Metabolites Associated with Polygenic Risk of Breast Cancer

 ,
,  and
and
Abstract
1. Introduction
2. Materials and Methods
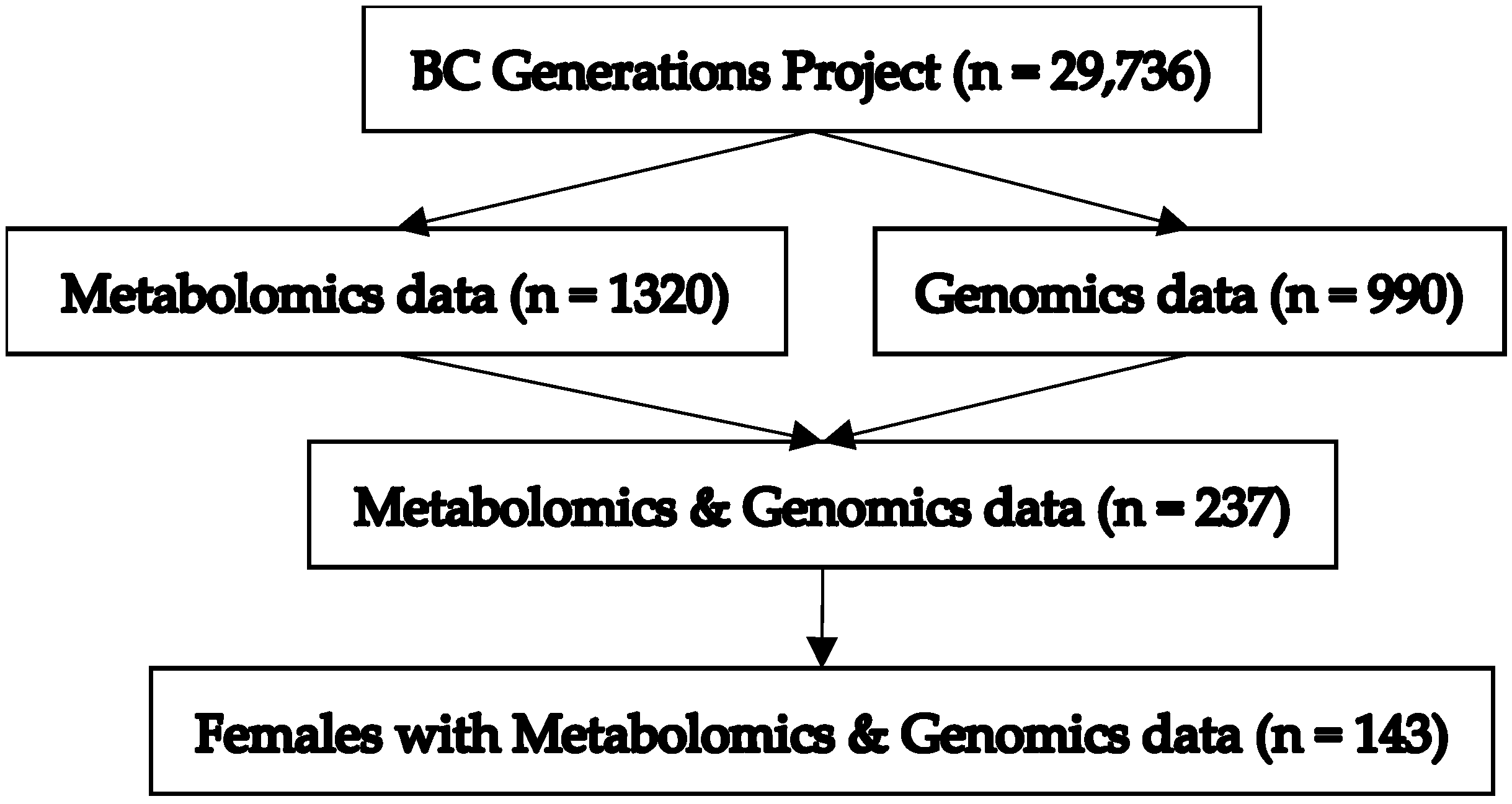
2.1. Study Population
2.2. Metabolomics Data
2.3. Genomics Data
2.4. Polygenic Risk Score
2.5. Statistical Analysis
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fachal, L.; Dunning, A.M. From candidate gene studies to GWAS and post-GWAS analyses in breast cancer. Curr. Opin. Genet. Dev. 2015, 30, 32–41. [Google Scholar] [CrossRef] [PubMed]
- Fanale, D.; Amodeo, V.; Corsini, L.R.; Rizzo, S.; Bazan, V.; Russo, A. Breast cancer genome-wide association studies: There is strength in numbers. Oncogene 2012, 31, 2121–2128. [Google Scholar] [CrossRef] [PubMed]
- Nik-Zainal, S.; Davies, H.; Staaf, J.; Ramakrishna, M.; Glodzik, D.; Zou, X.; Martincorena, I.; Alexandrov, L.B.; Martin, S.; Wedge, D.C.; et al. Landscape of somatic mutations in 560 breast cancer whole-genome sequences. Nature 2016, 534, 47–54. [Google Scholar] [CrossRef] [PubMed]
- Pharoah, P.D.P.; Antoniou, A.; Bobrow, M.; Zimmern, R.L.; Easton, D.F.; Ponder, B.A.J. Polygenic susceptibility to breast cancer and implications for prevention. Nat. Genet. 2002, 31, 33–36. [Google Scholar] [CrossRef]
- Ghoussaini, M.; Pharoah, P.D.P. Polygenic susceptibility to breast cancer: Current state-of-the-art. Future Oncol. 2009, 5, 689–701. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Rowley, S.M.; Thompson, E.R.; McInerny, S.; Devereux, L.; Amarasinghe, K.C.; Zethoven, M.; Lupat, R.; Goode, D.; Li, J.; et al. Evaluating the breast cancer predisposition role of rare variants in genes associated with low-penetrance breast cancer risk SNPs. Breast Cancer Res. 2018, 20, 3. [Google Scholar] [CrossRef] [PubMed]
- Hsu, P.P.; Sabatini, D.M. Cancer cell metabolism: Warburg and beyond. Cell 2008, 134, 703–707. [Google Scholar] [CrossRef] [PubMed]
- Gieger, C.; Geistlinger, L.; Altmaier, E.; de Angelis, M.H.; Kronenberg, F.; Meitinger, T.; Mewes, H.-W.; Wichmann, H.-E.; Weinberger, K.M.; Adamski, J.; et al. Genetics meets metabolomics: A genome-wide association study of metabolite profiles in human serum. PLoS Genet. 2008, 4, e1000282. [Google Scholar] [CrossRef] [PubMed]
