1. Introduction
The development of antibiotics has revolutionized public health and prevented millions of deaths. However, the excessive and irrational use of these drugs has led to their dispersion in the environment, promoting the emergence of antibiotic-resistant bacteria [
1]. These multidrug-resistant bacteria are one of the top threats to global food security, development, and health, according to a report by the WHO [
2].
The use of antibiotics has been on the rise in recent decades. Global antibiotic consumption increased by 46% between 2000 and 2010 [
3]. This large quantity of antibiotics ends up in wastewater, and even if it reaches treatment plants, they fail to eliminate these residues, releasing them into water bodies [
4]. Antibiotic residues in high amounts have been found in effluents; some of these residues are applied as fertilizers to the soil.
This widespread dissemination of antibiotic residues in the environment promotes selective pressure and the spread of resistance genes. Antibiotic resistance can result from mutations that modify the molecular targets of bacterial antibiotics [
5]. Multidrug-resistant pathogen infections are becoming more difficult to treat, which highlights the need for novel antimicrobial compounds with distinct mechanisms of action that can reduce resistance and, if possible, have fewer negative effects on the health of humans, animals, and the environment following the “One Health” philosophy [
6].
To battle strains that are resistant to several drugs, the WHO has created an action plan that centers on the creation of novel antimicrobial products. The search for natural compounds that may be effective as novel antibacterial agents has therefore been quite active.
Numerous natural plant-based medications, such as essential oils (EOs), have been investigated for the treatment and prevention of multidrug-resistant micro-organisms [
7,
8,
9]. Unfortunately, natural products generally have weaker antibiotic activity than common antibiotics; therefore, it is challenging for them to effectively replace current antibiotics in clinical practice. Nonetheless, it has been shown that a small number of plant-derived antimicrobial compounds may work in concert to boost antibiotic action [
10,
11]. By lowering the minimum inhibitory concentration (MIC) of both the antibiotic and the natural product, the synergistic interaction of natural compounds with commercial antibiotics can make the combination as effective as the antibiotic alone while preserving the use of commercial antibiotics (ATBs) [
12,
13].
Since combinations with synergistic effects can lower the likelihood of bacterial resistance emergence while maintaining effective pharmacological outcomes, using lower concentrations of both agents presents significant opportunities for exploring alternative solutions for the treatment of infectious diseases [
14]. Furthermore, as opposed to the side effects from large doses of synthetic medications, this may result in less toxicity from antibiotics [
15].
Polyphenols, alkaloids, carotenoids, terpenes, terpenoids, and sulfur compounds are among the phytochemical substances showing promising antimicrobial activity in vitro [
16]. Global demands for antibiotics are increasing, but antibiotic research programs are significantly inadequate; for example, no new class of Gram-negative antibiotics has been introduced in over 50 years [
17]. Therefore, medicinal plants can represent a rapid and safe source of innovation for new antimicrobial agents.
The
Lamiaceae family of plants is one of the most significant for yielding essential oils as it has antibacterial and antioxidant characteristics. The majority of aromatic plants that are high in essential oils are found in the Mediterranean region, where the extraction of oil is a profitable industry that promotes both ecological and economic growth. Species from the genus
Origanum are among these aromatic and therapeutic plants that are often used as spices due to their abundance of essential oils. Additionally, they display a range of biological activities, the potential of which has been demonstrated by numerous scientific investigations [
18,
19,
20]. The
Lamiaceae family includes the genus
Origanum, which is known as “Oregano” in English. In total, 38 species, including 6 subspecies and 17 hybrids, are found in the Mediterranean, Irano-Turanian, and Euro-Siberian regions. Moreover, it is divided into 10 parts [
20]. Its medicinal use goes back thousands of years because of its many culinary and therapeutic uses. The leaves were used as an antiseptic and for healing skin blemishes. The ancient Greek and Roman empires also employed them to treat various ailments, like indigestion, diarrhea, and asthma [
21]. A common treatment for colds and stomachaches in Greece is still an oregano infusion [
22]. Locally known as “Zaatar” or “Zwi” in Berber, oregano species are highly prized in Morocco. While the Zwi aqueous infusion is used to treat dysentery, colitis, bronchopulmonary diseases, gastric acidity, and gastrointestinal ailments, this plant is traditionally used to cure liver disorders in particular places, such as the Middle Atlas (Jbel-Bouiblane) [
23]. Additionally, several ethnic groups have been using Origanum species in traditional medicine since antiquity to cure and relieve a wide range of ailments. In addition to treating other illnesses, they have antibacterial, anti-inflammatory, antioxidant, anticancer, antifungal, antiviral, and antileishmanial properties [
24,
25,
26].
Finding combinations of these EOs with various classes of commercial antibiotics to find those that have synergistic effects and may allow the use of lower antibiotic doses is the aim of this study.
Origanum compactum and
Origanum elongatum EOs were harvested in different regions of Morocco. The chemical composition of the EOs was identified. We investigated the synergistic combinations by comparing the minimum inhibitory concentrations of the EOs under investigation with nine commonly used antibiotics, both in isolation and in combination, against microbial strains that cause a variety of human diseases. Following the WHO’s list of priority pathogens, the types of bacteria were selected based on their potential severity and capacity for resistance development, as well as their clinical significance as they cause some of the most prevalent diseases in existence today [
27]. Lastly, to gain a deeper comprehension of the fundamental mechanisms behind our experimental endeavors, we will employ drug similarity prediction, molecular dynamics, pharmacokinetics (ADME-Tox), and molecular docking simulations.
4. Discussion
As one of the top three health issues identified by the World Health Organization, antibiotic resistance has become a major factor in worldwide mortality in the twenty-first century [
41]. It happens when bacteria find methods to resist the effects of antibiotics; this is a problem that is made worse by the overuse and poor application of antibiotics in agricultural and medical settings [
42,
43]. Since the 1940s, when modern antibiotics became widely available, the problem has become worse, and almost all bacterial types now exhibit some degree of resistance. Therefore, it is imperative to quickly ascertain alternative materials capable of efficiently suppressing bacteria that have developed resistance to carbapenems. These include methicillin-resistant
Staphylococcus aureus (MRSA),
Acinetobacter baumannii, Enterobacteriaceae (especially
Klebsiella pneumoniae,
Escherichia coli, and
Enterobacter spp.), and
Pseudomonas aeruginosa.
The quest for efficacious antimicrobial therapies has conventionally centered on using the therapeutic characteristics of naturally occurring compounds derived from a wide array of organisms, including bacteria, fungi, plants, algae, and mammals. Exploring plant-based bioactive molecules as possible alternatives to conventional antibiotics has gained traction in recent times. The antibacterial qualities of essential oils, for example, have been the focus of much research due to their ability to combat common human diseases [
44,
45]. Their antibacterial capabilities can be strengthened by mixing them with other substances, such as essential oils and medicines [
46]. To determine the effectiveness of
O. compactum and
O. elongatum essential oils in preventing infection, we developed cultures of bacteria that were resistant to several drugs. We then exposed these strains to the essential oils alone and in combination with antibiotics.
