
Multi-Modal Investigation of Metabolism in Murine Breast Cancer Cell Lines Using Fluorescence Lifetime Microscopy and Hyperpolarized 13C-Pyruvate Magnetic Resonance Spectroscopy


 , , and
, , and
Abstract

1. Introduction
2. Methods
2.1. Cell Culture in the Bioreactor
2.2. Fluorescence Lifetime Imaging Microscopy
2.3. Hyperpolarized Pyruvate MRS
2.4. Data and Statistical Analysis
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- American Cancer Society, Breast Cancer Facts and Figures 2017–2018. Available online: www.cancer.org (accessed on 12 July 2024).
- Meltzer, A. Dormancy and breast cancer. J. Surg. Oncol. 1990, 43, 181–188. [Google Scholar] [CrossRef] [PubMed]
- Páez, D.; Labonte, M.J.; Bohanes, P.; Zhang, W.; Benhanim, L.; Ning, Y.; Wakatsuki, T.; Loupakis, F.; Lenz, H.J. Cancer dormancy: A model of early dissemination and late cancer recurrence. Clin. Cancer Res. 2012, 18, 645–653. [Google Scholar] [CrossRef]
- Brackstone, M.; Townson, J.L.; Chambers, A.F. Tumour dormancy in breast cancer: An update. Breast Cancer Res. 2007, 9, 208. [Google Scholar] [CrossRef] [PubMed]
- Cox, B.L.; Erickson-Bhatt, S.; Szulczewski, J.M.; Squirrell, J.M.; Ludwig, K.D.; Macdonald, E.B.; Swader, R.; Ponik, S.M.; Eliceiri, K.W.; Fain, S.B. A novel bioreactor for combined magnetic resonance spectroscopy and optical imaging of metabolism in 3D cell cultures. Magn. Reson. Med. 2019, 81, 3379–3391. [Google Scholar] [CrossRef]
- Warburg, O. On the origin of cancer cells. Science 1956, 123, 309–314. [Google Scholar] [CrossRef]
- Vander Heiden, M.G.; Cantley, L.C.; Thompson, C.B. Understanding the Warburg effect: The metabolic requirements of cell proliferation. Science 2009, 324, 1029–1033. [Google Scholar] [CrossRef] [PubMed]