- Lécuyer, L.; Bala, A.V.; Deschasaux, M.; Bouchemal, N.; Triba, M.N.; Vasson, M.-P.; Rossary, A.; Demidem, A.; Galan, P.; Hercberg, S.; et al. NMR metabolomic signatures reveal predictive plasma metabolites associated with long-term risk of developing breast cancer. Int. J. Epidemiol. 2018, 47, 484–494. [Google Scholar] [CrossRef]
- Lofterød, T.; Mortensen, E.S.; Nalwoga, H.; Wilsgaard, T.; Frydenberg, H.; Risberg, T.; Eggen, A.E.; McTiernan, A.; Aziz, S.; Wist, E.A.; et al. Impact of pre-diagnostic triglycerides and HDL-cholesterol on breast cancer recurrence and survival by breast cancer subtypes. BMC Cancer 2018, 18, 654. [Google Scholar] [CrossRef]
- His, M.; Viallon, V.; Dossus, L.; Gicquiau, A.; Achaintre, D.; Scalbert, A.; Ferrari, P.; Romieu, I.; Onland-Moret, N.C.; Weiderpass, E.; et al. Prospective analysis of circulating metabolites and breast cancer in EPIC. BMC Med. 2019, 17, 178. [Google Scholar] [CrossRef] [PubMed]
- Moore, S.C.; Mazzilli, K.M.; Sampson, J.N.; Matthews, C.E.; Carter, B.D.; Playdon, M.C.; Wang, Y.; Stevens, V.L. A Metabolomics Analysis of Postmenopausal Breast Cancer Risk in the Cancer Prevention Study II. Metabolites 2021, 11, 95. [Google Scholar] [CrossRef]
- Zeleznik, O.A.; Balasubramanian, R.; Zhao, Y.; Frueh, L.; Jeanfavre, S.; Avila-Pacheco, J.; Clish, C.B.; Tworoger, S.S.; Eliassen, A.H. Circulating amino acids and amino acid-related metabolites and risk of breast cancer among predominantly premenopausal women. NPJ Breast Cancer 2021, 7, 54. [Google Scholar] [CrossRef] [PubMed]
- Dhalla, A.; McDonald, T.E.; Gallagher, R.P.; Spinelli, J.J.; Brooks-Wilson, A.R.; Lee, T.K.; Lai, C.; Borugian, M.J.; Woods, R.R.; Le, N.D.; et al. Cohort Profile: The British Columbia Generations Project (BCGP). Int. J. Epidemiol. 2018, 48, 377–378. [Google Scholar] [CrossRef]
- Gu, Q.; Spinelli, J.J.; Dummer, T.B.J.; McDonald, T.E.; Moore, S.C.; Murphy, R.A. Metabolic profiling of adherence to diet, physical activity and body size recommendations for cancer prevention. Sci. Rep. 2018, 8, 16293. [Google Scholar] [CrossRef] [PubMed]
- Qi, J.; Spinelli, J.J.; Dummer, T.J.B.; Bhatti, P.; Playdon, M.C.; Levitt, J.O.; Hauner, B.; Moore, S.C.; Murphy, R.A. Metabolomics and cancer preventive behaviors in the BC Generations Project. Sci. Rep. 2021, 11, 12094. [Google Scholar] [CrossRef]
