
Temperature Dependence of Platelet Metabolism

 ,
,  and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Preparation and Storage of PCs
2.2. Sample Collection
2.3. Assays
2.4. Metabolomics
2.5. Temperature Coefficient Q10 Calculation
2.6. Constraint-Based Metabolic Modeling
3. Results
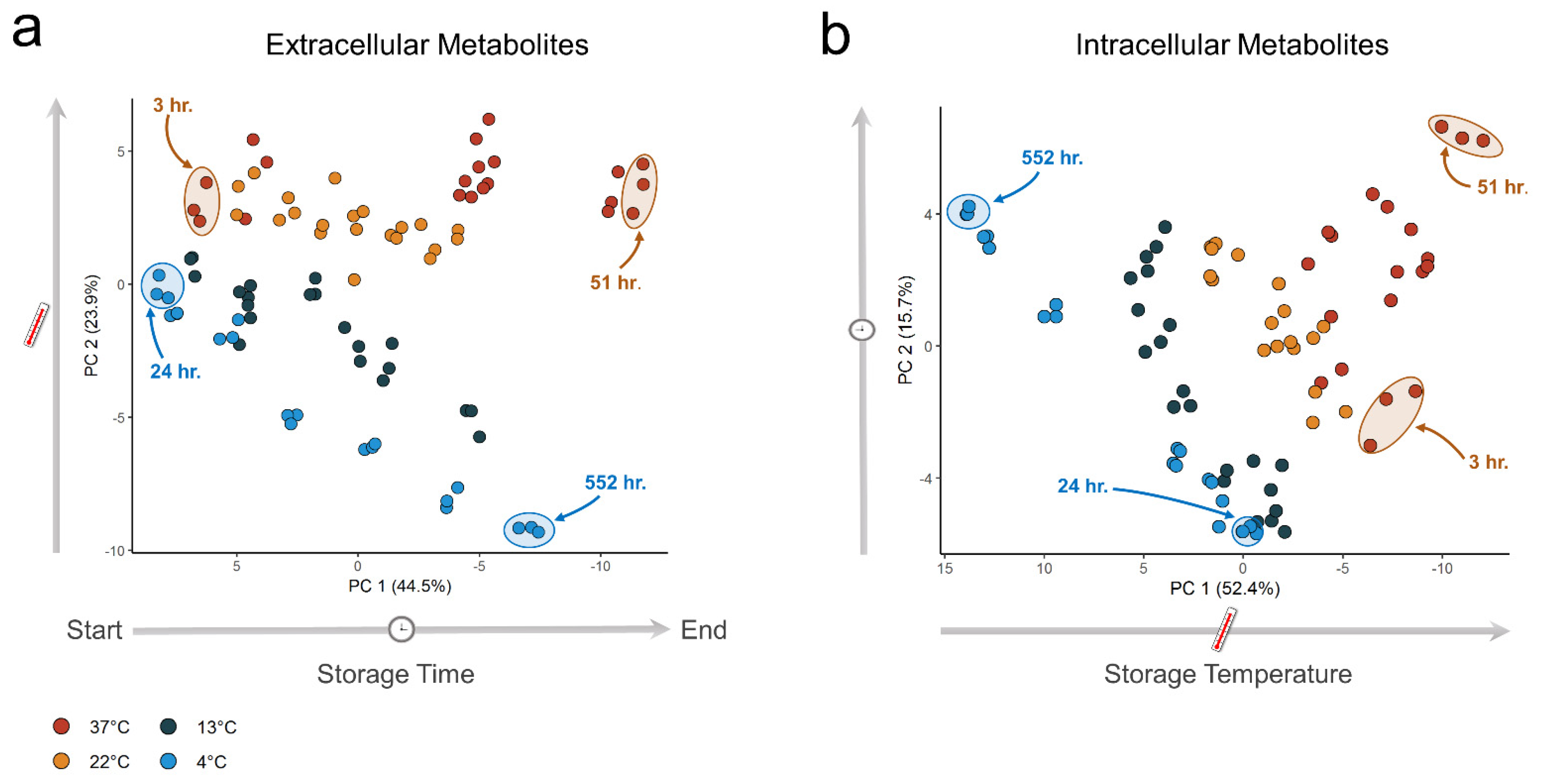
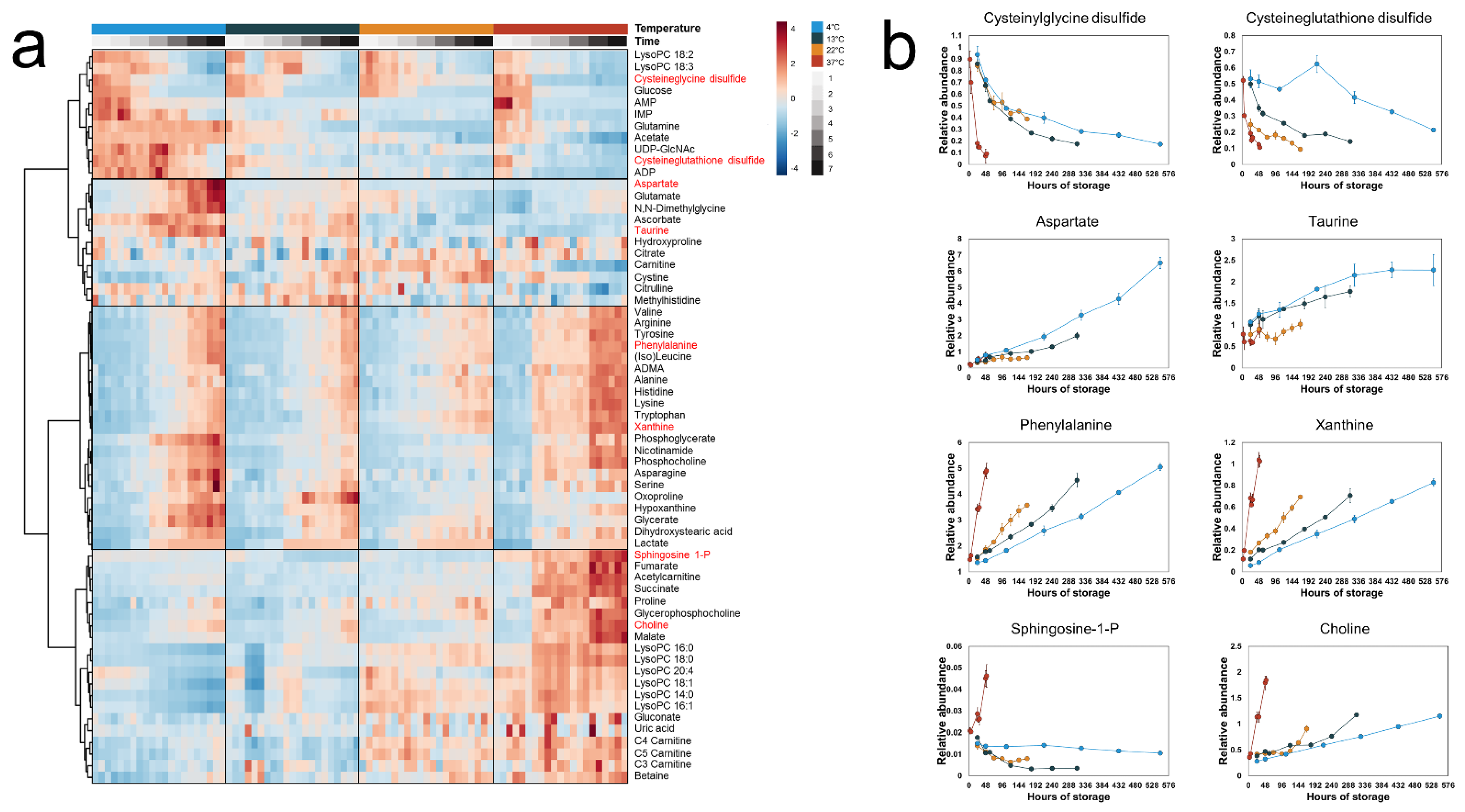
3.1. Metabolite Time–Concentration Profiles Scale Non-Uniformly with Temperature
3.2. A Subset of Extracellular Metabolites Follows an Arrhenius-Type Relationship with Temperature
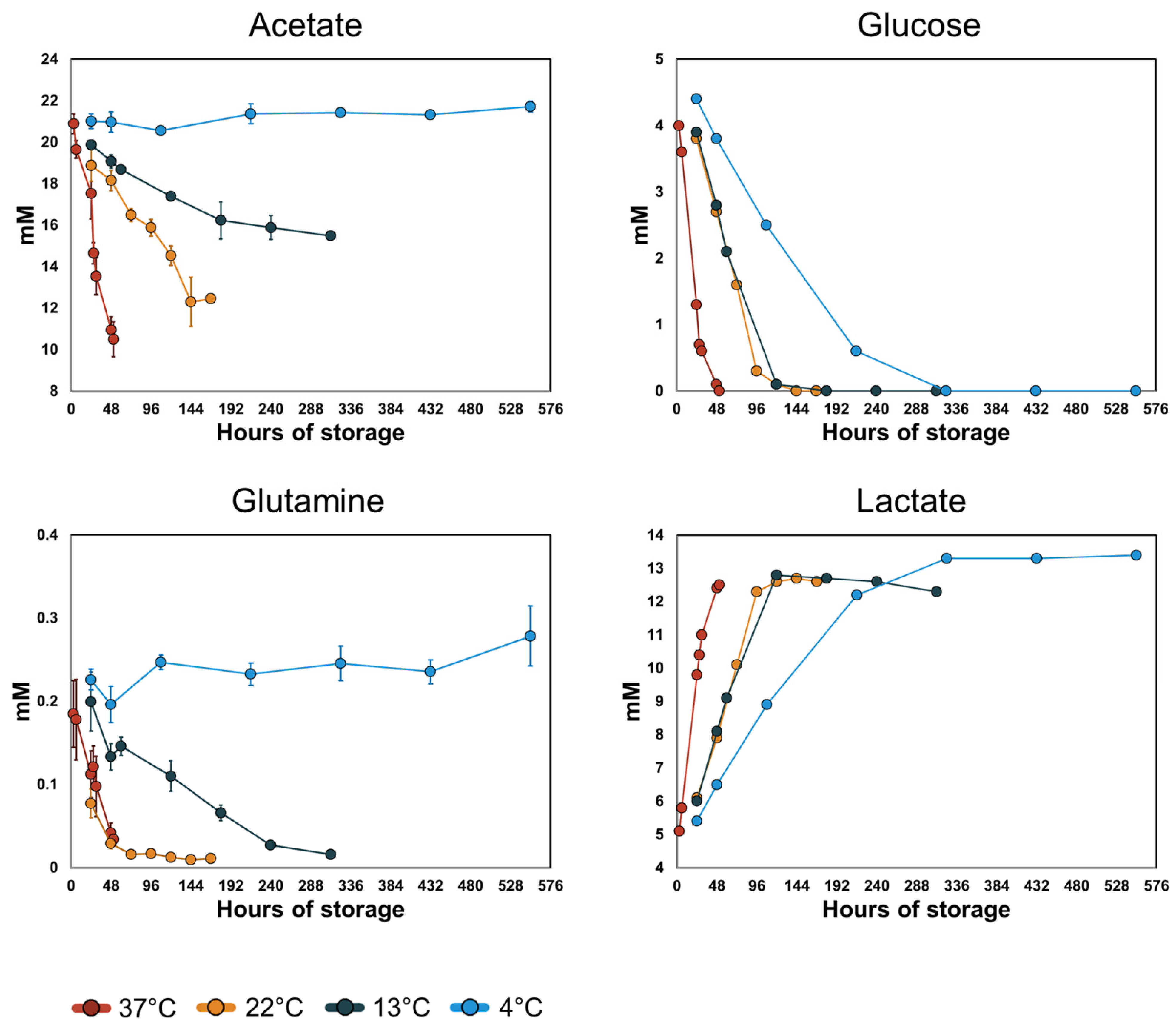
3.3. Acetate and Glutamine Metabolism Is More Sensitive to Temperature Than Glucose Metabolism
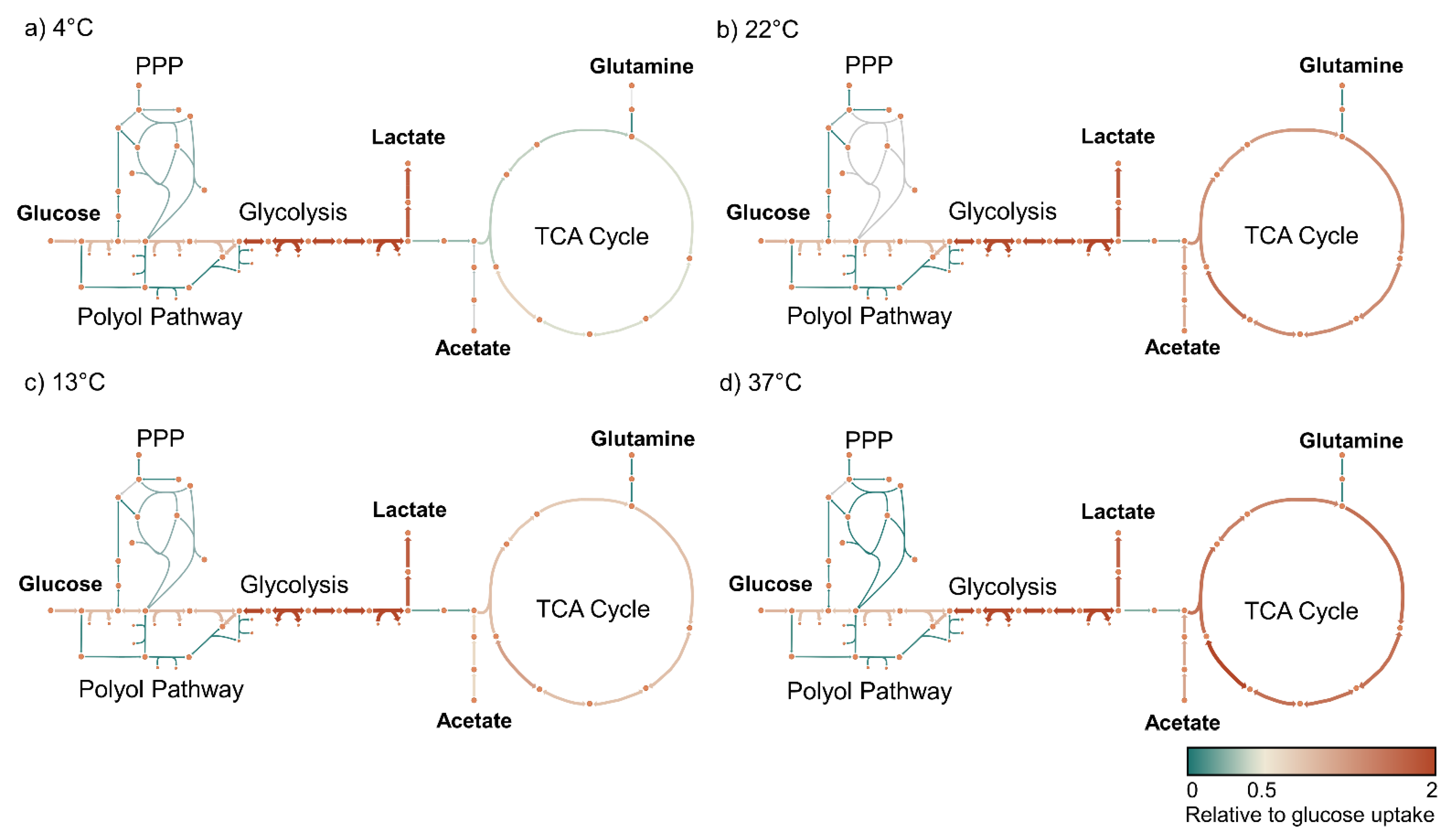
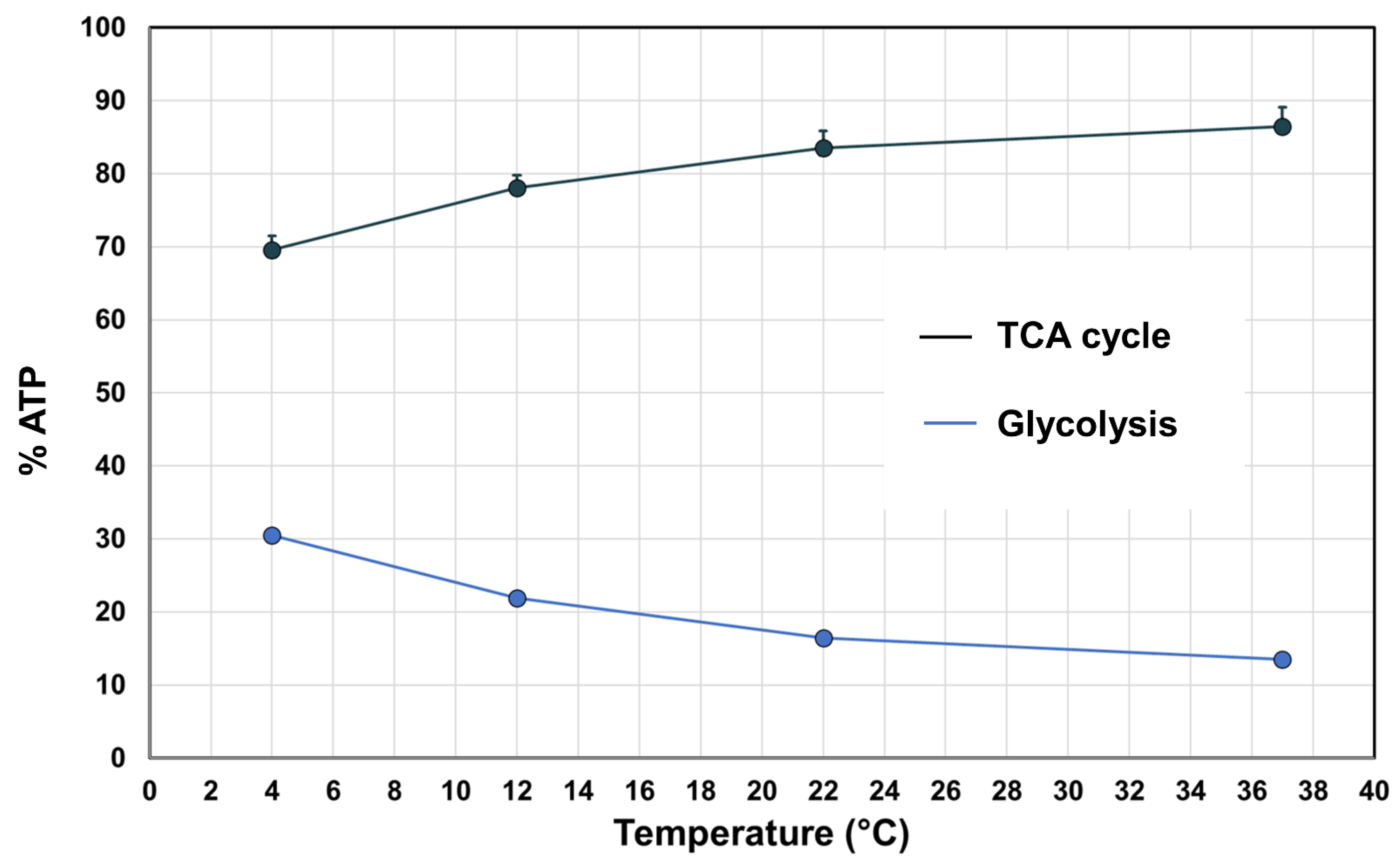
3.4. Metabolic Modeling Reveals Pathway-Specific Temperature Dependence
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Arrhenius, S. Über die Reaktionsgeschwindigkeit bei der Inversion von Rohrzucker durch Säuren. Z. Für Phys. Chem. 1889, 4, 226. [Google Scholar] [CrossRef]
- Laidler, K.J. Unconventional applications of the Arrhenius law. J. Chem. Educ. 1972, 49, 343. [Google Scholar] [CrossRef]
- Stein, R.B.; Gordon, T.; Shriver, J. Temperature dependence of mammalian muscle contractions and ATPase activities. Biophys. J. 1982, 40, 97–107. [Google Scholar] [CrossRef] [PubMed]
- Anderson, G.; Scott, M. Determination of product shelf life and activation energy for five drugs of abuse. Clin. Chem. 1991, 37, 398–402. [Google Scholar] [CrossRef] [PubMed]
- Jorjani, P.; Ozturk, S.S. Effects of cell density and temperature on oxygen consumption rate for different mammalian cell lines. Biotechnol. Bioeng. 1999, 64, 349–356. [Google Scholar]