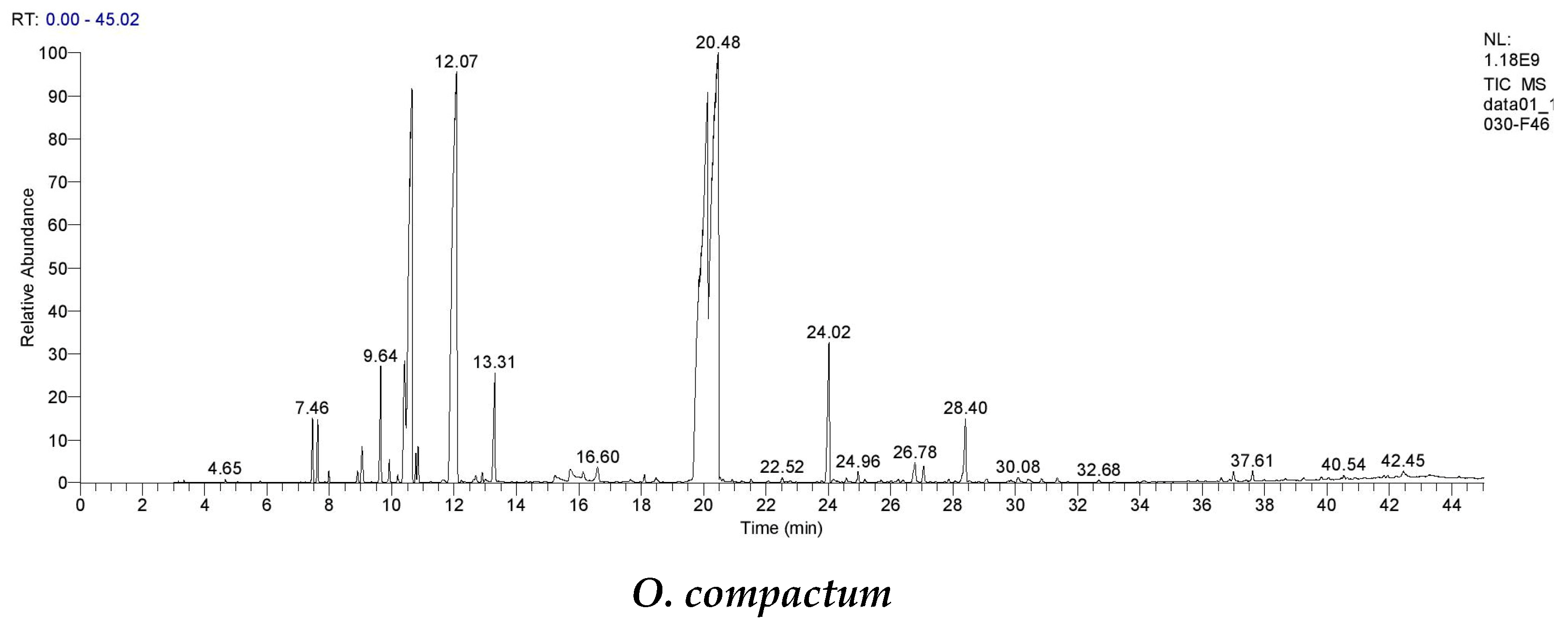
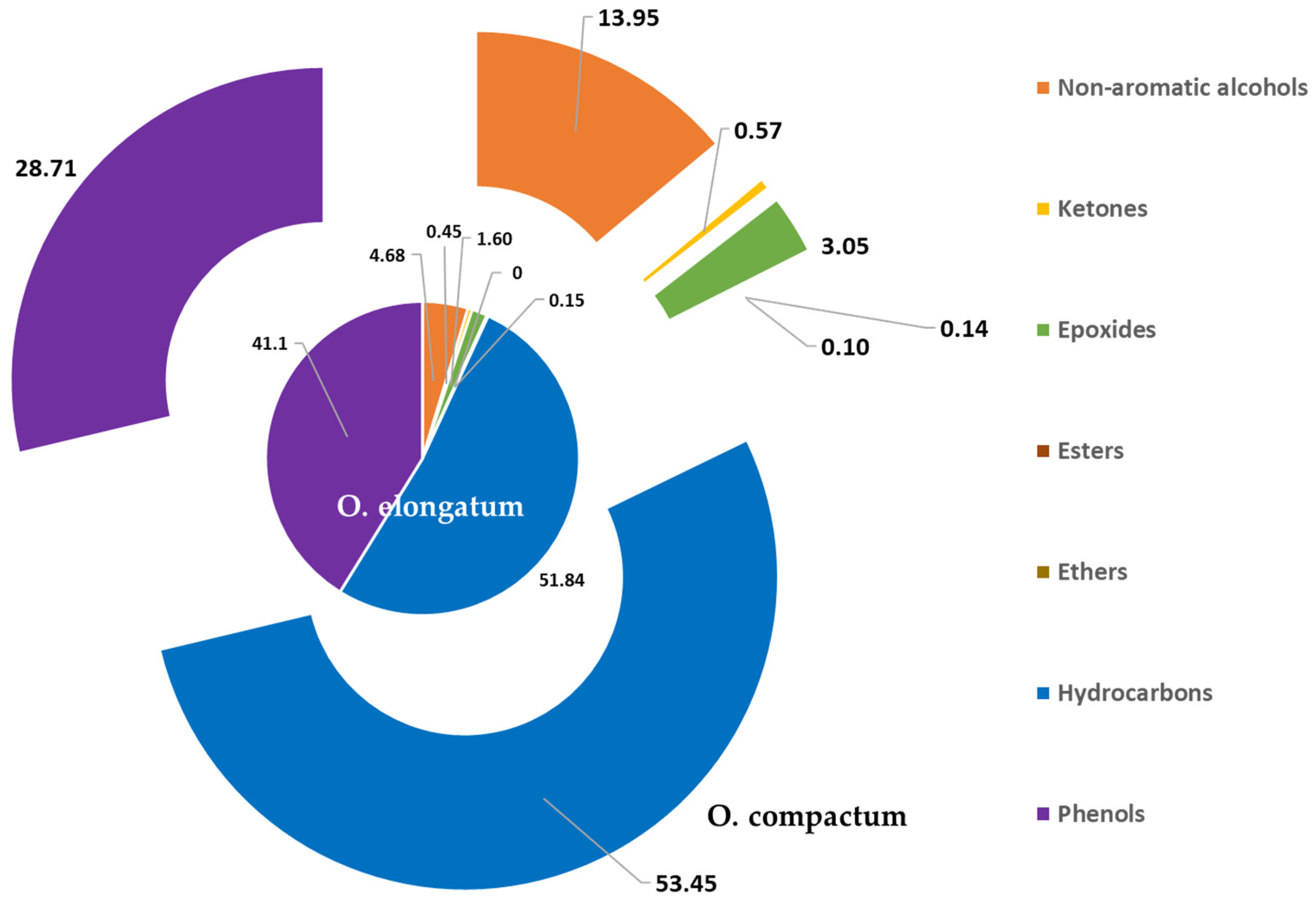
The essential oils under examination underwent a thorough investigation using GC-MS, which revealed their unique chemical signatures. Notably, O. compactum essential oil showed trace components, including €-caryophyllene and linalool together, with a substantial presence of beneficial compounds like carvacrol, γ-terpinene, p-cymene, and thymol. On the other hand, the main constituents of O. elongatum essential oil were p-cymene, thymol, and borneol. These results are consistent with earlier studies on the topic.