- Soininen, P.; Kangas, A.J.; Würtz, P.; Tukiainen, T.; Tynkkynen, T.; Laatikainen, R.; Jarvelin, M.-R.; Kähönen, M.; Lehtimäki, T.; Viikari, J.; et al. High-throughput serum NMR metabonomics for cost-effective holistic studies on systemic metabolism. Analyst 2009, 134, 1781–1785. [Google Scholar] [CrossRef] [PubMed]
- Das, S.; Forer, L.; Schönherr, S.; Sidore, C.; Locke, A.E.; Kwong, A.; Vrieze, S.I.; Chew, E.Y.; Levy, S.; McGue, M.; et al. Next-generation genotype imputation service and methods. Nat. Genet. 2016, 48, 1284–1287. [Google Scholar] [CrossRef] [PubMed]
- Mavaddat, N.; Michailidou, K.; Dennis, J.; Lush, M.; Fachal, L.; Lee, A.; Tyrer, J.P.; Chen, T.-H.; Wang, Q.; Bolla, M.K.; et al. Polygenic Risk Scores for Prediction of Breast Cancer and Breast Cancer Subtypes. Am. J. Hum. Genet. 2019, 104, 21–34. [Google Scholar] [CrossRef] [PubMed]
- Khera, A.V.; Chaffin, M.; Aragam, K.G.; Haas, M.E.; Roselli, C.; Choi, S.H.; Natarajan, P.; Lander, E.S.; Lubitz, S.A.; Ellinor, P.T.; et al. Genome-wide polygenic scores for common diseases identify individuals with risk equivalent to monogenic mutations. Nat. Genet. 2018, 50, 1219–1224. [Google Scholar] [CrossRef]
- Benjamini, Y.; Hochberg, Y. Controlling the False Discovery Rate: A Practical and Powerful Approach to Multiple Testing. J. R. Stat. Soc. Ser. B 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Nouri, M.; Mohsenpour, M.A.; Katsiki, N.; Ghobadi, S.; Jafari, A.; Faghih, S.; Banach, M.; Mazidi, M. Effect of Serum Lipid Profile on the Risk of Breast Cancer: Systematic Review and Meta-Analysis of 1,628,871 Women. J. Clin. Med. 2022, 11, 4503. [Google Scholar] [CrossRef] [PubMed]
- Cedó, L.; Reddy, S.T.; Mato, E.; Blanco-Vaca, F.; Escolà-Gil, J.C. HDL and LDL: Potential New Players in Breast Cancer Development. J. Clin. Med. 2019, 8, 853. [Google Scholar] [CrossRef]
- Shah, F.D.; Shukla, S.N.; Shah, P.M.; Patel, H.R.H.; Patel, P.S. Significance of alterations in plasma lipid profile levels in breast cancer. Integr. Cancer Ther. 2008, 7, 33–41. [Google Scholar] [CrossRef]
- Kökoğlu, E.; Karaarslan, I.; Karaarslan, H.M.; Baloğlu, H. Alterations of serum lipids and lipoproteins in breast cancer. Cancer Lett. 1994, 82, 175–178. [Google Scholar] [CrossRef] [PubMed]
- Chang, S.-J.; Hou, M.-F.; Tsai, S.-M.; Wu, S.-H.; Hou, L.A.; Ma, H.; Shann, T.-Y.; Tsai, L.-Y. The association between lipid profiles and breast cancer among Taiwanese women. Clin. Chem. Lab. Med. 2007, 45, 1219–1223. [Google Scholar] [CrossRef] [PubMed]