- Giacomin, M.; Schulte, P.M.; Wood, C.M. Differential Effects of Temperature on Oxygen Consumption and Branchial Fluxes of Urea, Ammonia, and Water in the Dogfish Shark (Squalus acanthias suckleyi). Physiol. Biochem. Zool. PBZ 2017, 90, 627–637. [Google Scholar] [CrossRef]
- Yurkovich, J.T.; Zielinski, D.C.; Yang, L.; Paglia, G.; Rolfsson, O.; Sigurjonsson, O.E.; Broddrick, J.T.; Bordbar, A.; Wichuk, K.; Brynjolfsson, S.; et al. Quantitative time-course metabolomics in human red blood cells reveal the temperature dependence of human metabolic networks. J. Biol. Chem. 2017, 292, 19556–19564. [Google Scholar] [CrossRef]
- Hegarty, T.W. Temperature Coefficient (Q10), Seed Germination and Other Biological Processes. Nature 1973, 243, 305–306. [Google Scholar] [CrossRef]
- Cohen Stuart, C.P. A study of temperature-coefficients and van‘t Hoff’s rule. K. Ned. Akad. Van Wet. Proc. Ser. B Phys. Sci. 1912, 14, 1159–1173. [Google Scholar]
- Chaui-Berlinck, J.G.; Navas, C.A.; Monteiro, L.H.; Bicudo, J.E. Temperature effects on a whole metabolic reaction cannot be inferred from its components. Proc. Biol. Sci. 2004, 271, 1415–1419. [Google Scholar] [CrossRef]
- Chaui-Berlinck, J.G.; Monteiro, L.H.; Navas, C.A.; Bicudo, J.E. Temperature effects on energy metabolism: A dynamic system analysis. Proc. Biol. Sci. 2002, 269, 15–19. [Google Scholar] [CrossRef]
- Donhoffer, S. The regulation of energy metabolism and van’t Hoff’s rule in the homeotherm animal. Helgol. Mar. Res. 1966, 14, 541–558. [Google Scholar] [CrossRef]
- Gillooly, J.F.; Brown, J.H.; West, G.B.; Savage, V.M.; Charnov, E.L. Effects of Size and Temperature on Metabolic Rate. Science 2001, 293, 2248–2251. [Google Scholar] [CrossRef] [PubMed]
- Orth, J.D.; Thiele, I.; Palsson, B. What is flux balance analysis? Nat. Biotechnol. 2010, 28, 245–248. [Google Scholar] [CrossRef] [PubMed]