Numerous research studies examining the characteristics of
O. compactum’s flowering tops have focused on Morocco. Two studies, in particular, have drawn notice due to their findings. In 2018, Laghmouchi and colleagues conducted an analysis of fourteen essential oils collected from six distinct zones in northern Morocco, spanning diverse geographic areas [
26]. The findings showed considerable quantities of p-cymene (6.69 to 42.64%), thymol (0.16 to 34.29%), and γ-terpinene (2.95 to 22.97%), followed by a high concentration of carvacrol (range from 2.18 to 63.65%). Building on this understanding, a follow-up research study was carried out in 2021 by Ez-zriouli and associates [
47]. Their findings demonstrated the prevalence of carvacrol (72.97%) in
O. compactum essential oil together with significant concentrations of p-cymene (14.5%) and γ-terpinene (6.01%). However, in other countries, several studies have shown that
O. compactum essential oil is characterized by a predominance of carvacrol [
48,
49,
50,
51,
52].
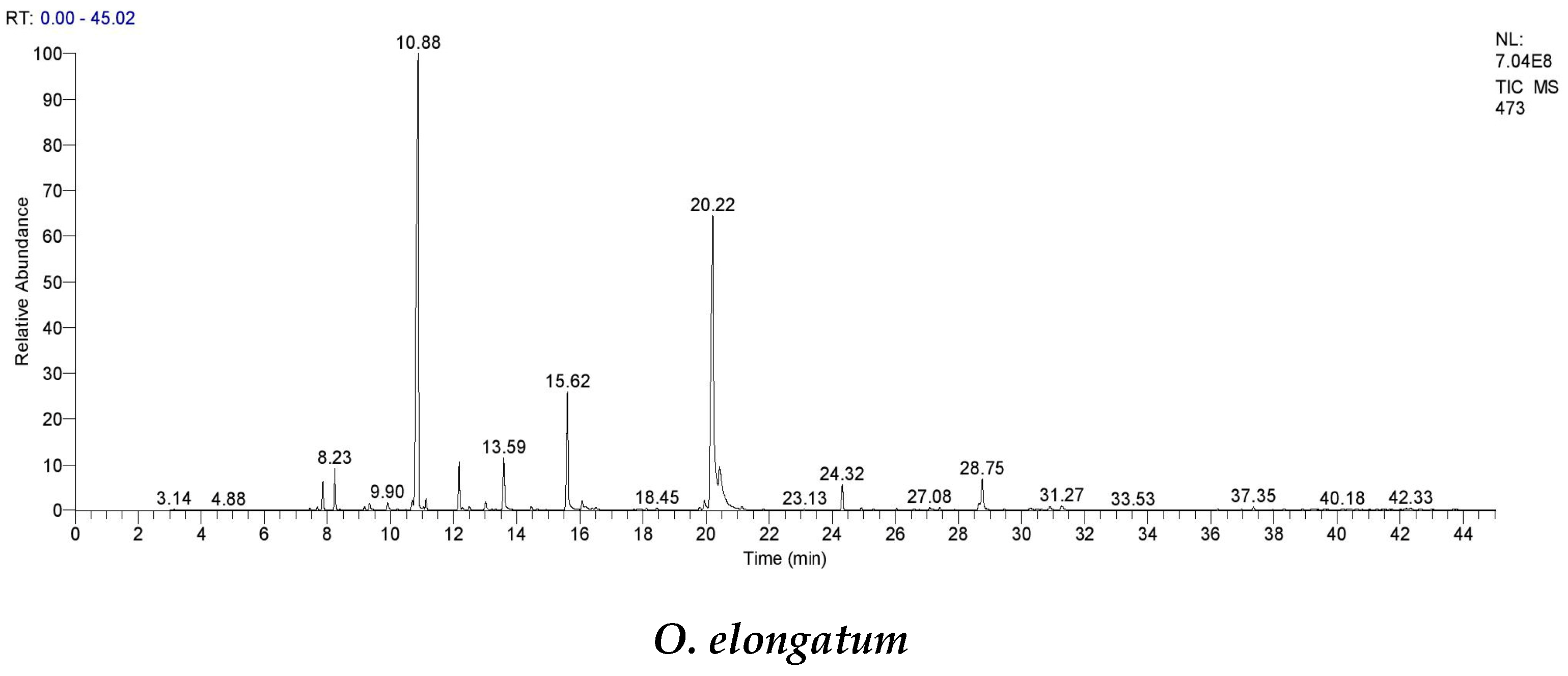
A recent Moroccan research study discovered that
O. elongatum essential oil exists in four chemical forms: carvacrol, carvacrol/thymol, carvacrol/p-cymene, and thymol [
49,
53,
54]. We detected the p-cymene/thymol/borneol form in our investigation. Given that p-cymene is a precursor in the biosynthesis pathways of both carvacrol and thymol, it is plausible that this variance results from harvesting at a different time.
The emergence of antibiotic-resistant bacterial pathogens has rendered most available antibiotics ineffective [
55]. Alternative strategies are therefore necessary to combat drug-resistant bacterial infections. Combined therapies between EOs and conventional antibiotics to enhance their effectiveness appear to be the most effective solution. Indeed, oregano species’ essential oils have shown significant antibacterial activity against all tested bacteria. It is noteworthy that
O. compactum essential oil exhibited the highest antibacterial efficacy, primarily attributable to the high levels of its bioactive constituents, including Carvacrol, γ-Terpinene, p-Cymene, Thymol, and (E)-Caryophyllene. These substances have been praised for their antibacterial properties because of their distinct chemical composition and strong synergy when combined [
56,
57,
58].
Rosato, Scandorieiro, Oumam, and colleagues concluded that oregano essential oils act against bacteria (Gram-positive and Gram-negative) and fungi (yeasts and molds), including multidrug-resistant strains [
59,
60,
61]. In our study, Gram-negative bacteria showed greater sensitivity to both evaluated oregano species than Gram-positive bacteria and fungi, corroborating the results of Lambert’s and Amakran’s studies [
62,
63]. The different sensitivity profiles of the tested strains could be justified by the structure of the cell wall.