- Kumar, K.; Sachdanandam, P.; Arivazhagan, R. Studies on the changes in plasma lipids and lipoproteins in patients with benign and malignant breast cancer. Biochem. Int. 1991, 23, 581–589. [Google Scholar]
- Feingold, K.R. Introduction to Lipids and Lipoproteins; Feingold, K.R., Anawalt, B., Blackman, M.R., Boyce, A., Chrousos, G., Corpas, E., de Herder, W.W., Dhatariya, K., Dungan, K., Hofland, J., et al., Eds.; Endotext; South Dartmouth; MDText.com, Inc.: South Dartmout MA, USA, 2000. Available online: http://www.ncbi.nlm.nih.gov/books/NBK305896/ (accessed on 9 September 2022).
- Agnoli, C.; Grioni, S.; Sieri, S.; Sacerdote, C.; Ricceri, F.; Tumino, R.; Frasca, G.; Pala, V.; Mattiello, A.; Chiodini, P.; et al. Metabolic syndrome and breast cancer risk: A case-cohort study nested in a multicentre italian cohort. PLoS ONE 2015, 10, e0128891. [Google Scholar] [CrossRef]
- Gordillo-Moscoso, A.; Ruiz, E.; Carnero, M.; Reguillo, F.; Rodriguez, E.; Tejerina, T.; Redondo, S. Relationship between serum levels of triglycerides and vascular inflammation, measured as COX-2, in arteries from diabetic patients: A translational study. Lipids Health Dis. 2013, 12, 62. [Google Scholar] [CrossRef]
- Harris, R.E.; Casto, B.C.; Harris, Z.M. Cyclooxygenase-2 and the inflammogenesis of breast cancer. World J. Clin. Oncol. 2014, 5, 677–692. [Google Scholar] [CrossRef]
- Sampson, J.N.; Boca, S.M.; Shu, X.O.; Stolzenberg-Solomon, R.Z.; Matthews, C.E.; Hsing, A.W.; Tan, Y.T.; Ji, B.-T.; Chow, W.-H.; Cai, Q.; et al. Metabolomics in epidemiology: Sources of variability in metabolite measurements and implications. Cancer Epidemiol. Biomark. Prev. 2013, 22, 631–640. [Google Scholar] [CrossRef] [PubMed]
- Sud, A.; Kinnersley, B.; Houlston, R.S. Genome-wide association studies of cancer: Current insights and future perspectives. Nat. Rev. Cancer 2017, 17, 692–704. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
| Characteristic | N (%) | Mean (SD) | Min, Max |
|---|---|---|---|
| Age (years) | 55.7 (7.7) | 40, 69 | |
| BMI (kg/m2) | 26.0 (4.8) | 18.6, 42.4 | |
| 18.5–25 | 75 (52.4%) | ||
| >25 | 68 (47.6%) | ||
| Alcohol Consumption | |||
| Never | 11 (7.7%) | ||
| <1/month | 23 (16.1%) | ||