- Bordbar, A.; Yurkovich, J.T.; Paglia, G.; Rolfsson, O.; Sigurjónsson, Ó.E.; Palsson, B.O. Elucidating dynamic metabolic physiology through network integration of quantitative time-course metabolomics. Sci. Rep. 2017, 7, 46249. Available online: https://www.nature.com/articles/srep46249#supplementary-information (accessed on 12 December 2018). [CrossRef] [PubMed]
- Schubert, P.; Devine, D.V. De novo protein synthesis in mature platelets: A consideration for transfusion medicine. Vox Sang. 2010, 99, 112–122. [Google Scholar] [CrossRef] [PubMed]
- Zimmerman, G.A.; Weyrich, A.S. Signal-dependent protein synthesis by activated platelets: New pathways to altered phenotype and function. Arterioscler. Thromb. Vasc. Biol. 2008, 28, s17–s24. [Google Scholar] [CrossRef] [PubMed]
- Guppy, M.; Whisson, M.E.; Sabaratnam, R.; Withers, P.; Brand, K. Alternative fuels for platelet storage: A metabolic study. Vox Sang. 1990, 59, 146–152. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Mhaskar, R.; Grossman, B.J.; Kaufman, R.M.; Tobian, A.A.; Kleinman, S.; Gernsheimer, T.; Tinmouth, A.T.; Djulbegovic, B. Platelet transfusion: A systematic review of the clinical evidence. Transfusion 2015, 55, 1116–1127, quiz 1115. [Google Scholar] [CrossRef]
- Devine, D.V.; Serrano, K. The platelet storage lesion. Clin. Lab. Med. 2010, 30, 475–487. [Google Scholar] [CrossRef]
- Murphy, S.; Gardner, F.H. Effect of storage temperature on maintenance of platelet viability—Deleterious effect of refrigerated storage. N. Engl. J. Med. 1969, 280, 1094–1098. [Google Scholar] [CrossRef]
- Marini, I.; Aurich, K.; Jouni, R.; Nowak-Harnau, S.; Hartwich, O.; Greinacher, A.; Thiele, T.; Bakchoul, T. Cold storage of platelets in additive solution: The impact of residual plasma in apheresis platelet concentrates. Haematologica 2019, 104, 207–214. [Google Scholar] [CrossRef]