Essential oils and antibiotics work synergistically, which is one of the newest tactics to tackle bacterial resistance [
10]. When two substances work better together than they do apart, this is known as a synergistic effect. It also occurs when the observed inhibition of a combination is greater than the expected inhibitions of the individual compounds. The presence of multiple active antibacterial constituents and interactions between different components of both oils are responsible for this synergy phenomenon. These interactions can disrupt the internal membranes of bacteria, increasing the permeability of structures, inhibiting microbial motility, inhibiting microbial ATPase, and/or inhibiting efflux pumps. They can also increase the solubility and/or availability of one or more oil constituents or act on different targets, resulting in enhanced antibacterial effects [
64].
This study has revealed synergistic interactions between oregano essential oils and selected antibiotics. The works of Wendy et al., 2012 [
14,
65,
66,
67], on the use of Origanum essential oils in combination with antibiotics have shown significant synergistic interactions. Our results support these studies by showing that Origanum essential oils have a synergistic effect on antibiotic-resistant bacteria, such as
Klebsiella pneumoniae, Escherichia coli, Pseudomonas aeruginosa, Staphylococcus aureus, and Acinetobacter baumannii, which are frequently responsible for infections linked to healthcare settings. Moreover, the correlation between the composition profile and antibacterial activity can elucidate the synergistic effect. This was especially noticeable when
O. compactum and
O. elongatum essential oils were combined with antibiotics, suggesting that the concentration of monoterpenes in the studied essential oils may be related to their potency.
The FICI values obtained in our study on the 108 interactions examined in vitro, as well as the strong binding affinities to the protein targets studied in silico, reveal promising synergies in the use of oregano essential oils in combination with different antibiotics. When combined with ampicillin against
P. aeruginosa, amoxicillin against strains of
S. aureus, A. baumannii, and K. pneumoniae, and ciprofloxacin against
P. aeruginosa, E. cloacae, and
K. pneumoniae, it has been observed that the essential oils of
O. elongatum and
O. compactum are highly effective. Furthermore, when coupled with the essential oils, ceftriaxone has demonstrated encouraging synergistic benefits against
P. aeruginosa and
E. coli ESBL. Additionally, when combined with tetracycline, the essential oils elicit synergistic activity against strains of
E. cloacae, P. aeruginosa, and
S. aureus. Furthermore, the results of molecular docking demonstrate a noteworthy affinity of these optimal combinations, particularly with EOs high in (E)-caryophyllene and thymol. Our investigation demonstrated a considerable increase in antibacterial activity when ampicillin was combined with the essential oils of
O. compactum and
O. elongatum, with FICI values of 0.188 and 0.066, respectively. Recent research has demonstrated that a potent combination of ampicillin and oregano essential oil is very successful in treating lung infections brought on by bacteria that are resistant to drugs [
68,
69]. The mechanism of action of this potent medication is to directly target and inhibit the transpeptidase enzyme, which is in charge of binding the essential peptidoglycan molecules that makeup bacteria’s cell walls. Ampicillin interferes with this process, making the cell wall brittle and weak, which eventually leads to its rupture. Studies have shown that some essential oils work well when paired with amoxicillin, in addition to displaying encouraging fractional inhibitory concentration index values.
O. compactum and
O. elongatum had FICI values of 0.125 and 0.123, respectively, against
S. aureus. On the other hand,
O. elongatum showed a FICI of 0.375 against
A. baumannii, and
O. compactum demonstrated a FICI of 0.281 against
K. pneumoniae. Amoxicillin works by binding to the transpeptidase enzyme, which breaks the bonds between peptidoglycan molecule-essential components of the bacterial cell wall. Its stability in the acidic environment of the stomach further increases its effectiveness when taken orally. Because of its decreased vulnerability to degradation by beta-lactamase enzymes, it is a more effective treatment for bacteria resistant to antibiotics [
70]. These findings demonstrate how effective essential oils may be used in tandem with medicines to treat bacterial illnesses. Regarding the FICI against
P. aeruginosa (FICI = 0.250),
E. cloacae (FICI = 0.375), and
K. pneumoniae (FICI = 0.129), the combination of
O. elongatum essential oil and ciprofloxacin has shown promising results. The drug ciprofloxacin works by blocking DNA gyrase and topoisomerase IV. It is used for some bacterial infections, including respiratory, urinary, gastrointestinal, and skin infections. Ciprofloxacin inhibits these enzymes’ ability to operate through their interactions, which causes DNA supercoils to accumulate. Compounds present in
O. elongatum essential oils and the antibiotic ciprofloxacin work together to impair DNA repair, transcription, and replication forks, making it impossible for the bacterium to proliferate and survive. While the specific reaction of bacterial cells to fluoroquinolone exposure is still not fully understood, a DNA-damage-induced SOS response is thought to be important [
71,
72]. When combined with ceftriaxone, oregano EO has demonstrated encouraging FICI values against
P. aeruginosa (FICI = 0.188 and 0.125 for
O. elongatum and
O. compactum, respectively) and
E. coli ESBL (FICI = 0.375 for
O. compactum). This partnership suggests that treating bacterial illnesses may be more successful. Ceftriaxone is a more potent alternative against bacterial strains resistant to antibiotics due to its higher stability when compared to first- and second-generation cephalosporins [
73]. It can also be used to treat infections of the central nervous system due to its capacity to cross the blood–brain barrier. Furthermore, it differs from other cephalosporins that are usually only administered intravenously in that they can be administered intramuscularly or intravenously [
74]. The effectiveness of the combination of oregano essential oils (EOs) with tetracycline was studied against different multidrug-resistant bacterial strains. The results obtained revealed promising values of the FICI, highlighting significant synergies between oregano EOs and tetracycline. The found FICI value for
P. aeruginosa (
O. elongatum) is 0.094, 0.258 for
E. cloacae (
O. compactum), and 0.313 and 0.490 for
S. aureus (
O. compactum and
O. elongatum), respectively. To stop messenger RNA from being converted into essential proteins, tetracycline binds to the ribosomal RNA of the bacterium. This inhibition gives tetracycline a bacteriostatic property, preventing bacterial growth without directly killing them, due to its ability to penetrate cell membranes because of its lipophilicity. Furthermore, tetracycline demonstrates particular efficacy against bacteria-producing beta-lactamase enzymes capable of breaking down penicillin antibiotics [
75].
