



Impact of Heavy Metal Exposure on Mytilus galloprovincialis Spermatozoa: A Metabolomic Investigation

 ,
,  , ,
, ,  , ,
, ,  and
and 
Abstract
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Ethical Statement
2.2. Exposure of Mussels
2.3. Processing and Sampling of Spermatozoa
2.3.1. Metabolite Extraction, Purification and Derivatization
2.3.2. GC-MS Analysis
2.3.3. Metabolite Identification
2.4. Statistical Analysis
2.4.1. Animals’ Characteristics
2.4.2. Metabolomic Data Analysis
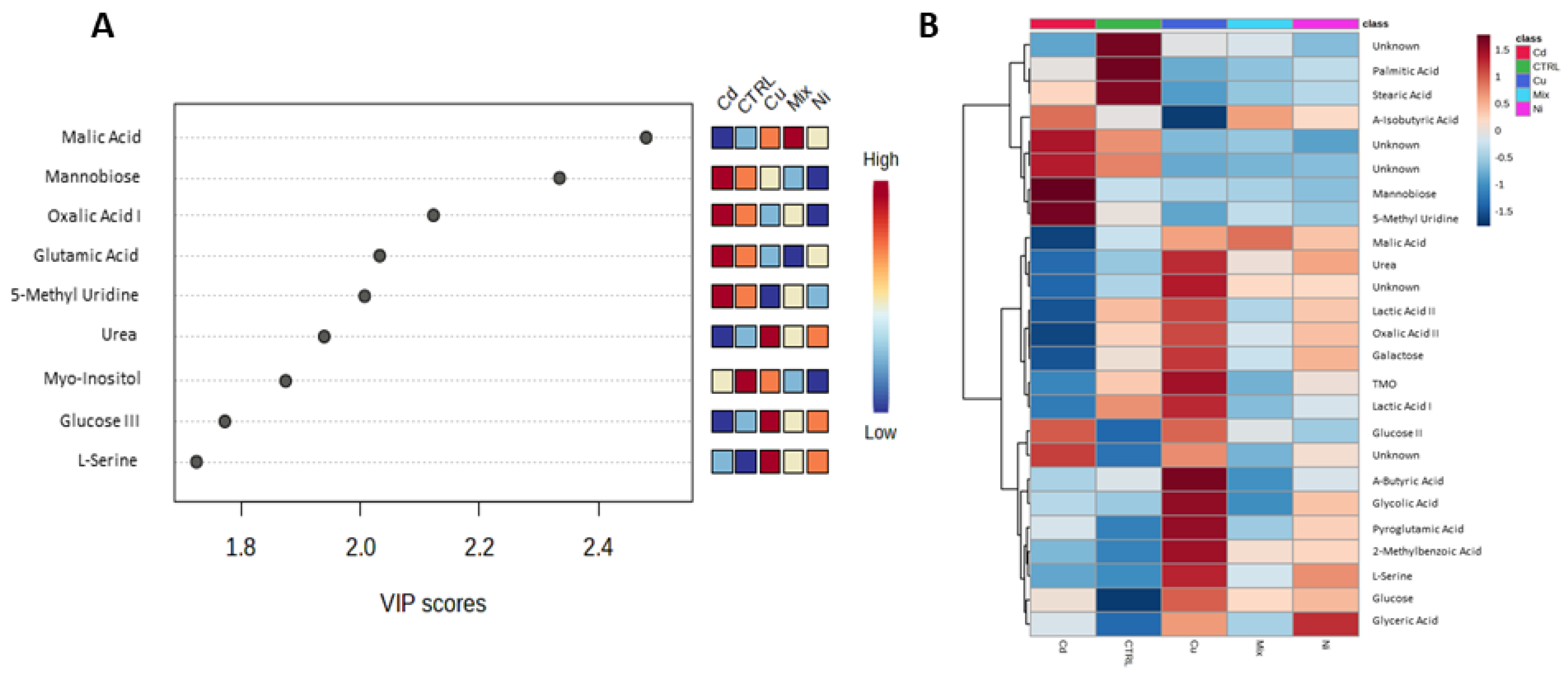
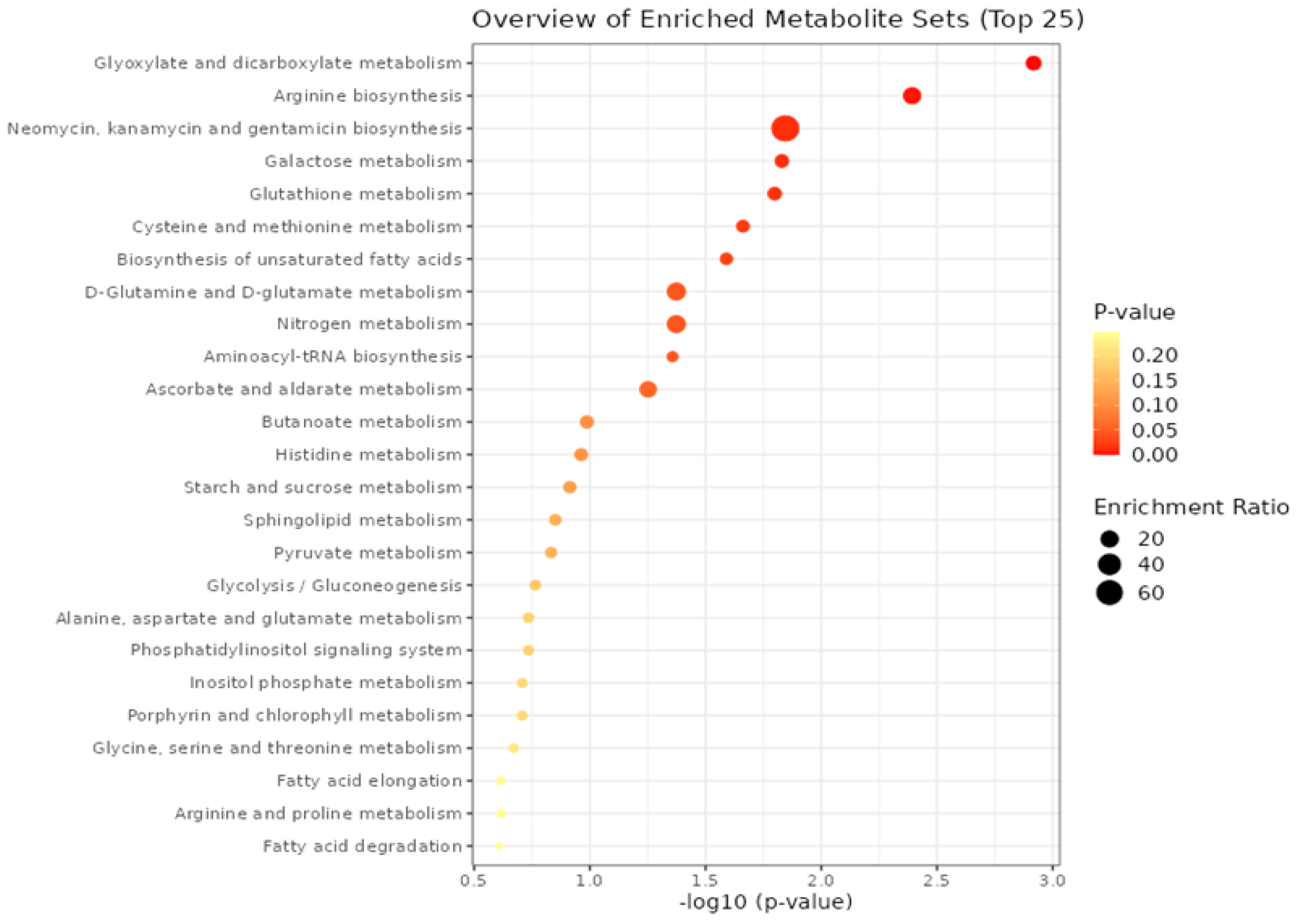
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Cossu, C.; Doyotte, A.; Babut, M.; Exinger, A.; Vasseur, P. Antioxidant Biomarkers in Freshwater Bivalves, Unio Tumidus, in Response to Different Contamination Profiles of Aquatic Sediments. Ecotoxicol. Environ. Saf. 2000, 45, 106–121. [Google Scholar] [CrossRef] [PubMed]
- Regoli, F.; Pellegrini, D.; Winston, G.W.; Gorbi, S.; Giuliani, S.; Virno-Lamberti, C.; Bompadre, S. Application of Biomarkers for Assessing the Biological Impact of Dredged Materials in the Mediterranean: The Relationship between Antioxidant Responses and Susceptibility to Oxidative Stress in the Red Mullet (Mullus Barbatus). Mar. Pollut. Bull. 2002, 44, 912–922. [Google Scholar] [CrossRef]
- De Luca-Abbott, S.B.; Richardson, B.J.; McClellan, K.E.; Zheng, G.J.; Martin, M.; Lam, P.K.S. Field Validation of Antioxidant Enzyme Biomarkers in Mussels (Perna viridis) and Clams (Ruditapes philippinarum) Transplanted in Hong Kong Coastal Waters. Mar. Pollut. Bull. 2005, 51, 694–707. [Google Scholar] [CrossRef]
- Ali, M.M.; Hossain, D.; Al-Imran, A.; Khan, M.S.; Begum, M.; Osman, M.H. Environmental Pollution with Heavy Metals: A Public Health Concern. In Heavy Metals—Their Environmental Impacts and Mitigation; IntechOpen: London, UK, 2021; ISBN 978-1-83968-122-6. [Google Scholar]
- Gorinstein, S.; Jung, S.-T.; Moncheva, S.; Arancibia-Avila, P.; Park, Y.-S.; Kang, S.-G.; Goshev, I.; Trakhtenberg, S.; Namiesnik, J. Partial Characterization of Proteins from Mussel Mytilus Galloprovincialis as a Biomarker of Contamination. Arch. Environ. Contam. Toxicol. 2005, 49, 504–510. [Google Scholar] [CrossRef]
