
Ultra-High-Performance Liquid Chromatography–High-Definition Mass Spectrometry-Based Metabolomics to Reveal the Potential Anti-Arthritic Effects of Illicium verum in Cultured Fibroblast-like Synoviocytes Derived from Rheumatoid Arthritis

 , ,
, ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Chemicals and Reagents
2.2. Herbal Extraction Preparation
2.3. Cell Culture
2.4. Cell Proliferation Assay
2.5. Cell Migration Assay
2.6. Cell Inflammatory Factors Analysis
2.7. Cell Metabolomic Analysis
2.7.1. Sample Preparation
2.7.2. UPLC-MS Analysis
2.7.3. Data Processing
2.8. Statistical Analysis
3. Results
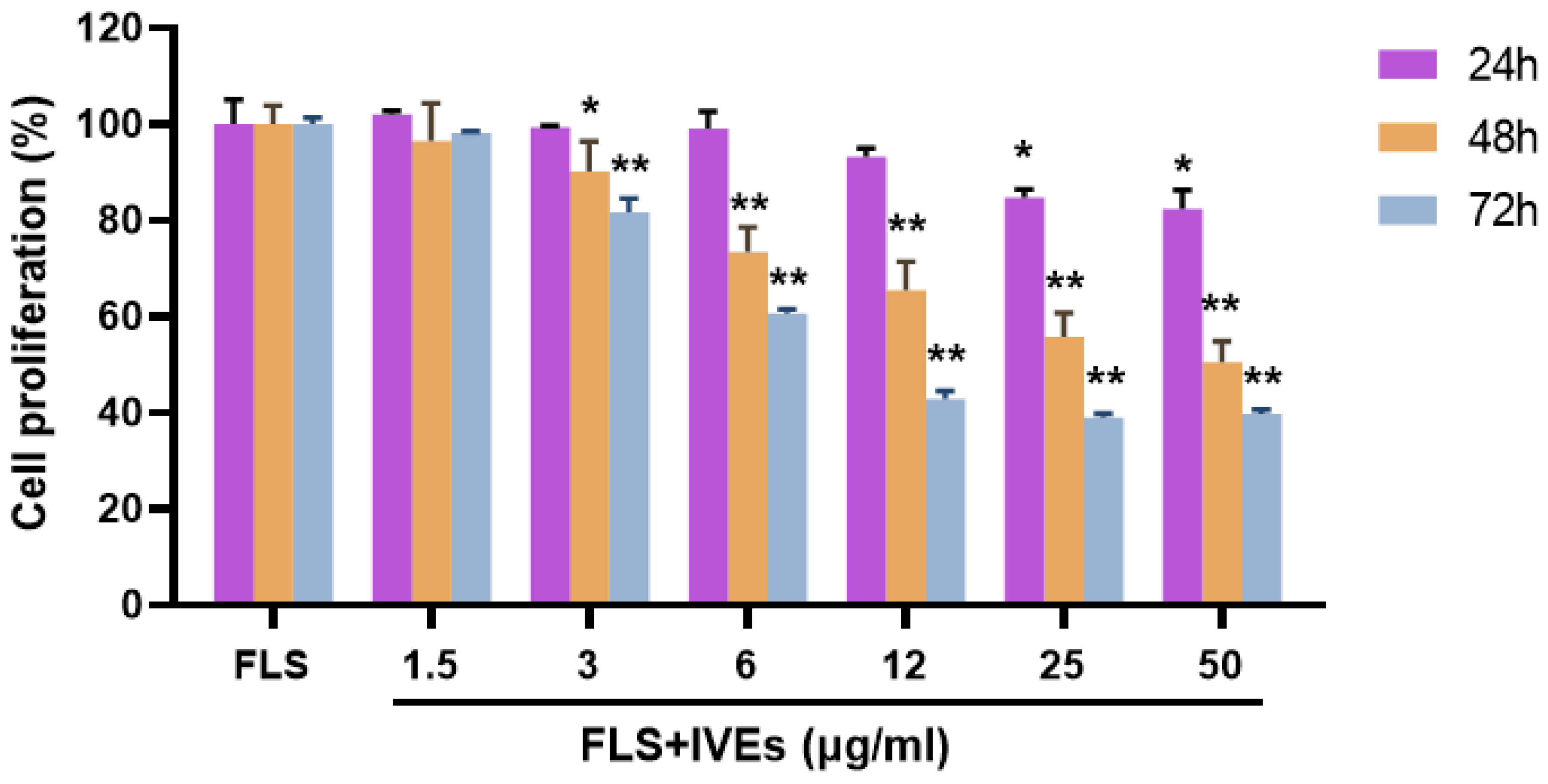
3.1. IVEs Inhibited the Cell Proliferation of RA-FLS
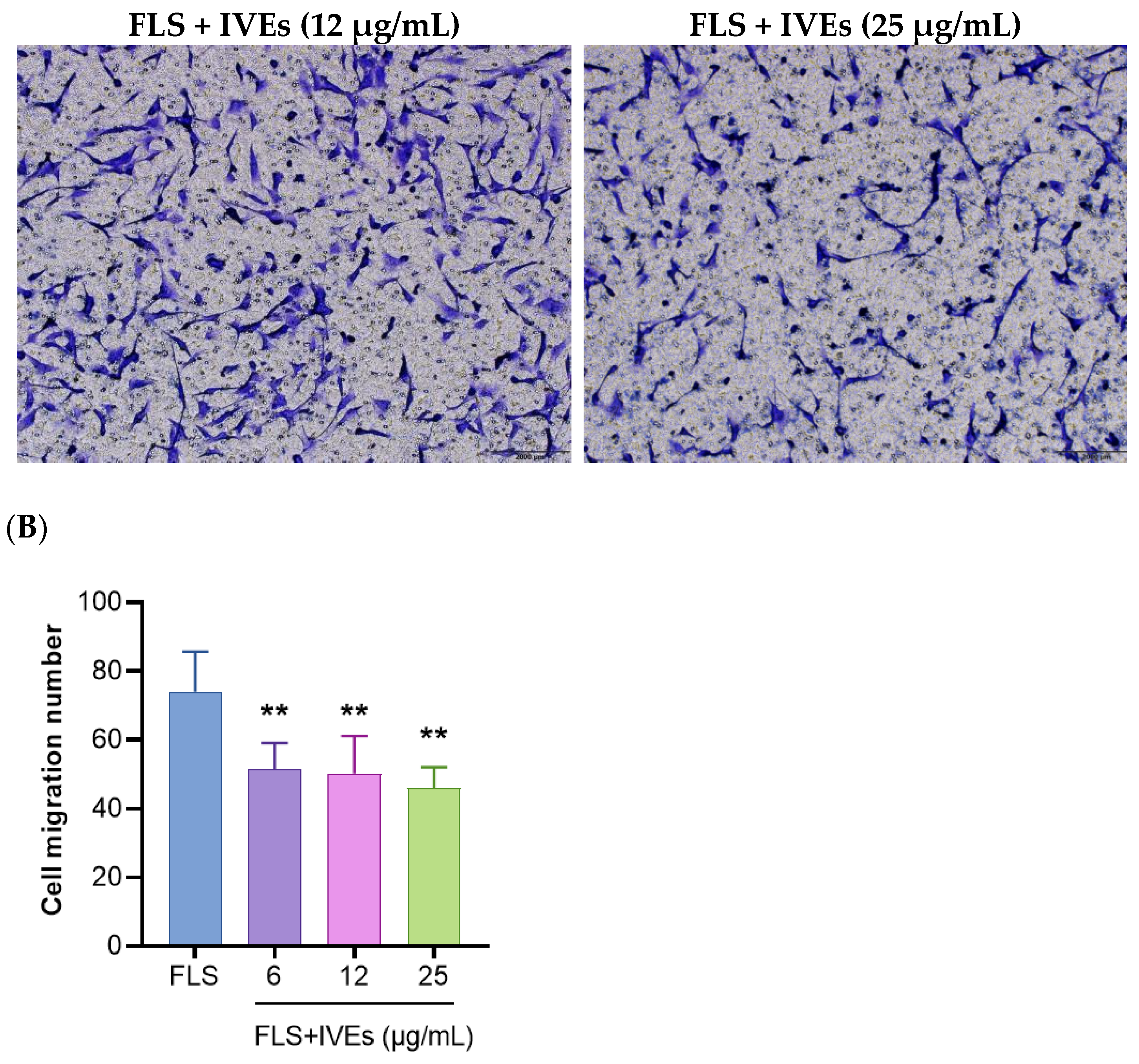
3.2. IVEs Inhibited the Cell Migration of RA-FLS
3.3. IVEs Inhibited the Production of Inflammatory Factors from RA-FLS
3.4. IVEs Regulated the Cell Metabolomic Profiles of RA-FLS
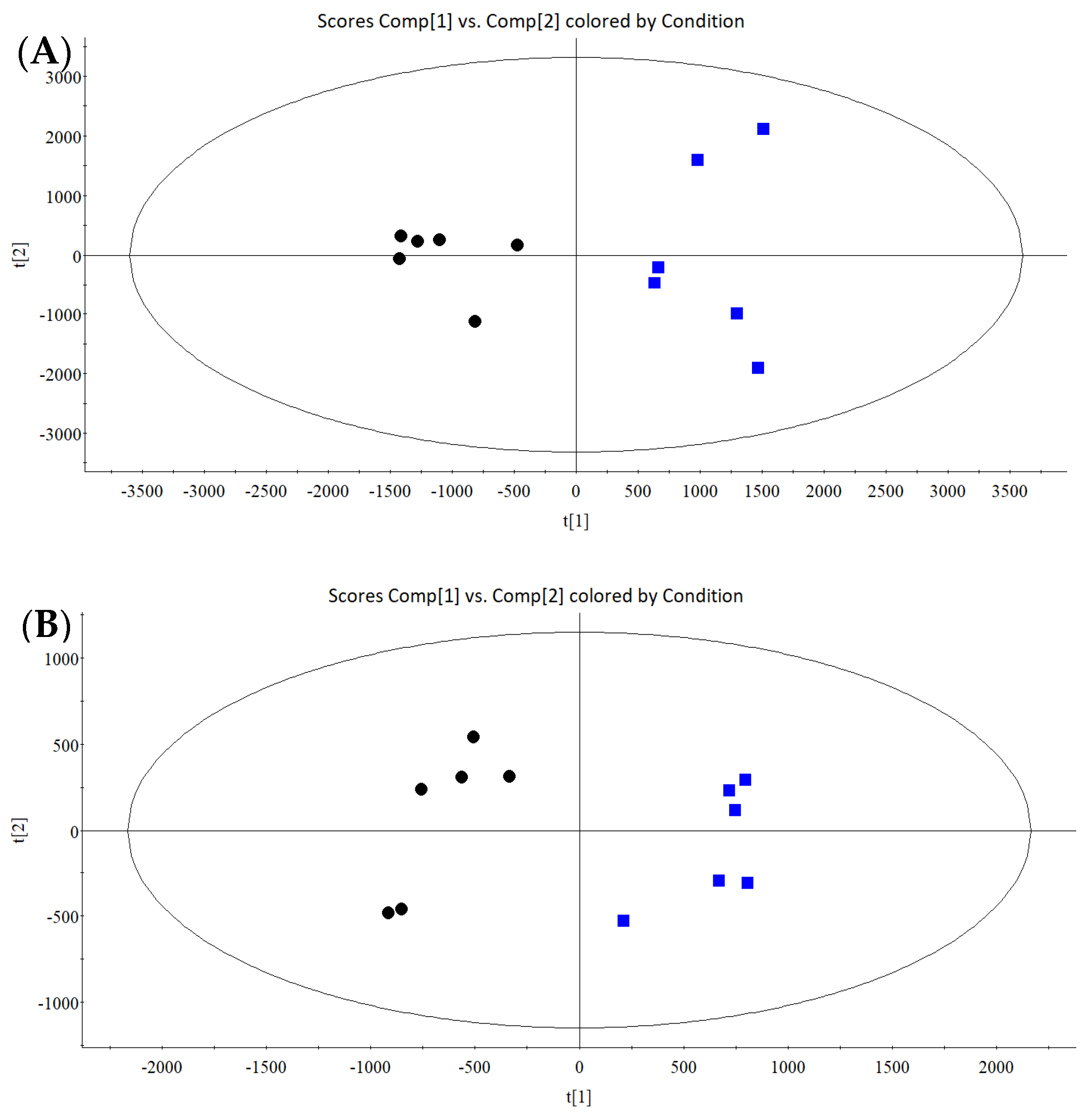
3.4.1. Multivariate Statistical Analysis
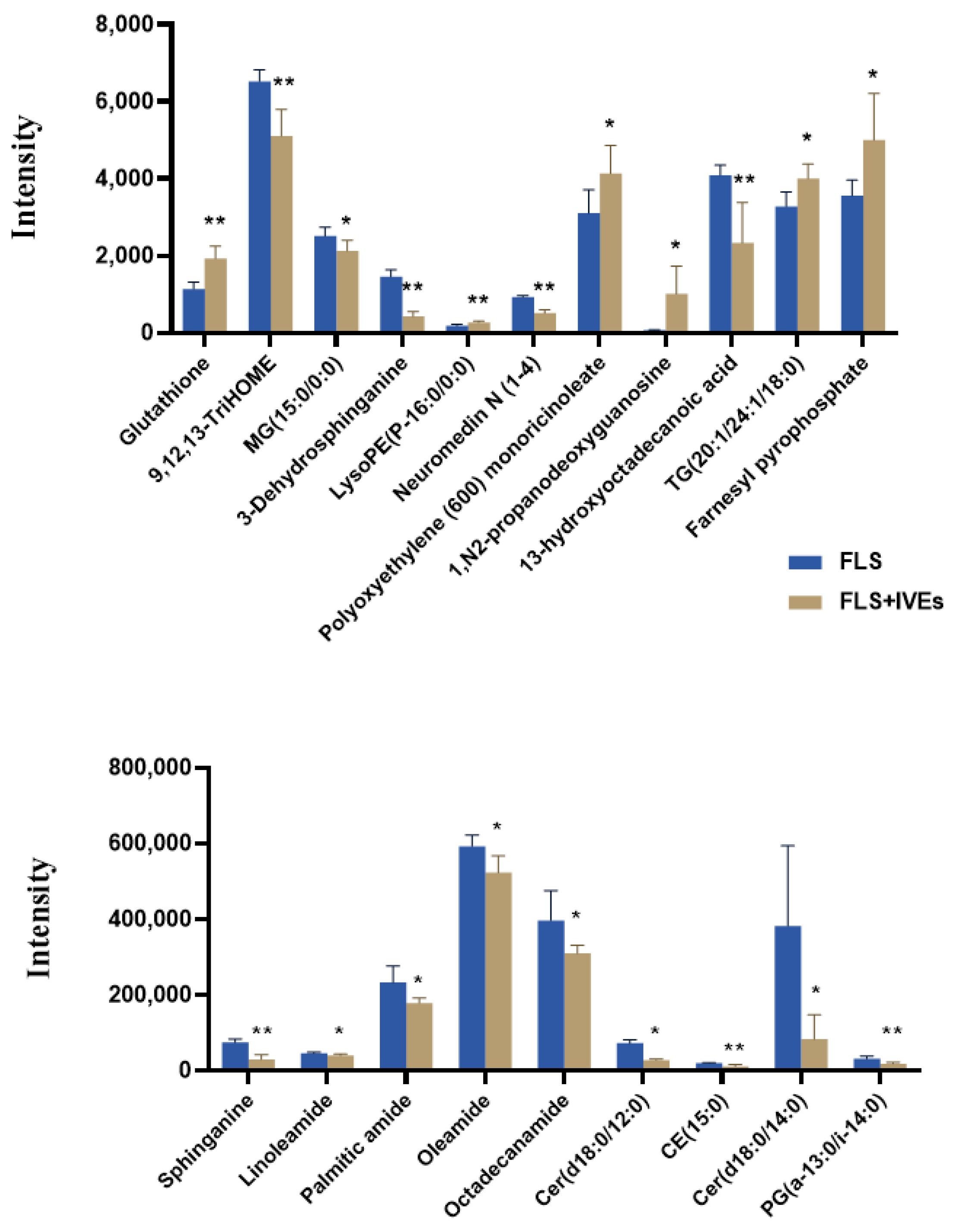
3.4.2. Differential Metabolite Identification
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Smolen, J.S.; Aletaha, D.; McInnes, I. Rheumatoid arthritis. Lancet 2016, 388, 2023–2038. [Google Scholar] [CrossRef] [PubMed]
- Robinson, G.; Pineda-Torra, I.; Ciurtin, C.; Jury, E.C. Lipid metabolism in autoimmune rheumatic disease: Implications for modern and conventional therapies. J. Clin. Investig. 2022, 132, e148552. [Google Scholar] [CrossRef] [PubMed]
- McInnes, I.B.; Schett, G. Pathogenetic insights from the treatment of rheumatoid arthritis. Lancet 2017, 389, 2328–2337. [Google Scholar] [CrossRef] [PubMed]