- Stubbs, J.R.; Tran, S.A.; Emery, R.L.; Hammel, S.A.; Haugen, D.A.L.; Zielinski, M.D.; Zietlow, S.P.; Jenkins, D. Cold platelets for trauma-associated bleeding: Regulatory approval, accreditation approval, and practice implementation—Just the “tip of the iceberg”. Transfusion 2017, 57, 2836–2844. [Google Scholar] [CrossRef]
- Wood, B.; Johnson, L.; Hyland, R.A.; Marks, D.C. Maximising platelet availability by delaying cold storage. Vox Sang. 2018, 113, 403–411. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Devine, D.V. The Missing Pieces to the Cold-Stored Platelet Puzzle. Int. J. Mol. Sci. 2022, 23, 1100. [Google Scholar] [CrossRef] [PubMed]
- Kilkson, H.; Holme, S.; Murphy, S. Platelet metabolism during storage of platelet concentrates at 22 degrees C. Blood 1984, 64, 406–414. [Google Scholar] [CrossRef]
- Murphy, S.; Shimizu, T.; Miripol, J. Platelet storage for transfusion in synthetic media: Further optimization of ingredients and definition of their roles. Blood 1995, 86, 3951–3960. [Google Scholar] [CrossRef]
- Murphy, S. The oxidation of exogenously added organic anions by platelets facilitates maintenance of pH during their storage for transfusion at 22 degrees C. Blood 1995, 85, 1929–1935. [Google Scholar] [CrossRef]
- Paglia, G.; Sigurjonsson, O.E.; Rolfsson, O.; Valgeirsdottir, S.; Hansen, M.B.; Brynjolfsson, S.; Gudmundsson, S.; Palsson, B.O. Comprehensive metabolomic study of platelets reveals the expression of discrete metabolic phenotypes during storage. Transfusion 2014, 54, 2911–2923. [Google Scholar] [CrossRef] [PubMed]
- Paglia, G.; Sigurjonsson, O.E.; Rolfsson, O.; Hansen, M.B.; Brynjolfsson, S.; Gudmundsson, S.; Palsson, B.O. Metabolomic analysis of platelets during storage: A comparison between apheresis- and buffy coat-derived platelet concentrates. Transfusion 2015, 55, 301–313. [Google Scholar] [CrossRef]