- Santovito, G.; Piccinni, E.; Cassini, A.; Irato, P.; Albergoni, V. Antioxidant Responses of the Mediterranean Mussel, Mytilus Galloprovincialis, to Environmental Variability of Dissolved Oxygen. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2005, 140, 321–329. [Google Scholar] [CrossRef] [PubMed]
- Phillips, D.J.H.; Rainbow, P.S. Strategies of Trace Metal Sequestration in Aquatic Organisms. Mar. Environ. Res. 1989, 28, 207–210. [Google Scholar] [CrossRef]
- Perić, L.; Nerlović, V.; Žurga, P.; Žilić, L.; Ramšak, A. Variations of Biomarkers Response in Mussels Mytilus Galloprovincialis to Low, Moderate and High Concentrations of Organic Chemicals and Metals. Chemosphere 2017, 174, 554–562. [Google Scholar] [CrossRef]
- Lettieri, G.; Notariale, R.; Ambrosino, A.; Di Bonito, A.; Giarra, A.; Trifuoggi, M.; Manna, C.; Piscopo, M. Spermatozoa Transcriptional Response and Alterations in PL Proteins Properties after Exposure of Mytilus Galloprovincialis to Mercury. Int. J. Mol. Sci. 2021, 22, 1618. [Google Scholar] [CrossRef]
- González-Fernández, C.; Albentosa, M.; Campillo, J.A.; Viñas, L.; Romero, D.; Franco, A.; Bellas, J. Effect of Nutritive Status on Mytilus Galloprovincialis Pollution Biomarkers: Implications for Large-Scale Monitoring Programs. Aquat. Toxicol. 2015, 167, 90–105. [Google Scholar] [CrossRef]
- González-Fernández, C.; Albentosa, M.; Campillo, J.A.; Viñas, L.; Franco, A.; Bellas, J. Effect of Mussel Reproductive Status on Biomarker Responses to PAHs: Implications for Large-Scale Monitoring Programs. Aquat. Toxicol. 2016, 177, 380–394. [Google Scholar] [CrossRef]
- Lin, C.Y.; Viant, M.R.; Tjeerdema, R.S. Metabolomics: Methodologies and Applications in the Environmental Sciences. J. Pestic. Sci. 2006, 31, 245–251. [Google Scholar] [CrossRef]
- Hines, A.; Yeung, W.H.; Craft, J.; Brown, M.; Kennedy, J.; Bignell, J.; Stentiford, G.D.; Viant, M.R. Comparison of Histological, Genetic, Metabolomics, and Lipid-Based Methods for Sex Determination in Marine Mussels. Anal. Biochem. 2007, 369, 175–186. [Google Scholar] [CrossRef] [PubMed]
- Hines, A.; Staff, F.J.; Widdows, J.; Compton, R.M.; Falciani, F.; Viant, M.R. Discovery of Metabolic Signatures for Predicting Whole Organism Toxicology. Toxicol. Sci. 2010, 115, 369–378. [Google Scholar] [CrossRef] [PubMed]