- Dafni, H.; Ronen, S.M. Dynamic nuclear polarization in metabolic imaging of metastasis: Common sense, hypersense and compressed sensing. Cancer Biomark. 2010, 7, 189–199. [Google Scholar] [CrossRef]
- Brindle, K.M. Imaging metabolism with hyperpolarized (13)C-labeled cell substrates. J. Am. Chem. Soc. 2015, 137, 6418–6427. [Google Scholar] [CrossRef]
- Christensen, C.E.; Karlsson, M.; Winther, J.R.; Jensen, P.R.; Lerche, M.H. Non-invasive in-cell determination of free cytosolic [NAD+]/[NADH] ratios using hyperpolarized glucose show large variations in metabolic phenotypes. J. Biol. Chem. 2014, 289, 2344–2352. [Google Scholar] [CrossRef]
- Witney, T.H.; Kettunen, M.I.; Hu, D.E.; Gallagher, F.A.; Bohndiek, S.E.; Napolitano, R.; Brindle, K.M. Detecting Treatment Response in a Model of Human Breast Adenocarcinoma Using Hyperpolarised [1-13C]Pyruvate and [1,4-13C2]Fumarate. Br. J. Cancer 2010, 103, 1400–1406. [Google Scholar] [CrossRef]
- Harris, T.; Eliyahu, G.; Frydman, L.; Degani, H. Kinetics of Hyperpolarized 13C1-Pyruvate Transport and Metabolism in Living Human Breast Cancer Cells. Proc. Natl. Acad. Sci. USA 2009, 106, 18131–18136. [Google Scholar] [CrossRef] [PubMed]
- Macdonald, E.B.; Begovatz, P.; Barton, G.P.; Erickson-Bhatt, S.; Inman, D.R.; Cox, B.L.; Eliceiri, K.W.; Strigel, R.M.; Ponik, S.M.; Fain, S.B. Hyperpolarized 13C Magnetic Resonance Spectroscopic Imaging of Pyruvate Metabolism in Murine Breast Cancer Models of Different Metastatic Potential. Metabolites 2021, 11, 274. [Google Scholar] [CrossRef] [PubMed]
- Grashei, M.; Biechl, P.; Schilling, F.; Otto, A.M. Conversion of Hyperpolarized [1-13 C]Pyruvate in Breast Cancer Cells Depends on Their Malignancy, Metabolic Program and Nutrient Microenvironment. Cancers 2022, 14, 1845. [Google Scholar] [CrossRef] [PubMed]