- Köhler, B.M.; Günther, J.; Kaudewitz, D.; Lorenz, H.M. Current Therapeutic Options in the Treatment of Rheumatoid Arthritis. J. Clin. Med. 2019, 8, 938. [Google Scholar] [CrossRef] [PubMed]
- Burmester, G.R.; Pope, J.E. Novel treatment strategies in rheumatoid arthritis. Lancet 2017, 389, 2338–2348. [Google Scholar] [CrossRef]
- Handa, O.; Naito, Y.; Fukui, A.; Omatsu, T.; Yoshikawa, T. The impact of non-steroidal anti-inflammatory drugs on the small intestinal epithelium. J. Clin. Biochem. Nutr. 2014, 54, 2–6. [Google Scholar] [CrossRef]
- Solomon, D.H.; Glynn, R.J.; Karlson, E.W.; Lu, F.; Ridker, P.M. Adverse Effects of Low-Dose Methotrexate: A Randomized Trial. Ann. Intern. Med. 2020, 172, 369–380. [Google Scholar] [CrossRef]
- Vanni, K.M.M.; Berliner, N.; Paynter, N.P.; Glynn, R.J.; MacFadyen, J.; Colls, J.; Lu, F.; Xu, C.; Ridker, P.M.; Solomon, D.H. Adverse Effects of Low-Dose Methotrexate in a Randomized Double-Blind Placebo-Controlled Trial: Adjudicated Hematologic and Skin Cancer Outcomes in the Cardiovascular Inflammation Reduction Trial. ACR Open Rheumatol. 2020, 2, 697–704. [Google Scholar] [CrossRef]
- Shahrajabian, M.H.; Sun, W.; Cheng, Q. Survey on Chemical Constituent, Traditional and Modern Pharmaceutical and Health Benefits of Chinese Star Anise, a Treasure from the East. Pharmacogn. Commun. 2021, 11, 31–35. [Google Scholar] [CrossRef]
- Kerna, N.A.; Anderson, I.I.J.; Pruitt, K.D.; Flores, J.V.; Ortigas, M.A.C.; Carsrud, N.D.V.; Holets, H.M.; Nomel, R.; Nwokorie, U.; Nwachukwu, D. Star Anise: A Purported Antiviral Herb with Numerous Associated Health Benefits. EC Clin. Med. Case Rep. 2021, 4, 63–73. [Google Scholar]