- Lankadurai, B.P.; Nagato, E.G.; Simpson, M.J. Environmental Metabolomics: An Emerging Approach to Study Organism Responses to Environmental Stressors. Environ. Rev. 2013, 21, 180–205. [Google Scholar] [CrossRef]
- Pomfret, S.M.; Brua, R.B.; Izral, N.M.; Yates, A.G. Metabolomics for Biomonitoring: An Evaluation of the Metabolome as an Indicator of Aquatic Ecosystem Health. Environ. Rev. 2020, 28, 89–98. [Google Scholar] [CrossRef]
- Izral, N.M.; Brua, R.B.; Culp, J.M.; Yates, A.G. Crayfish Tissue Metabolomes Effectively Distinguish Impacts of Wastewater and Agriculture in Aquatic Ecosystems. Sci. Total Environ. 2021, 760, 143322. [Google Scholar] [CrossRef]
- Patterson, M.A.; Parker, B.C.; Neves, R.J. Glycogen Concentration in the Mantle Tissue of Freshwater Mussels (Bivalvia: Unionidae) During Starvation and Controlled Feeding. Am. Malacol. Bull. 1999, 15, 47–50. [Google Scholar]
- Fritts, A.K.; Peterson, J.T.; Hazelton, P.D.; Bringolf, R.B. Evaluation of Methods for Assessing Physiological Biomarkers of Stress in Freshwater Mussels. Can. J. Fish. Aquat. Sci. 2015, 72, 1450–1459. [Google Scholar] [CrossRef]
- Fritts, A.K.; Peterson, J.T.; Wisniewski, J.M.; Bringolf, R.B. Nonlethal Assessment of Freshwater Mussel Physiological Response to Changes in Environmental Factors. Can. J. Fish. Aquat. Sci. 2015, 72, 1460–1468. [Google Scholar] [CrossRef]
- Haag, W.R.; Berg, D.J.; Garton, D.W.; Farris, J.L. Reduced Survival and Fitness in Native Bivalves in Response to Fouling by the Introduced Zebra Mussel (Dreissena polymorpha) in Western Lake Erie. Can. J. Fish. Aquat. Sci. 1993, 50, 13–19. [Google Scholar] [CrossRef]
- Zhang, L.; Liu, X.; You, L.; Zhou, D.; Wang, Q.; Li, F.; Cong, M.; Li, L.; Zhao, J.; Liu, D.; et al. Benzo(a)Pyrene-Induced Metabolic Responses in Manila Clam Ruditapes Philippinarum by Proton Nuclear Magnetic Resonance (1H NMR) Based Metabolomics. Environ. Toxicol. Pharmacol. 2011, 32, 218–225. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Liu, X.; You, L.; Zhou, D.; Wu, H.; Li, L.; Zhao, J.; Feng, J.; Yu, J. Metabolic Responses in Gills of Manila Clam Ruditapes Philippinarum Exposed to Copper Using NMR-Based Metabolomics. Mar. Environ. Res. 2011, 72, 33–39. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Wang, W.-X. NMR-Based Metabolomic Studies on the Toxicological Effects of Cadmium and Copper on Green Mussels Perna Viridis. Aquat. Toxicol. 2010, 100, 339–345. [Google Scholar] [CrossRef] [PubMed]