| 1/month | 23 (16.1%) | ||
| 2–3/month | 15 (10.5%) | ||
| 1/week | 20 (14.0%) | ||
| 2–3/week | 25 (17.5%) | ||
| 4–5/week | 14 (9.8%) | ||
| 6–7/week | 12 (8.4%) | ||
| Age of menarche (years) | 13 (1.36) | 9, 17 | |
| Gravidity | 2.5 (1.57) | 0, 8 | |
| Menopausal status at baseline | |||
| Pre-menopausal | 59 (41.3%) | ||
| Post-menopausal | 84 (58.7%) | ||
| Family History of Any Cancer | |||
| Yes | 98 (69%) | ||
| No | 45 (31%) | ||
| Family History of Breast Cancer | |||
| Mother | 17 (11.9%) | ||
| Sibling | 6 (4.2%) |
| Metabolite Measure Notation a | Metabolite Measure Description a | β-Estimate b | p-Value | q-Value c |
|---|---|---|---|---|
| S-VLDL-TG | Small VLDL triglycerides | 0.45 | 0.005 | 0.1 |
| XL-HDL-TG-% | Very large HDL triglycerides to total lipids ratio, % | 0.35 | 0.005 | 0.1 |
| L-VLDL-TG | Large VLDL triglycerides | 0.51 | 0.006 | 0.1 |
| L-VLDL-P | Large VLDL particles | 0.50 | 0.006 | 0.1 |
| L-VLDL-L | Large VLDL lipids | 0.50 | 0.006 | 0.1 |
| L-VLDL-PL | Large VLDL phospholipids | 0.50 | 0.006 | 0.1 |
| L-VLDL-CE | Large VLDL cholesterol esters | 0.50 | 0.006 | 0.1 |
| M-VLDL-TG | Medium VLDL triglycerides | 0.45 | 0.006 | 0.1 |
| L-VLDL-C | Large VLDL cholesterol | 0.50 | 0.008 | 0.1 |
| M-VLDL-P | Medium VLDL particles | 0.42 | 0.008 | 0.1 |
| M-VLDL-PL | Medium VLDL phospholipids | 0.41 | 0.008 | 0.1 |
| M-VLDL-L | Medium VLDL lipids | 0.41 | 0.009 | 0.1 |
| S-VLDL-P | Small VLDL particles | 0.40 | 0.009 | 0.1 |
| L-HDL-TG-% | Large HDL triglycerides to total lipids ratio, % | 0.34 | 0.009 | 0.1 |
| M-VLDL-TG-% | Medium VLDL triglycerides to total lipids ratio, % | 0.53 | 0.01 | 0.1 |
| VLDL-TG | VLDL triglycerides | 0.42 | 0.01 | 0.1 |
| S-HDL-TG | Small HDL triglycerides | 0.42 | 0.01 | 0.1 |
| VLDL-D | Mean diameter of VLDL particles | 0.40 | 0.01 | 0.1 |
| S-VLDL-L | Small VLDL lipids | 0.39 | 0.01 | 0.1 |
| M-HDL-TG-% | Medium HDL triglycerides to total lipids ratio, % | 0.39 | 0.01 | 0.1 |
| S-VLDL-PL | Small VLDL phospholipids | 0.39 | 0.01 | 0.1 |
| M-VLDL-FC | Medium VLDL free cholesterol | 0.38 | 0.01 | 0.1 |
| ApoB/ApoA1 | Apolipoprotein B to apolipoprotein A1 ratio | 0.37 | 0.01 | 0.1 |
| TG/PG | Ratio of triglycerides to phosphoglycerides | 0.36 | 0.01 | 0.1 |
| HDL2-C | HDL2 cholesterol | −0.33 | 0.01 | 0.1 |
