 0.345 |
| Valine | 6.1 (−8.79; 21.02) | 0.422 | 6.8 (−9.18; 22.81) | 0.403 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Relva, B.; Samuelsson, L.M.; Duarte, I.F.; Fasol, U.; Edwards, P.J.B.; Fogelholm, M.; Raben, A.; Poppitt, S.D.; Silvestre, M.P. Changes in Serum Metabolome Following Low-Energy Diet-Induced Weight Loss in Women with Overweight and Prediabetes: A PREVIEW-New Zealand Sub-Study. Metabolites 2024, 14, 401. https://doi.org/10.3390/metabo14080401
Relva B, Samuelsson LM, Duarte IF, Fasol U, Edwards PJB, Fogelholm M, Raben A, Poppitt SD, Silvestre MP. Changes in Serum Metabolome Following Low-Energy Diet-Induced Weight Loss in Women with Overweight and Prediabetes: A PREVIEW-New Zealand Sub-Study. Metabolites. 2024; 14(8):401. https://doi.org/10.3390/metabo14080401
Chicago/Turabian StyleRelva, Bárbara, Linda M. Samuelsson, Iola F. Duarte, Ulrike Fasol, Patrick J. B. Edwards, Mikael Fogelholm, Anne Raben, Sally D. Poppitt, and Marta P. Silvestre. 2024. "Changes in Serum Metabolome Following Low-Energy Diet-Induced Weight Loss in Women with Overweight and Prediabetes: A PREVIEW-New Zealand Sub-Study" Metabolites 14, no. 8: 401. https://doi.org/10.3390/metabo14080401
APA StyleRelva, B., Samuelsson, L. M., Duarte, I. F., Fasol, U., Edwards, P. J. B., Fogelholm, M., Raben, A., Poppitt, S. D., & Silvestre, M. P. (2024). Changes in Serum Metabolome Following Low-Energy Diet-Induced Weight Loss in Women with Overweight and Prediabetes: A PREVIEW-New Zealand Sub-Study. Metabolites, 14(8), 401. https://doi.org/10.3390/metabo14080401