- Sidani, M.; Wyckoff, J.; Xue, C.; Segall, J.E.; Condeelis, J. Probing the microenvironment of mammary tumors using multiphoton microscopy. J. Mammary Gland. Biol. Neoplasia 2006, 11, 151–163. [Google Scholar] [CrossRef] [PubMed]
- Provenzano, P.P.; Rueden, C.T.; Trier, S.M.; Yan, L.; Ponik, S.M.; Inman, D.R.; Keely, P.J.; Eliceiri, K.W. Nonlinear optical imaging and spectral-lifetime computational analysis of endogenous and exogenous fluorophores in breast cancer. J. Biomed. Opt. 2008, 13, 031220. [Google Scholar] [CrossRef]
- Conklin, M.W.; Provenzano, P.P.; Eliceiri, K.W.; Sullivan, R.; Keely, P.J. Fluorescence lifetime imaging of endogenous fluorophores in histopathology sections reveals differences between normal and tumor epithelium in carcinoma in situ of the breast. Cell Biochem. Biophys. 2009, 53, 145–157. [Google Scholar] [CrossRef]
- Li, L.Z.; Xu, H.N.; Ranji, M.; Nioka, S.; Chance, B. Mitochondrial redox imaging for cancer diagnostic and therapeutic studies. J. Innov. Opt. Health Sci. 2009, 2, 325–341. [Google Scholar] [CrossRef]
- Ostrander, J.H.; McMahon, C.M.; Lem, S.; Millon, S.R.; Brown, J.Q.; Seewaldt, V.L.; Ramanujam, N. Optical redox ratio differentiates breast cancer cell lines based on estrogen receptor status. Cancer Res. 2010, 70, 4759–4766. [Google Scholar] [CrossRef]
- Walsh, A.; Cook, R.S.; Rexer, B.; Arteaga, C.L.; Skala, M.C. Optical imaging of metabolism in HER2 overexpressing breast cancer cells. Biomed. Opt. Express 2012, 3, 75–85. [Google Scholar] [CrossRef]