| HDL-C | HDL cholesterol | −0.35 | 0.01 | 0.1 |
| HDL-D | Mean diameter of HDL particles | −0.37 | 0.01 | 0.1 |
| M-VLDL-CE-% | Medium VLDL cholesterol esters to total lipids ratio, % | −0.41 | 0.01 | 0.1 |
| XL-VLDL-L | Very large VLDL lipids | 0.41 | 0.02 | 0.1 |
| L-VLDL-FC | Large VLDL free cholesterol | 0.42 | 0.02 | 0.1 |
| XL-VLDL-TG | Very large VLDL triglycerides | 0.42 | 0.02 | 0.1 |
| S-VLDL-TG-% | Small VLDL triglycerides to total lipids ratio, % | 0.42 | 0.02 | 0.1 |
| XL-VLDL-P | Very large VLDL particles | 0.41 | 0.02 | 0.1 |
| S-HDL-TG-% | Small HDL triglycerides to total lipids ratio, % | 0.40 | 0.02 | 0.1 |
| XL-VLDL-C | Very large VLDL cholesterol | 0.39 | 0.02 | 0.1 |
| XS-VLDL-TG-% | Very small VLDL triglycerides to total lipids ratio, % | 0.38 | 0.02 | 0.1 |
| Serum-TG | Serum triglycerides | 0.37 | 0.02 | 0.1 |
| XS-VLDL-TG | Very small VLDL triglycerides | 0.36 | 0.02 | 0.1 |
| S-VLDL-FC | Small VLDL free cholesterol | 0.36 | 0.02 | 0.1 |
| L-HDL-C | Large HDL total cholesterol | −0.31 | 0.02 | 0.1 |
| L-HDL-CE | Large HDL cholesterol esters | −0.32 | 0.02 | 0.1 |
| L-HDL-P | Large HDL particles | −0.32 | 0.02 | 0.1 |
| L-HDL-L | Large HDL lipids | −0.32 | 0.02 | 0.1 |
| L-HDL-PL | Large HDL phospholipids | −0.33 | 0.02 | 0.1 |
| M-VLDL-C-% | Medium VLDL total cholesterol to total lipids ratio, % | −0.39 | 0.02 | 0.1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Samuels, E.; Parks, J.; Chu, J.; McDonald, T.; Spinelli, J.; Murphy, R.A.; Bhatti, P. Metabolites Associated with Polygenic Risk of Breast Cancer. Metabolites 2024, 14, 295. https://doi.org/10.3390/metabo14060295
Samuels E, Parks J, Chu J, McDonald T, Spinelli J, Murphy RA, Bhatti P. Metabolites Associated with Polygenic Risk of Breast Cancer. Metabolites. 2024; 14(6):295. https://doi.org/10.3390/metabo14060295
Chicago/Turabian StyleSamuels, Elizabeth, Jaclyn Parks, Jessica Chu, Treena McDonald, John Spinelli, Rachel A. Murphy, and Parveen Bhatti. 2024. "Metabolites Associated with Polygenic Risk of Breast Cancer" Metabolites 14, no. 6: 295. https://doi.org/10.3390/metabo14060295
APA StyleSamuels, E., Parks, J., Chu, J., McDonald, T., Spinelli, J., Murphy, R. A., & Bhatti, P. (2024). Metabolites Associated with Polygenic Risk of Breast Cancer. Metabolites, 14(6), 295. https://doi.org/10.3390/metabo14060295