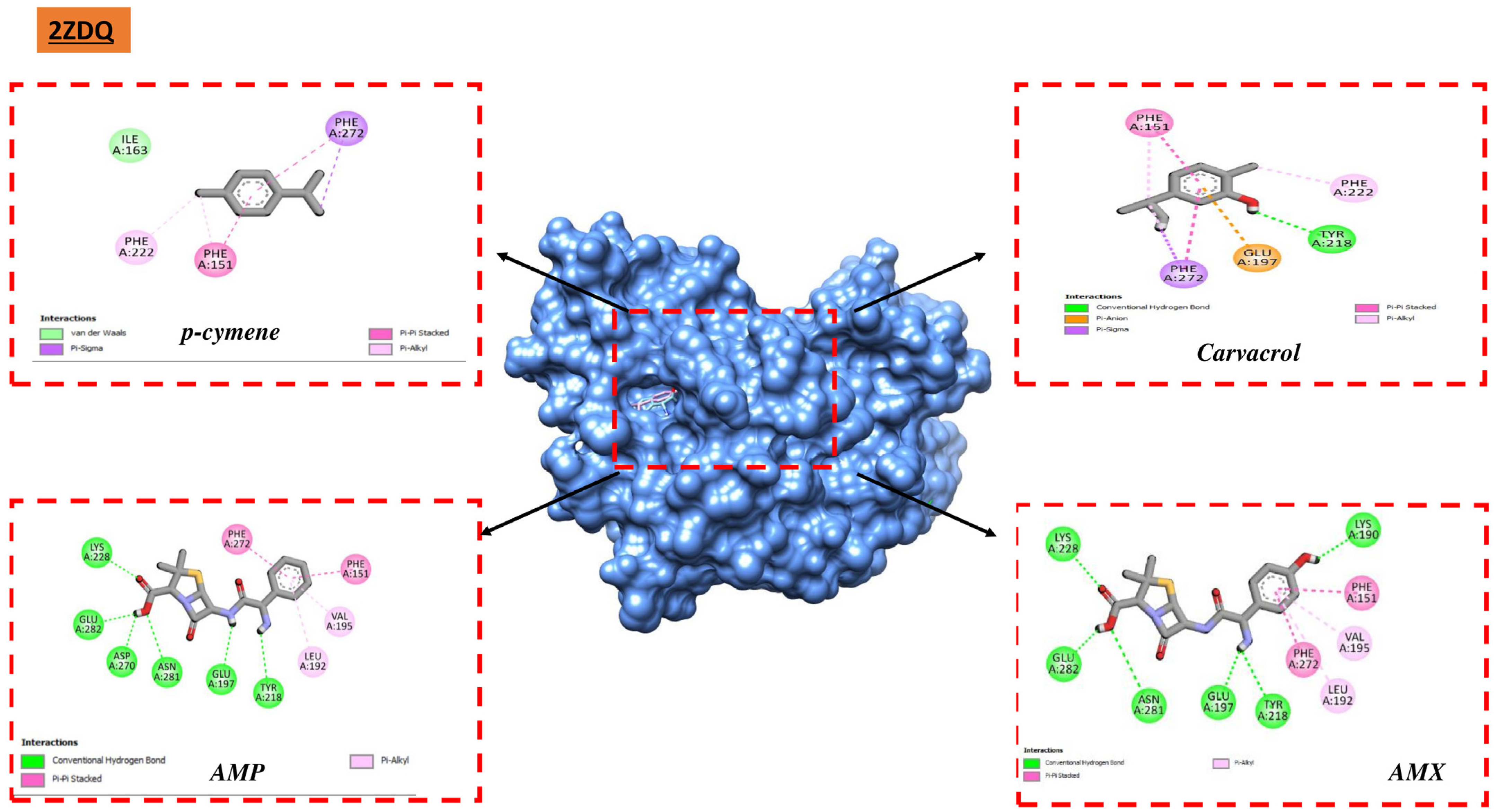
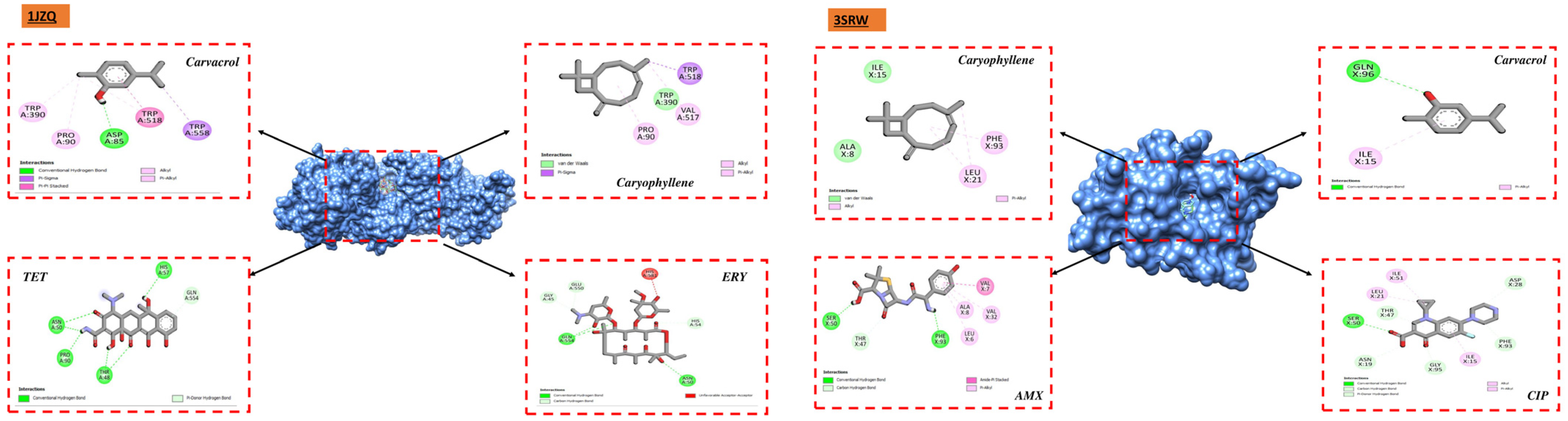
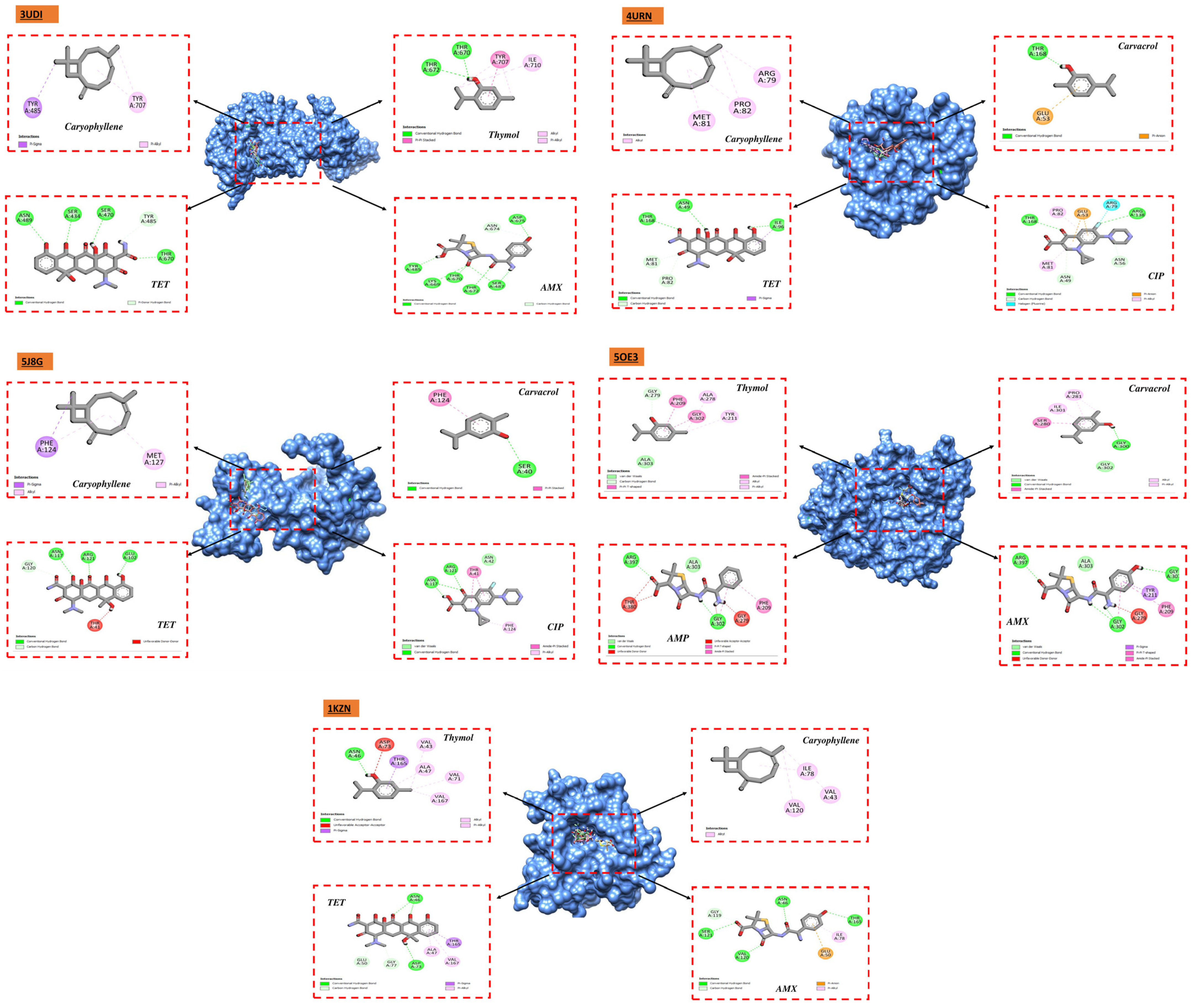
Studies utilizing molecular docking are widely utilized to ascertain the possible interaction between a protein and a ligand. These investigations seem to be beneficial in revealing significant information on the antibacterial qualities of natural sources. They also provide important insights into the complex interactions and possible mechanisms of action at the binding sites of various bacterial proteins [
76]. We carried out docking analyses on the main constituents of the essential oils of the two oregano species under investigation to better comprehend these mechanisms. The receptors that we selected for our study were 1JZQ, 2VEG, 2ZDQ, 3RAE, 3SRW, 3UDI, 1KZN, 5J8G, 5OE3, and 4URN. These essential chemicals are vital in stabilizing the ligand–protein complex and eventually raising binding affinity through the establishment of connections. The compound’s ability to regulate biological systems is ultimately improved by the specific arrangement of hydrogen bonds and corresponding hydrophobic areas, which promotes a deeper and more focused link between the ligand and protein. Furthermore, there may be increased potential for permeability [