- Xu, H.N.; Zheng, G.; Tchou, J.; Nioka, S.; Li, L.Z. Characterizing the metabolic heterogeneity in human breast cancer xenografts by 3D high resolution fluorescence imaging. Springerplus 2013, 2, 73. [Google Scholar] [CrossRef]
- Walsh, A.J.; Cook, R.S.; Sanders, M.E.; Aurisicchio, L.; Ciliberto, G.; Arteaga, C.L.; Skala, M.C. Quantitative optical imaging of primary tumor organoid metabolism predicts drug response in breast cancer. Cancer Res. 2014, 74, 5184–5194. [Google Scholar] [CrossRef] [PubMed]
- Cannon, T.M.; Shah, A.T.; Walsh, A.J.; Skala, M.C. High-throughput measurements of the optical redox ratio using a commercial microplate reader. J. Biomed. Opt. 2015, 20, 010503. [Google Scholar] [CrossRef] [PubMed]
- Sun, N.; Xu, H.N.; Luo, Q.; Li, L.Z. Potential Indexing of the Invasiveness of Breast Cancer Cells by Mitochondrial Redox Ratios. Adv. Exp. Med. Biol. 2016, 923, 121–127. [Google Scholar] [CrossRef] [PubMed]
- Alhallak, K.; Rebello, L.G.; Muldoon, T.J.; Quinn, K.P.; Rajaram, N. Optical redox ratio identifies metastatic potential-dependent changes in breast cancer cell metabolism. Biomed. Opt. Express. 2016, 7, 4364–4374. [Google Scholar] [CrossRef]
- Cannon, T.M.; Shah, A.T.; Skala, M.C. Autofluorescence imaging captures heterogeneous drug response differences between 2D and 3D breast cancer cultures. Biomed. Opt. Express 2017, 8, 1911–1925. [Google Scholar] [CrossRef]
- Hou, J.; Williams, J.; Botvinick, E.L.; Potma, E.O.; Tromberg, B.J. Visualization of Breast Cancer Metabolism Using Multimodal Nonlinear Optical Microscopy of Cellular Lipids and Redox State. Cancer Res. 2018, 78, 2503–2512. [Google Scholar] [CrossRef]
- Aslakson, C.J.; Miller, F.R. Selective events in the metastatic process defined by analysis of the sequential dissemination of subpopulations of a mouse mammary tumor. Cancer Res. 1992, 52, 1399–1405. [Google Scholar]