- Johannsson, F.; Guðmundsson, S.; Paglia, G.; Guðmundsson, S.; Palsson, B.; Sigurjonsson, O.E.; Rolfsson, O. Systems analysis of metabolism in platelet concentrates during storage in platelet additive solution. Biochem. J. 2018, 475, 2225–2240. [Google Scholar] [CrossRef] [PubMed]
- Thomas, A.; Rahmanian, S.; Bordbar, A.; Palsson, B.O.; Jamshidi, N. Network reconstruction of platelet metabolism identifies metabolic signature for aspirin resistance. Sci. Rep. 2014, 4, 3925. [Google Scholar] [CrossRef] [PubMed]
- Sandgren, P.; Shanwell, A.; Gulliksson, H. Storage of buffy coat-derived platelets in additive solutions: In vitro effects of storage at 4 degrees C. Transfusion 2006, 46, 828–834. [Google Scholar] [CrossRef] [PubMed]
- NasrEldin, E. Effect of cold storage on platelets quality stored in a small containers: Implications for pediatric transfusion. Pediatr. Hematol. Oncol. J. 2017, 2, 29–34. [Google Scholar] [CrossRef]
- Johnson, L.; Tan, S.; Wood, B.; Davis, A.; Marks, D.C. Refrigeration and cryopreservation of platelets differentially affect platelet metabolism and function: A comparison with conventional platelet storage conditions. Transfusion 2016, 56, 1807–1818. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.W.; Serrano, K.; Stefanoni, D.; D’Alessandro, A.; Devine, D.V. In Vitro Characterization and Metabolomic Analysis of Cold-Stored Platelets. J. Proteome Res. 2021, 20, 2251–2265. [Google Scholar] [CrossRef] [PubMed]
- D’Alessandro, A.; Thomas, K.A.; Stefanoni, D.; Gamboni, F.; Shea, S.M.; Reisz, J.A.; Spinella, P.C. Metabolic phenotypes of standard and cold-stored platelets. Transfusion 2020, 60 (Suppl. S3), S96–S106. [Google Scholar] [CrossRef] [PubMed]