77], absorption, and bioavailability in substances with smaller molecular weights, less lipophilicity, and decreased hydrogen bonding capacity [
78,
79].
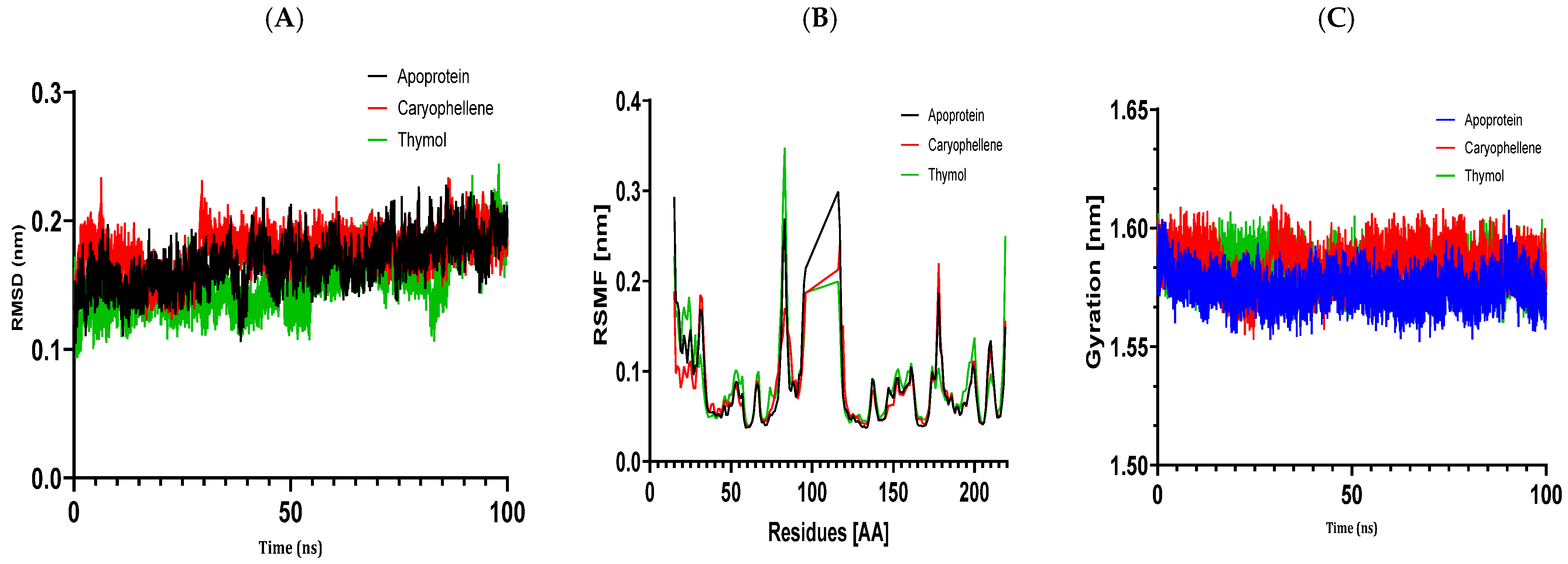
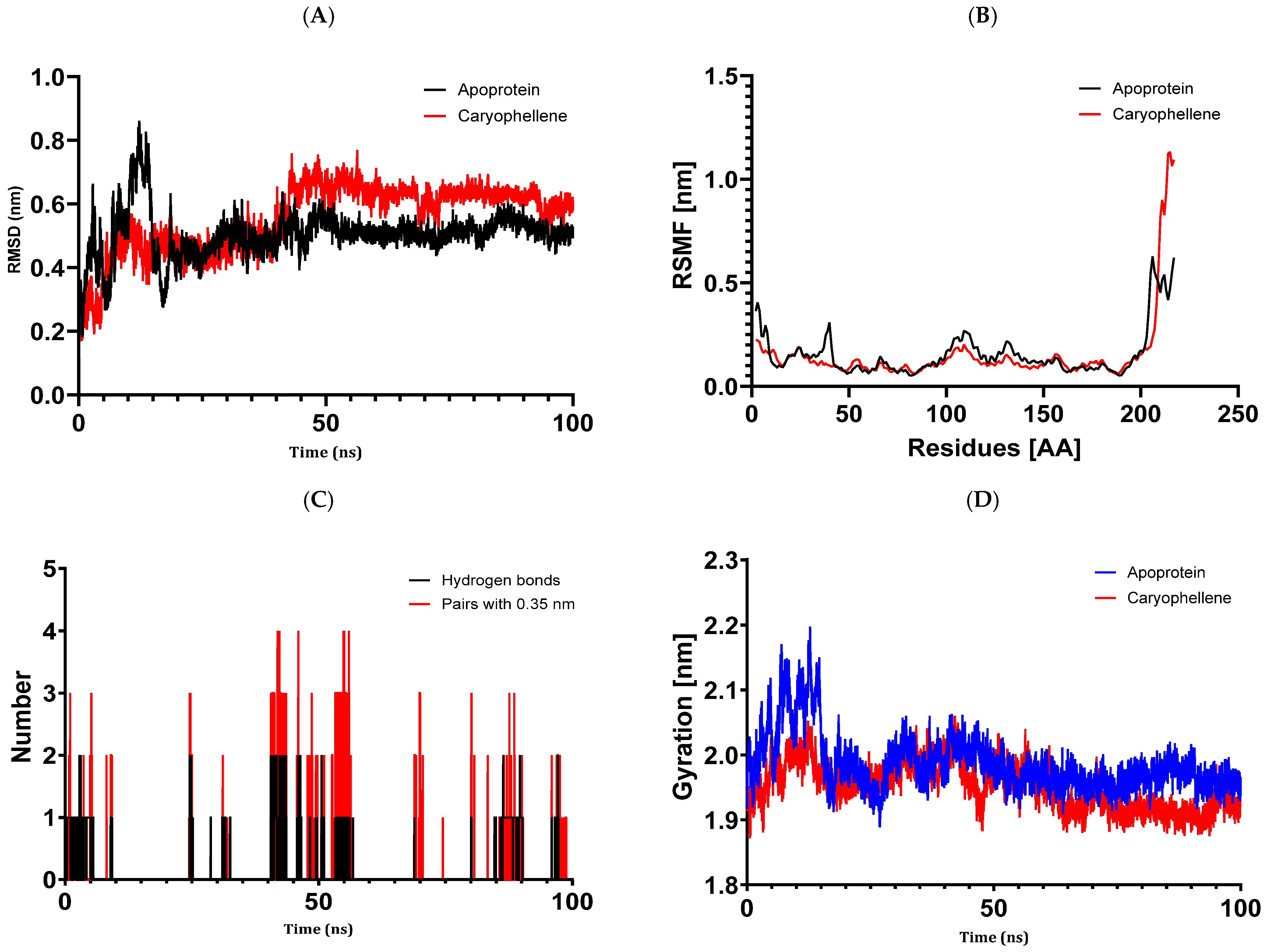
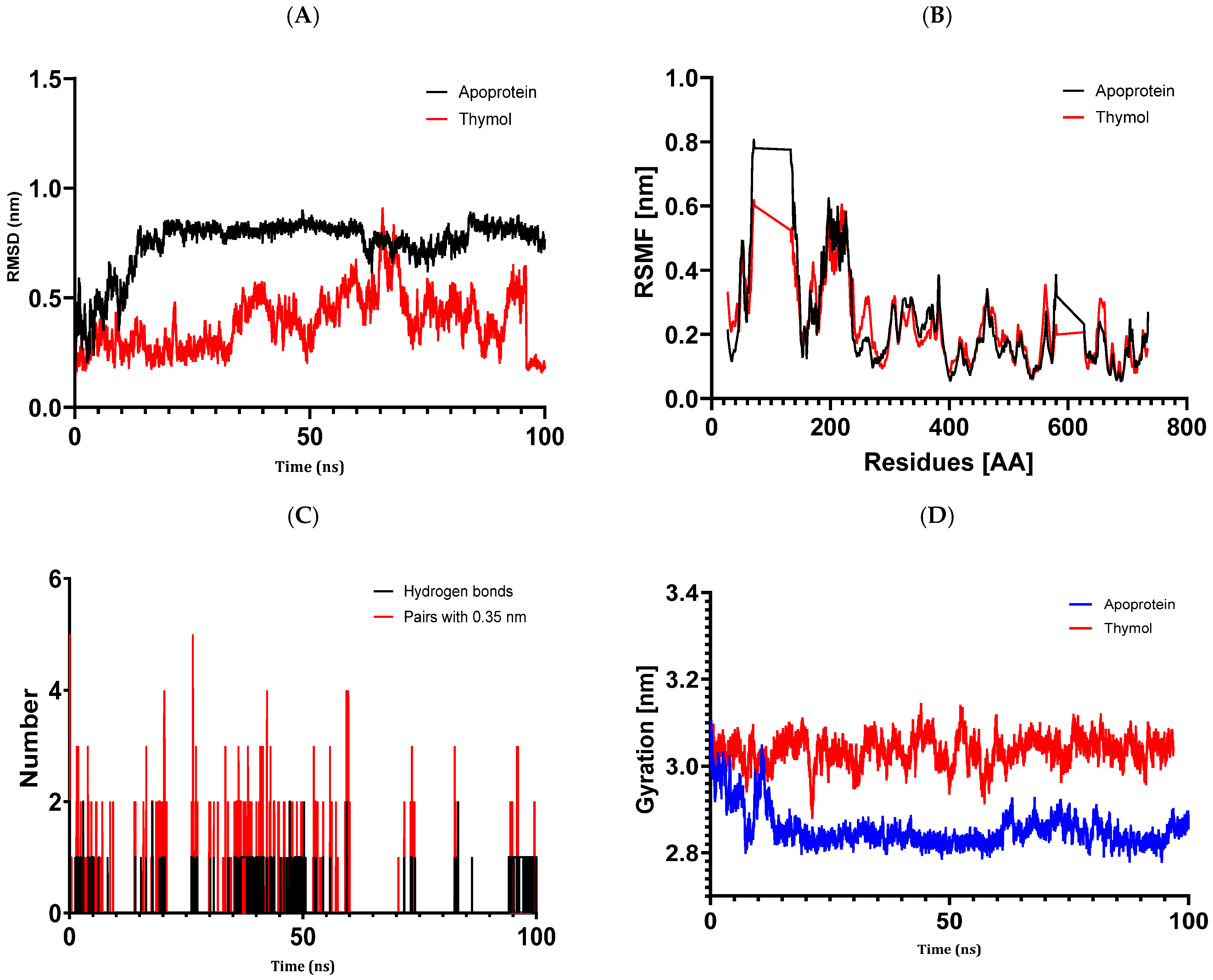
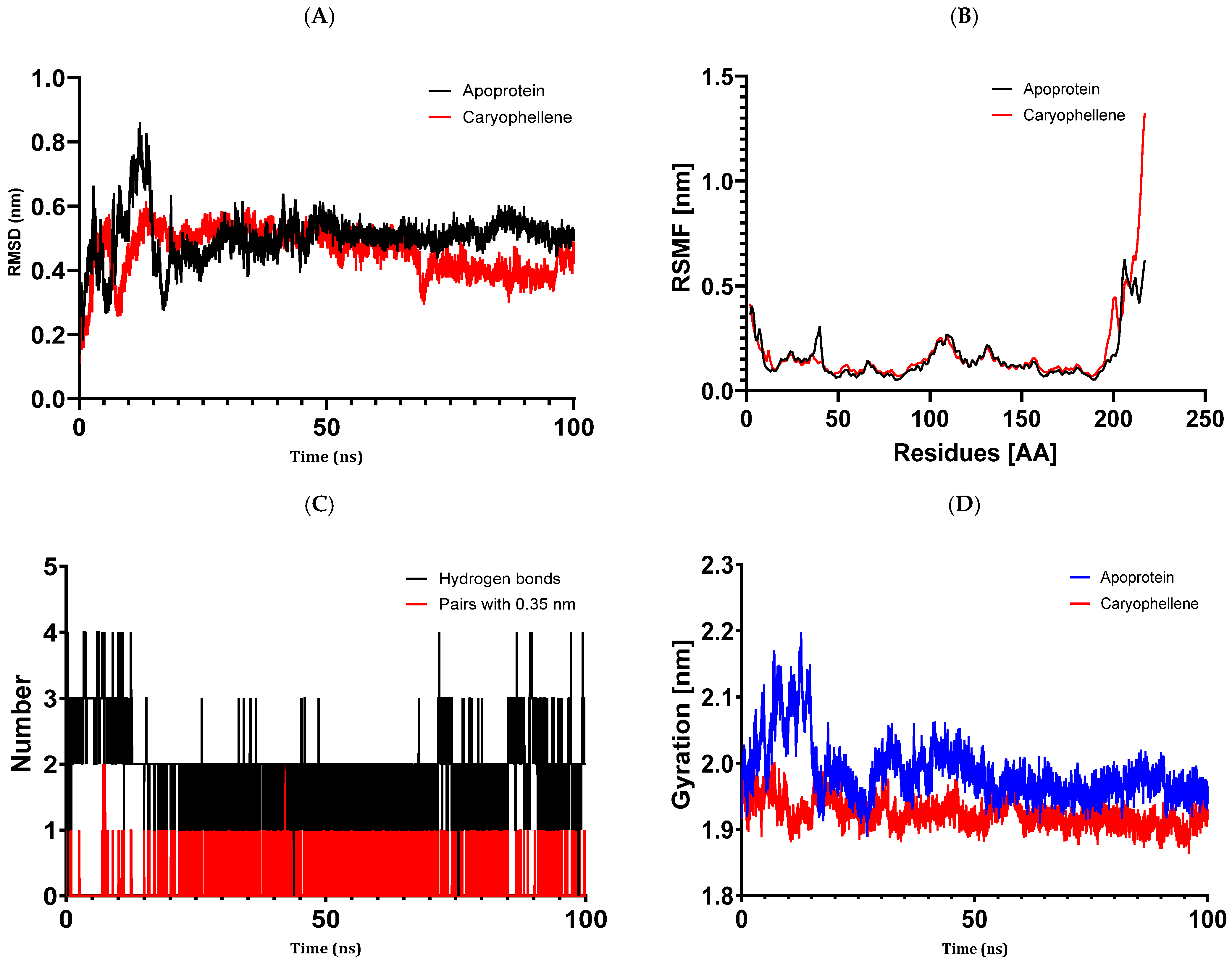
The results of molecular docking investigations and in vitro trials are further validated by the results of molecular dynamics simulations conducted on complex models, such as 5J8G + caryophyllene, 3UDI + thymol, 1JZQ + caryophyllene, 1KZN + caryophyllene, and 1KZN + thymol. This demonstrates the significance of these investigations. They validate that caryophyllene and thymol interact steadily with target proteins, bolstering the theory that these substances are the most potent inhibitors. In light of the possible creation of novel therapeutic antimicrobial medicines, our data provide credence to the theory that these chemicals form dynamically stable complexes throughout 100 ns simulations.
The main constituents of an essential oil as well as the mixture of all of its constituents determine how resistant bacteria are to antimicrobial agents. An essential oil’s ability to combat germs is directly related to its chemical makeup, the main compounds’ functional groups, and the way these components work together. This synergy can arise from several processes, including the