- Tuseef, H.; Raza, M.L.; Assad, T. Comparative Evaluation of Analgesic, Antipyretic & Anti-Inflammatory Effects of Various Extracts of Dried Fruit of Illicium verum Hook.f (Star anise) in Rodents. Walailak J. Sci. Technol. 2021, 18, 9456. [Google Scholar]
- Shahrajabian, M.H.; Sun, W.; Cheng, Q. Chinese star anise (Illicium verum) and pyrethrum (Chrysanthemum cinerariifolium) as natural alternatives for organic farming and health care—A review. Aust. J. Crop Sci. 2020, 14, 517–523. [Google Scholar] [CrossRef]
- Ritter, A.M.V.; Hernandes, L.; da Rocha, B.A.; Estevão-Silva, C.F.; Wisniewski-Rebecca, E.S.; Cezar, J.D.S.; Caparroz-Assef, S.M.; Cuman, R.K.N.; Bersani-Amado, C.A. Anethole reduces inflammation and joint damage in rats with adjuvant-induced arthritis. Inflamm. Res. 2017, 66, 725–737. [Google Scholar] [CrossRef]
- Németh, T.; Nagy, G.; Pap, T. Synovial fibroblasts as potential drug targets in rheumatoid arthritis, where do we stand and where shall we go? Ann. Rheum. Dis. 2022, 81, 1055–1064. [Google Scholar] [CrossRef] [PubMed]
- Bottini, N.; Firestein, G.S. Duality of fibroblast-like synoviocytes in RA: Passive responders and imprinted aggressors. Nat. Rev. Rheumatol. 2013, 9, 24–33. [Google Scholar] [CrossRef]
- Ahn, J.K.; Kim, S.; Hwang, J.; Kim, J.; Kim, K.H.; Cha, H.S. GC/TOF-MS-based metabolomic profiling in cultured fibroblast-like synoviocytes from rheumatoid arthritis. Jt. Bone Spine 2016, 83, 707–713. [Google Scholar] [CrossRef]
- Zhang, A.H.; Sun, H.; Wang, X.J. Potential applications and development of cell metabolomics in natural products. J. Drug Metab. Toxicol. 2014, 5, 2. [Google Scholar]