- Burkel, B.; Morris, B.A.; Ponik, S.M.; Riching, K.M.; Eliceiri, K.W.; Keely, P.J. Preparation of 3D Collagen Gels and Microchannels for the Study of 3D Interactions In Vivo. J. Vis. Exp. 2016, 111, e53989. [Google Scholar] [CrossRef]
- Becker, W. Fluorescence lifetime imaging--techniques and applications. J. Microsc. 2012, 247, 119–136. [Google Scholar] [CrossRef]
- Ardenkjaer-Larsen, J.H.; Fridlund, B.; Gram, A.; Hansson, G.; Hansson, L.; Lerche, M.H.; Servin, R.; Thaning, M.; Golman, K. Increase in signal-to-noise ratio of >10,000 times in liquid-state NMR. Proc. Natl. Acad. Sci. USA 2003, 100, 10158–10163. [Google Scholar] [CrossRef]
- Rowland, I.J.; Peterson, E.T.; Gordon, J.W.; Fain, S.B. Hyperpolarized 13carbon MR. Curr. Pharm. Biotechnol. 2010, 11, 709–719. [Google Scholar] [CrossRef] [PubMed]
- Naressi, A.; Couturier, C.; Devos, J.M.; Janssen, M.; Mangeat, C.; de Beer, R.; Graveron-Demilly, D. Java-based graphical user interface for the MRUI quantitation package. MAGMA 2001, 12, 141–152. [Google Scholar] [CrossRef] [PubMed]
- Morris, B.A.; Burkel, B.; Ponik, S.M.; Fan, J.; Condeelis, J.S.; Aguirre-Ghiso, J.A.; Castracane, J.; Denu, J.M.; Keely, P.J. Collagen Matrix Density Drives the Metabolic Shift in Breast Cancer Cells. EBioMedicine 2016, 13, 146–156. [Google Scholar] [CrossRef] [PubMed]
- Xing, J.; Qi, L.; Liu, X.; Shi, G.; Sun, X.; Yang, Y. Roles of mitochondrial fusion and fission in breast cancer progression: A systematic review. World J. Surg. Oncol. 2022, 20, 331. [Google Scholar] [CrossRef] [PubMed]