inhibition of metabolic pathways or the agents involved, as well as the disruption of the bacterial cell membrane, which makes it easier for additional antibacterial agents to work against the bacterium [
80,
81].
Based on the findings of molecular docking studies and molecular dynamics (MD) simulations, it is possible to explain the potential mechanism of action of compounds from Origanum compactum and Origanum elongatum essential oils (EOs), such as thymol, (E)-caryophyllene, carvacrol, p-cymene, borneol, and linalool, with antibiotics against the multidrug-resistant bacterial strains (K. pneumoniae, E. coli, E. cloacae, P. aeruginosa, S. aureus, and A. baumannii) that are studied.
According to molecular docking studies, thymol, carvacrol, (E)-caryophyllene, and other EO components have substantial interaction with a variety of bacterial protein targets. For instance, D-alanine ligase, an enzyme required for the construction of bacterial cell walls, has demonstrated a significant affinity for carvacrol and p-cymene. The (E)-caryophyllene has demonstrated great affinity with many protein targets, including isoleucyl-tRNA synthetase and dihydropteroate synthase, whereas thymol has demonstrated considerable affinity with protein PBP1a. These interactions imply that these substances may block these proteins, interfering with processes that are necessary for the survival of bacteria.
Molecular dynamics simulations have confirmed the stability of the complexes formed between essential oil compounds and target proteins. For example, in the case of the 1KZN + caryophyllene complex involving the DNA gyrase protein, the root means square deviation (RMSD) fluctuations of the protein remained stable throughout the simulation, indicating a robust interaction between caryophyllene and the protein. The root means square fluctuations and radius of gyration values also remained within acceptable ranges, confirming the stability of the protein–ligand complex. This work demonstrates the important impact that essential oils have on bacterial cells because of their hydrophobic characteristics, which increase the permeability of the cells and cause cytoplasmic components to seep out. Remarkably, some other pathways have also been discovered, such as cell wall breakdown, ATP generation interference, pH level disruption, suppression of protein synthesis, and even DNA damage [
82,
83]. These results point to a potential relationship between the antibiotics under investigation and oregano essential oils, either by focusing on a common mechanism or by making the target location easier to reach.



 ,
, 











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}