- Hounoum, B.M.; Blasco, H.; Emond, P.; Mavel, S. Liquid chromatography–high-resolution mass spectrometry-based cell metabolomics: Experimental design, recommendations, and applications. Trends Anal. Chem. 2016, 75, 118–128. [Google Scholar] [CrossRef]
- McGrath, C.M.; Young, S.P. Lipid and Metabolic Changes in Rheumatoid Arthritis. Curr. Rheumatol. Rep. 2015, 17, 57. [Google Scholar] [CrossRef]
- Bustamante, M.F.; Garcia-Carbonell, R.; Whisenant, K.D.; Guma, M. Fibroblast-like synoviocyte metabolism in the pathogenesis of rheumatoid arthritis. Arthritis Res. Ther. 2017, 19, 110. [Google Scholar] [CrossRef]
- Falconer, J.; Murphy, A.N.; Young, S.P.; Clark, A.R.; Tiziani, S.; Guma, M.; Buckley, C.D. Review: Synovial Cell Metabolism and Chronic Inflammation in Rheumatoid Arthritis. Arthritis Rheumatol. 2018, 70, 984–999. [Google Scholar] [CrossRef] [PubMed]
- Han, X. Lipidomics for studying metabolism. Nat. Rev. Endocrinol. 2016, 12, 668–679. [Google Scholar] [CrossRef]
- Kloska, A.; Węsierska, M.; Malinowska, M.; Gabig-Cimińska, M.; Jakóbkiewicz-Banecka, J. Lipophagy and Lipolysis Status in Lipid Storage and Lipid Metabolism Diseases. Int. J. Mol. Sci. 2020, 21, 6113. [Google Scholar] [CrossRef] [PubMed]
- Hwang, S.; Chung, K.W. Targeting fatty acid metabolism for fibrotic disorders. Arch. Pharm. Res. 2021, 44, 839–856. [Google Scholar] [CrossRef] [PubMed]
- Veselinovic, M.; Barudzic, N.; Vuletic, M.; Zivkovic, V.; Tomic-Lucic, A.; Djuric, D.; Jakovljevic, V. Oxidative stress in rheumatoid arthritis patients: Relationship to diseases activity. Mol. Cell. Biochem. 2014, 391, 225–232. [Google Scholar] [CrossRef]
- Seal, A.; Hughes, M.; Wei, F.; Pugazhendhi, A.S.; Ngo, C.; Ruiz, J.; Schwartzman, J.D.; Coathup, M.J. Sphingolipid-Induced Bone Regulation and Its Emerging Role in Dysfunction Due to Disease and Infection. Int. J. Mol. Sci. 2024, 25, 3024. [Google Scholar] [CrossRef]