- Xia, J.; Wishart, D.S. Using MetaboAnalyst 3.0 for Comprehensive Metabolomics Data Analysis. Curr. Protoc. Bioinform. 2016, 55, 14.10.1–14.10.91. [Google Scholar] [CrossRef]
- Schellenberger, J.; Palsson, B.O. Use of randomized sampling for analysis of metabolic networks. J. Biol. Chem. 2009, 284, 5457–5461. [Google Scholar] [CrossRef]
- Heirendt, L.; Arreckx, S.; Pfau, T.; Mendoza, S.N.; Richelle, A.; Heinken, A.; Haraldsdóttir, H.S.; Wachowiak, J.; Keating, S.M.; Vlasov, V.; et al. Creation and analysis of biochemical constraint-based models using the COBRA Toolbox v.3.0. Nat. Protoc. 2019, 14, 639–702. [Google Scholar] [CrossRef]
- Worley, B.; Powers, R. Multivariate Analysis in Metabolomics. Curr. Metabolomics 2013, 1, 92–107. [Google Scholar] [CrossRef]
- European Directorate for the Quality of Medicines & HealthCare. Guide to the Preparation, Use and Quality Assurance of Blood Components: Recommendation no. R (95) 15; European Directorate for the Quality of Medicines & HealthCare: Strasbourg, France, 2020. [Google Scholar]
- Thiele, I.; Swainston, N.; Fleming, R.M.T.; Hoppe, A.; Sahoo, S.; Aurich, M.K.; Haraldsdottir, H.; Mo, M.L.; Rolfsson, O.; Stobbe, M.D.; et al. A community-driven global reconstruction of human metabolism. Nat. Biotechnol. 2013, 31, 419. Available online: https://www.nature.com/articles/nbt.2488#supplementary-information (accessed on 20 December 2018). [CrossRef]
- Bordbar, A.; Monk, J.M.; King, Z.A.; Palsson, B.O. Constraint-based models predict metabolic and associated cellular functions. Nat. Rev. Genet. 2014, 15, 107–120. [Google Scholar] [CrossRef]
- Ravi, S.; Chacko, B.; Sawada, H.; Kramer, P.A.; Johnson, M.S.; Benavides, G.A.; O’Donnell, V.; Marques, M.B.; Darley-Usmar, V.M. Metabolic plasticity in resting and thrombin activated platelets. PLoS ONE 2015, 10, e0123597. [Google Scholar] [CrossRef]