- Zakic, T.; Kalezic, A.; Drvendzija, Z.; Udicki, M.; Ivkovic Kapicl, T.; Srdic Galic, B.; Korac, A.; Jankovic, A.; Korac, B. Breast Cancer: Mitochondria-Centered Metabolic Alterations in Tumor and Associated Adipose Tissue. Cells 2024, 13, 155. [Google Scholar] [CrossRef]
- Sabharwal, S.S.; Schumacker, P.T. Mitochondrial ROS in cancer: Initiators, amplifiers or an Achilles’ heel? Nat. Rev. Cancer 2014, 14, 709–721. [Google Scholar] [CrossRef]

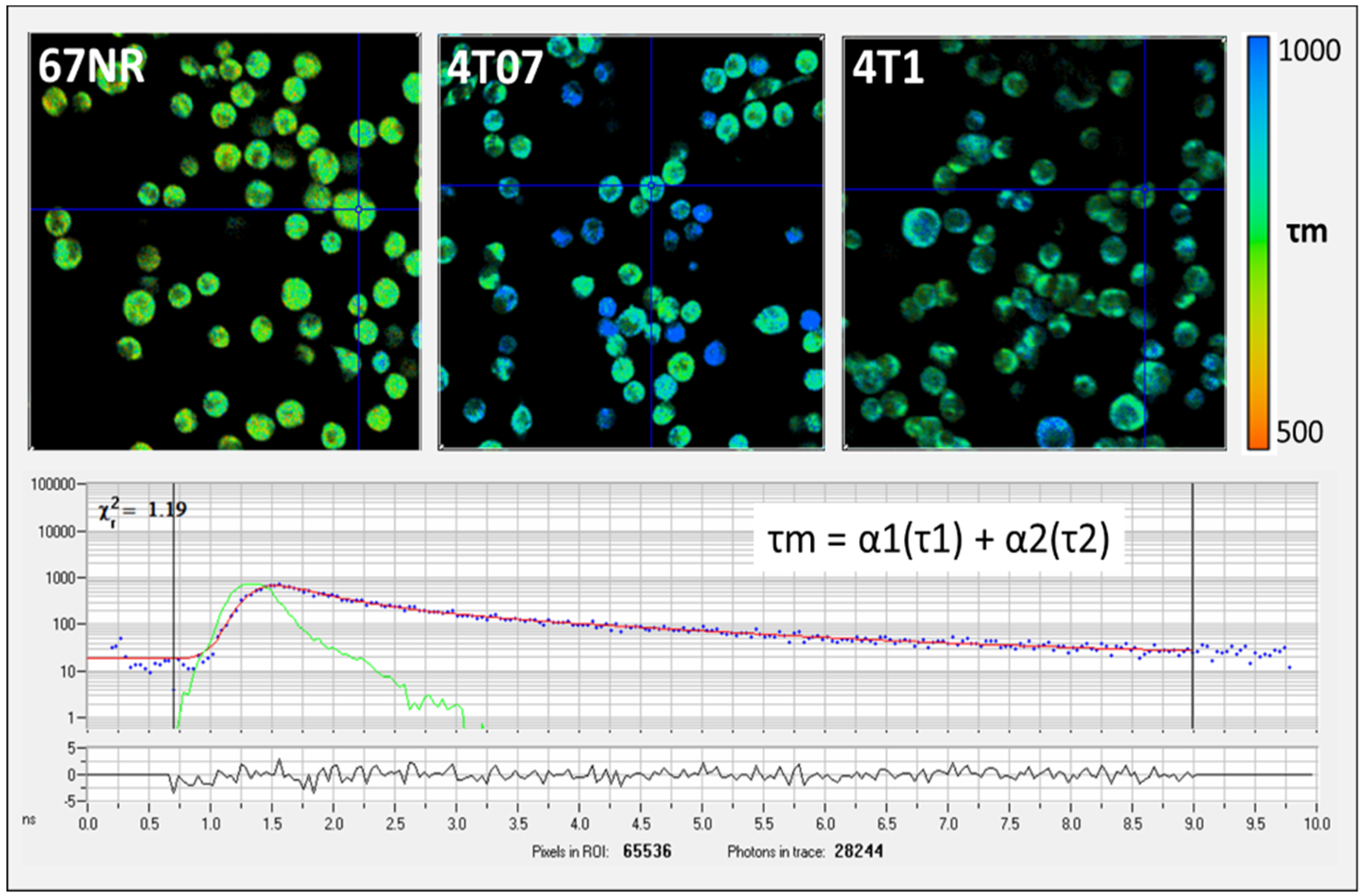
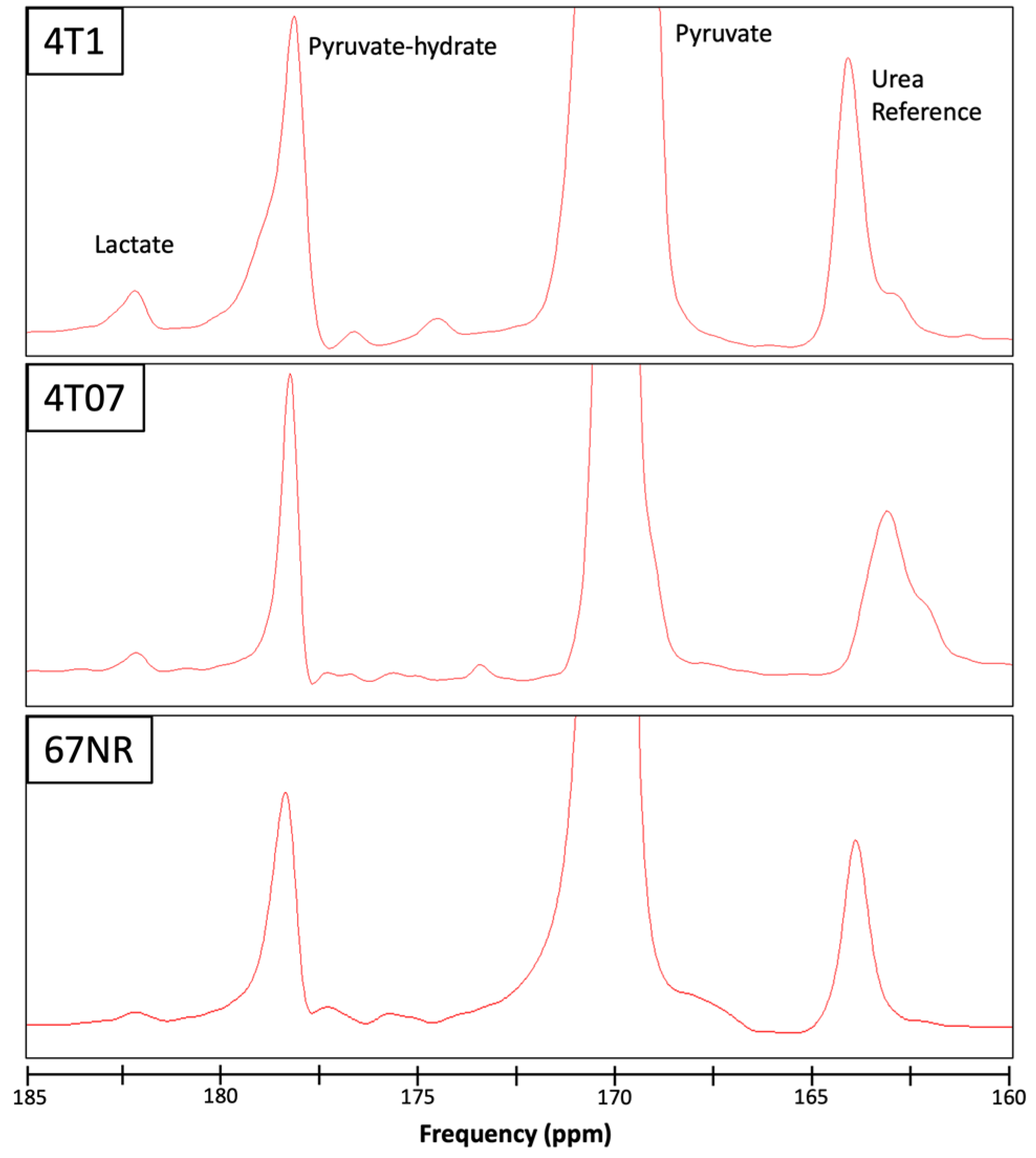
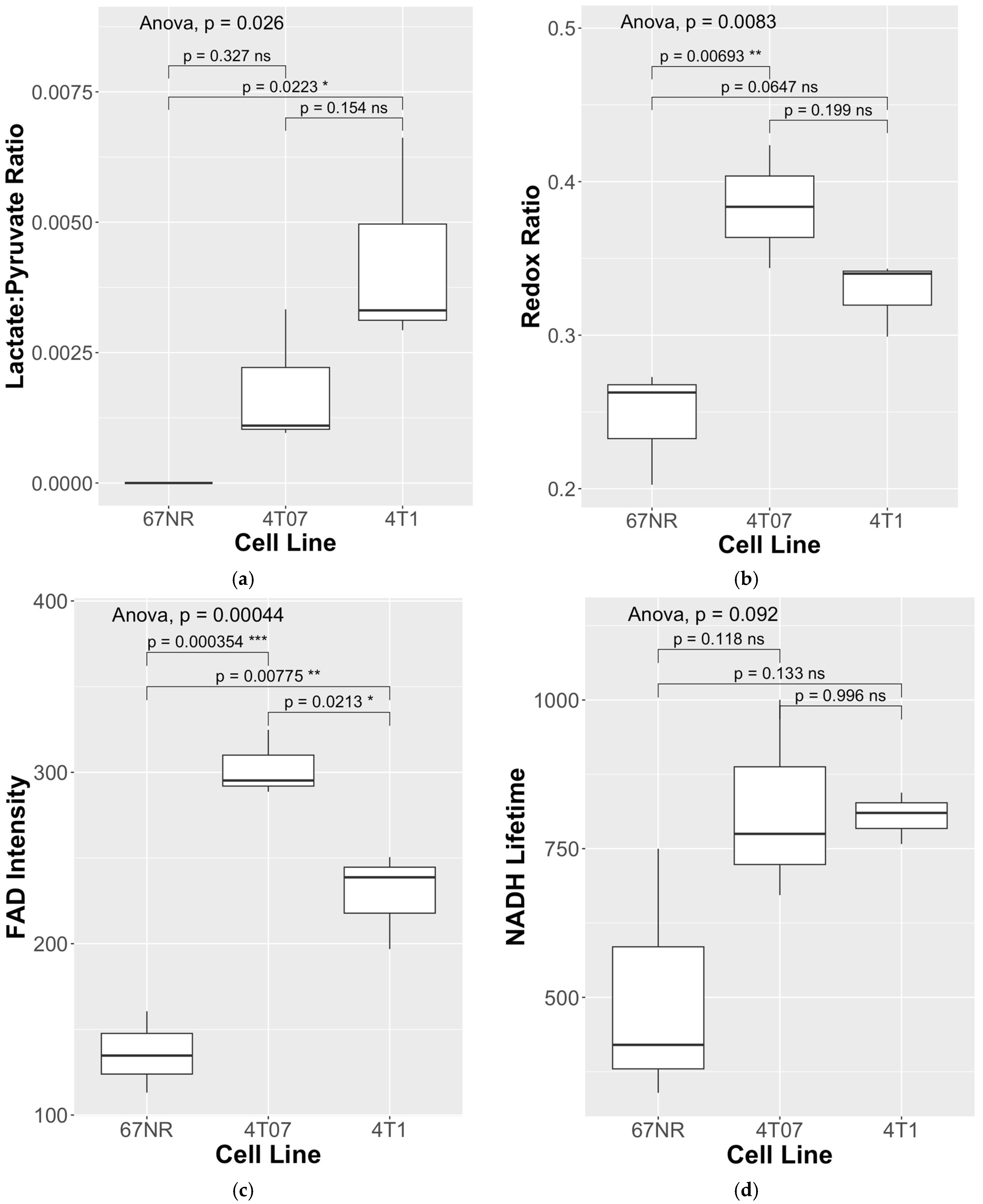

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| NADH | FAD | |||||
|---|---|---|---|---|---|---|
| Cell Line | Lac/Pyr Ratio | Redox Ratio | Intensity | Lifetime (Tau Mean, ps) | Intensity | Lifetime (Tau Mean, ps) |
| 4T1 | 0.00293 | 0.299 | 613.58 | 757.8 | 250.56 | 751.8 |
| 4T1 | 0.00331 | 0.343 | 347.40 | 810.1 | 196.88 | 835.2 |
| 4T1 | 0.00662 | 0.340 | 416.14 | 844.2 | 238.73 | 975.8 |
| Mean (SD) | 0.00429 (0.0020) | 0.327 (0.02) | 459.04 (138.2) | 804.0 (43.5) | 228.7 (28.2) | 854.3 (113.2) |
| 4T07 | 0.00333 | 0.424 | 432.05 | 1000.3 | 324.82 | 964.1 |
| 4T07 | 0.0011 | 0.344 | 520.70 | 671.9 | 288.71 | 910.8 |
| 4T07 | 0.00096 | 0.384 | 438.34 | 774.9 | 295.24 | 971.7 |
| Mean (SD) | 0.00180 (0.0013) | 0.384 (0.04) | 463.70 (49.5) | 815.7 (168.0) | 302.9 (19.2) | 948.9 (33.2) |
| 67NR | 0 | 0.273 | 464.36 | 749.8 | 134.70 | 813.2 |
| 67NR | 0 | 0.263 | 414.45 | 339.5 | 160.55 | 396.9 |
| 67NR | 0 | 0.203 | 448.20 | 420.1 | 113.11 | 935.1 |
| Mean (SD) | 0 (0) | 0.246 (0.04) | 442.33 (25.5) | 503.1 (217.4) | 136.1 (23.8) | 715.1 (282.2) |
| Comparison HP-MRS Lac/Pyr Ratio vs. | Spearman Correlation Coefficient | p-Value |
|---|---|---|
| Redox Ratio | 0.63 | 0.071 + |
| NADH Intensity | −0.25 | 0.51 |