- Gomez-Larrauri, A.; Presa, N.; Dominguez-Herrera, A.; Ouro, A.; Gomez-Muoz, A. Role of bioactive sphingolipids in physiology and pathology. Essays Biochem. 2020, 64, 579–589. [Google Scholar]
- Ohanian, J.; Ohanian, V. Sphingolipids in mammalian cell signalling. Cell. Mol. Life Sci. 2001, 58, 2053–2068. [Google Scholar] [CrossRef]
- Hannun, Y.A.; Obeid, L.M. Sphingolipids and their metabolism in physiology and disease. Nat. Rev. Mol. Cell Biol. 2018, 19, 175–191. [Google Scholar] [CrossRef]
- Chaurasia, B.; Summers, S.A. Ceramides in Metabolism: Key Lipotoxic Players. Annu. Rev. Physiol. 2021, 83, 303–330. [Google Scholar] [CrossRef]
- Summers, S.A.; Chaurasia, B.; Holland, W.L. Metabolic Messengers: Ceramides. Nat. Metab. 2019, 1, 1051–1058. [Google Scholar] [CrossRef] [PubMed]
- Maceyka, M.; Spiegel, S. Sphingolipid metabolites in inflammatory disease. Nature 2014, 510, 58–67. [Google Scholar] [CrossRef] [PubMed]
- Downton, P.; Sanna, F.; Maidstone, R.; Poolman, T.M.; Hayter, E.A.; Dickson, S.H.; Ciccone, N.A.; Early, J.O.; Adamson, A.; Spiller, D.G.; et al. Chronic inflammatory arthritis drives systemic changes in circadian energy metabolism. Proc. Natl. Acad. Sci. USA 2022, 119, e2112781119. [Google Scholar] [CrossRef] [PubMed]
- Kosinska, M.K.; Liebisch, G.; Lochnit, G.; Wilhelm, J.; Klein, H.; Kaesser, U.; Lasczkowski, G.; Rickert, M.; Schmitz, G.; Steinmeyer, J. Sphingolipids in human synovial fluid—A lipidomic study. PLoS ONE 2014, 9, e91769. [Google Scholar] [CrossRef] [PubMed]