- Koebmann, B.J.; Westerhoff, H.V.; Snoep, J.L.; Nilsson, D.; Jensen, P.R. The glycolytic flux in Escherichia coli is controlled by the demand for ATP. J. Bacteriol. 2002, 184, 3909–3916. [Google Scholar] [CrossRef]
- Aibibula, M.; Naseem, K.M.; Sturmey, R.G. Glucose metabolism and metabolic flexibility in blood platelets. J. Thromb. Haemost. JTH 2018, 16, 2300–2314. [Google Scholar] [CrossRef] [PubMed]
- Guppy, M.; Abas, L.; Arthur, P.G.; Whisson, M.E. The Pasteur effect in human platelets: Implications for storage and metabolic control. Br. J. Haematol. 1995, 91, 752–757. [Google Scholar] [CrossRef] [PubMed]
- Bynum, J.A.; Meledeo, M.A.; Getz, T.M.; Rodriguez, A.C.; Aden, J.K.; Cap, A.P.; Pidcoke, H.F. Bioenergetic profiling of platelet mitochondria during storage: 4 degrees C storage extends platelet mitochondrial function and viability. Transfusion 2016, 56 (Suppl. S1), S76–S84. [Google Scholar] [CrossRef] [PubMed]
- Murphy, M.P. How mitochondria produce reactive oxygen species. Biochem. J. 2009, 417, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Aledo, J.C. Life-history Constraints on the Mechanisms that Control the Rate of ROS Production. Curr. Genom. 2014, 15, 217–230. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Metabolite | Q10 | R2 |
|---|---|---|
| Alanine | 2.37 | 0.99 |
| Arginine | 2.02 | 0.92 |
| Cysteineglycine disulfide | 2.14 | 0.94 |
| Glucose | 1.73 | 0.99 |
| Glycerate | 1.54 | 0.89 |
| Histidine | 2.30 | 0.98 |
| Hypoxanthine | 1.77 | 0.84 |
| (Iso)Leucine | 2.04 | 0.95 |
| Lactate | 1.70 | 0.98 |
| Lysine | 2.25 | 0.94 |
| Malate | 3.04 | 0.93 |
| Nicotinamide | 2.01 | 0.83 |
| Phenylalanine | 2.05 | 0.94 |
| Phosphocholine | 2.16 | 0.80 |
| sn-Glycero-3-phosphocholine | 2.50 | 0.98 |
| Tryptophan | 2.15 | 0.96 |
| Tyrosine | 2.15 | 0.94 |