| NADH Lifetime | 0.86 | 0.0026 * |
| FAD Intensity | 0.59 | 0.092 |
| FAD Lifetime | 0.49 | 0.18 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Erickson-Bhatt, S.; Cox, B.L.; Macdonald, E.; Chacko, J.V.; Begovatz, P.; Keely, P.J.; Ponik, S.M.; Eliceiri, K.W.; Fain, S.B. Multi-Modal Investigation of Metabolism in Murine Breast Cancer Cell Lines Using Fluorescence Lifetime Microscopy and Hyperpolarized 13C-Pyruvate Magnetic Resonance Spectroscopy. Metabolites 2024, 14, 550. https://doi.org/10.3390/metabo14100550
Erickson-Bhatt S, Cox BL, Macdonald E, Chacko JV, Begovatz P, Keely PJ, Ponik SM, Eliceiri KW, Fain SB. Multi-Modal Investigation of Metabolism in Murine Breast Cancer Cell Lines Using Fluorescence Lifetime Microscopy and Hyperpolarized 13C-Pyruvate Magnetic Resonance Spectroscopy. Metabolites. 2024; 14(10):550. https://doi.org/10.3390/metabo14100550
Chicago/Turabian StyleErickson-Bhatt, Sarah, Benjamin L. Cox, Erin Macdonald, Jenu V. Chacko, Paul Begovatz, Patricia J. Keely, Suzanne M. Ponik, Kevin W. Eliceiri, and Sean B. Fain. 2024. "Multi-Modal Investigation of Metabolism in Murine Breast Cancer Cell Lines Using Fluorescence Lifetime Microscopy and Hyperpolarized 13C-Pyruvate Magnetic Resonance Spectroscopy" Metabolites 14, no. 10: 550. https://doi.org/10.3390/metabo14100550
APA StyleErickson-Bhatt, S., Cox, B. L., Macdonald, E., Chacko, J. V., Begovatz, P., Keely, P. J., Ponik, S. M., Eliceiri, K. W., & Fain, S. B. (2024). Multi-Modal Investigation of Metabolism in Murine Breast Cancer Cell Lines Using Fluorescence Lifetime Microscopy and Hyperpolarized 13C-Pyruvate Magnetic Resonance Spectroscopy. Metabolites, 14(10), 550. https://doi.org/10.3390/metabo14100550