- Miltenberger-Miltenyi, G.; Cruz-Machado, A.R.; Saville, J.; Conceição, V.A.; Calado, Â.; Lopes, I.; Fuller, M.; Fonseca, J.E. Increased monohexosylceramide levels in the serum of established rheumatoid arthritis patients. Rheumatology 2020, 59, 2085–2089. [Google Scholar] [CrossRef]
- Dekkers, K.F.; Slieker, R.C.; Ioan-Facsinay, A.; van Iterson, M.; BIOS Consortium; Ikram, M.A.; van Greevenbroek, M.M.J.; Veldink, J.H.; Franke, L.; Boomsma, D.I.; et al. Lipid-induced transcriptomic changes in blood link to lipid metabolism and allergic response. Nat. Commun. 2023, 14, 544. [Google Scholar] [CrossRef]
- Abdel-Khalek, M.A.; El-Barbary, A.M.; Essa, S.A.; Ghobashi, A.S. Serum hepcidin: A direct link between anemia of inflammation and coronary artery atherosclerosis in patients with rheumatoid arthritis. J. Rheumatol. 2011, 38, 2153–2159. [Google Scholar] [CrossRef]
- Li, R.; Guo, L.X.; Li, Y.; Chang, W.Q.; Liu, J.Q.; Liu, L.F.; Xin, G.Z. Dose-response characteristics of Clematis triterpenoid saponins and clematichinenoside AR in rheumatoid arthritis rats by liquid chromatography/mass spectrometry-based serum and urine metabolomics. J. Pharm. Biomed. Anal. 2017, 136, 81–91. [Google Scholar] [CrossRef]
- Perricone, C.; De Carolis, C.; Perricone, R. Glutathione: A key player in autoimmunity. Autoimmun. Rev. 2009, 8, 697–701. [Google Scholar] [CrossRef]
- Hayes, J.D.; McLellan, L.I. Glutathione and glutathione-dependent enzymes represent a co-ordinately regulated defence against oxidative stress. Free Radic. Res. 1999, 31, 273–300. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No | Metabolite | Formula | Retention Time (min) | m/z | Adducts | Trends | Category |
|---|---|---|---|---|---|---|---|
| 1 | Glutathione | C10H17N3O6S | 0.91 | 308.0917 | M + H | ↑** | Amino acids and peptides |