| Valine | 2.18 | 0.97 |
| Xanthine | 2.23 | 0.94 |
| Subsystem | Q10 | R2 | Number of Reactions |
|---|---|---|---|
| Alanine and aspartate metabolism | 2.28 (1.97–2.80) | 0.91 (0.90–0.93) | 5 (5) |
| Citric acid cycle | 2.58 (1.97–2.70) | 0.97 (0.90–0.98) | 9 (13) |
| Fatty acid oxidation | 2.21 (2.22–2.55) | 0.91 (0.76–0.98) | 8 (16) |
| Glycerophospholipid metabolism | 2.23 (2.14–2.33) | 0.72 (0.71–0.95) | 37 (101) |
| Glycolysis/gluconeogenesis | 1.76 (1.72–1.77) | 0.98 (0.96–0.99) | 12 (18) |
| Inositol Phosphate metabolism | 2.21 (2.21–2.22) | 0.95 (0.93–0.96) | 4 (124) |
| Nucleotide interconversion | 2.26 (2.22–2.70) | 0.97 (0.95–0.98) | 4 (21) |
| Oxidative phosphorylation | 2.46 (2.27–2.47) | 0.98 (0.98–0.98) | 5 (5) |
| Triacylglycerol synthesis | 2.22 (2.21–2.25) | 0.82 (0.73–0.90) | 7 (43) |
| Other | 2.27 (1.73–3.65) | 0.95 (0.74–0.99) | 26 (184) |
| Total | 2.23 (1.72–3.65) | 0.90 (0.71–0.99) | 117 (530) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jóhannsson, F.; Yurkovich, J.T.; Guðmundsson, S.; Sigurjónsson, Ó.E.; Rolfsson, Ó. Temperature Dependence of Platelet Metabolism. Metabolites 2024, 14, 91. https://doi.org/10.3390/metabo14020091
Jóhannsson F, Yurkovich JT, Guðmundsson S, Sigurjónsson ÓE, Rolfsson Ó. Temperature Dependence of Platelet Metabolism. Metabolites. 2024; 14(2):91. https://doi.org/10.3390/metabo14020091
Chicago/Turabian StyleJóhannsson, Freyr, James T. Yurkovich, Steinn Guðmundsson, Ólafur E. Sigurjónsson, and Óttar Rolfsson. 2024. "Temperature Dependence of Platelet Metabolism" Metabolites 14, no. 2: 91. https://doi.org/10.3390/metabo14020091
APA StyleJóhannsson, F., Yurkovich, J. T., Guðmundsson, S., Sigurjónsson, Ó. E., & Rolfsson, Ó. (2024). Temperature Dependence of Platelet Metabolism. Metabolites, 14(2), 91. https://doi.org/10.3390/metabo14020091