| 2 | 9,12,13-TriHOME | C18H34O5 | 2.65 | 329.2332 | M − H | ↓** | Fatty Acyls |
| 3 | MG (15:0/0:0) | C18H36O4 | 3.93 | 315.2535 | M − H | ↓* | Glycerophospholipids |
| 4 | Sphinganine | C18H39NO2 | 4.00 | 302.3059 | M + H | ↓** | Sphingolipids |
| 5 | 3-Dehydrosphinganine | C18H37NO2 | 4.27 | 300.2903 | M + H | ↓** | Sphingolipids |
| 6 | LysoPE (P-16:0/0:0) | C21H44NO6P | 4.85 | 436.2807 | M − H | ↑** | Glycerophospholipids |
| 7 | Linoleamide | C18H33NO | 5.89 | 280.2646 | M + H | ↓* | Fatty acyls |
| 8 | Palmitic amide | C16H33NO | 6.64 | 256.2648 | M + H | ↓* | Fatty acyls |
| 9 | Oleamide | C18H35NO | 6.89 | 282.2804 | M + H | ↓* | Fatty acyls |
| 10 | Octadecanamide | C18H37NO | 8.10 | 284.296 | M + H | ↓* | Amino acids, peptides, and analogs |
| 11 | Neuromedin N (1-4) | C26H40N4O6 | 8.14 | 1007.5872 | 2M − H | ↓** | Fatty acyls |
| 12 | Polyoxyethylene (600) monoricinoleic acid | C21H40O3 | 8.58 | 341.3032 | M + H | ↑* | Fatty acyls |
| 13 | 1,N2-propanodeoxyguanosine | C13H17N5O4 | 9.31 | 330.1163 | M + Na | ↑* | Nucleoside and nucleotide analogs |
| 14 | Cer (d18:0/12:0) | C30H61NO3 | 9.56 | 484.4722 | M + H | ↓* | Sphingolipids |
| 15 | CE (15:0) | C42H74O2 | 10.50 | 609.5558 | M − H | ↓** | Sterol lipids |
| 16 | 13-hydroxyoctadecanoic acid | C18H36O3 | 10.53 | 599.5258 | 2M − H | ↓** | Fatty acyls |
| 17 | Cer (d18:0/14:0) | C32H65NO3 | 10.62 | 512.504 | M + H | ↓* | Sphingolipids |
| 18 | PG (a-13:0/i-14:0) | C33H65O10P | 11.07 | 653.4431 | M + H | ↓** | Glycerophospholipids |
| 19 | TG (20:1/24:1/18:0) | C65H124O5 | 11.54 | 1007.9286 | M + Na | ↑* | Glycerolipids |
| 20 | Farnesyl pyrophosphate | C15H28O7P2 | 11.86 | 383.1421 | M + H | ↑* | Terpenoid backbone biosynthesis |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Qin, M.; Chen, L.; Hou, X.; Wu, W.; Liu, Y.; Pan, Y.; Zhang, M.; Tan, Z.; Huang, D. Ultra-High-Performance Liquid Chromatography–High-Definition Mass Spectrometry-Based Metabolomics to Reveal the Potential Anti-Arthritic Effects of Illicium verum in Cultured Fibroblast-like Synoviocytes Derived from Rheumatoid Arthritis. Metabolites 2024, 14, 517. https://doi.org/10.3390/metabo14100517
Qin M, Chen L, Hou X, Wu W, Liu Y, Pan Y, Zhang M, Tan Z, Huang D. Ultra-High-Performance Liquid Chromatography–High-Definition Mass Spectrometry-Based Metabolomics to Reveal the Potential Anti-Arthritic Effects of Illicium verum in Cultured Fibroblast-like Synoviocytes Derived from Rheumatoid Arthritis. Metabolites. 2024; 14(10):517. https://doi.org/10.3390/metabo14100517
Chicago/Turabian StyleQin, Mingzhen, Lu Chen, Xiaoli Hou, Wuwei Wu, Yu Liu, Yu Pan, Mengli Zhang, Zhien Tan, and Danna Huang. 2024. "Ultra-High-Performance Liquid Chromatography–High-Definition Mass Spectrometry-Based Metabolomics to Reveal the Potential Anti-Arthritic Effects of Illicium verum in Cultured Fibroblast-like Synoviocytes Derived from Rheumatoid Arthritis" Metabolites 14, no. 10: 517. https://doi.org/10.3390/metabo14100517
APA StyleQin, M., Chen, L., Hou, X., Wu, W., Liu, Y., Pan, Y., Zhang, M., Tan, Z., & Huang, D. (2024). Ultra-High-Performance Liquid Chromatography–High-Definition Mass Spectrometry-Based Metabolomics to Reveal the Potential Anti-Arthritic Effects of Illicium verum in Cultured Fibroblast-like Synoviocytes Derived from Rheumatoid Arthritis. Metabolites, 14(10), 517. https://doi.org/10.3390/metabo14100517