- Fasulo, S.; Iacono, F.; Cappello, T.; Corsaro, C.; Maisano, M.; D’Agata, A.; Giannetto, A.; De Domenico, E.; Parrino, V.; Lo Paro, G.; et al. Metabolomic Investigation of Mytilus Galloprovincialis (Lamarck 1819) Caged in Aquatic Environments. Ecotoxicol. Environ. Saf. 2012, 84, 139–146. [Google Scholar] [CrossRef]
- Ji, C.; Wu, H.; Zhou, M.; Zhao, J. Multiple Biomarkers of Biological Effects Induced by Cadmium in Clam Ruditapes Philippinarum. Fish Shellfish. Immunol. 2015, 44, 430–435. [Google Scholar] [CrossRef]
- Carbone, G.; Lettieri, G.; Marinaro, C.; Costabile, M.; Notariale, R.; Bianchi, A.R.; De Maio, A.; Piscopo, M. A Molecular Mechanism to Explain the Nickel-Induced Changes in Protamine-like Proteins and Their DNA Binding Affecting Sperm Chromatin in Mytilus Galloprovincialis: An In Vitro Study. Biomolecules 2023, 13, 520. [Google Scholar] [CrossRef]
- De Guglielmo, V.; Puoti, R.; Notariale, R.; Maresca, V.; Ausió, J.; Troisi, J.; Verrillo, M.; Basile, A.; Febbraio, F.; Piscopo, M. Alterations in the Properties of Sperm Protamine-like II Protein after Exposure of Mytilus Galloprovincialis (Lamarck 1819) to Sub-Toxic Doses of Cadmium. Ecotoxicol. Environ. Saf. 2019, 169, 600–606. [Google Scholar] [CrossRef]
- Lettieri, G.; Mollo, V.; Ambrosino, A.; Caccavale, F.; Troisi, J.; Febbraio, F.; Piscopo, M. Molecular Effects of Copper on the Reproductive System of Mytilus Galloprovincialis. Mol. Reprod Dev. 2019, 86, 1357–1368. [Google Scholar] [CrossRef] [PubMed]
- Lettieri, G.; Carusone, N.; Notariale, R.; Prisco, M.; Ambrosino, A.; Perrella, S.; Manna, C.; Piscopo, M. Morphological, Gene, and Hormonal Changes in Gonads and In-Creased Micrococcal Nuclease Accessibility of Sperm Chromatin Induced by Mercury. Biomolecules 2022, 12, 87. [Google Scholar] [CrossRef]
- Piscopo, M.; Ricciardiello, M.; Palumbo, G.; Troisi, J. Selectivity of Metal Bioaccumulation and Its Relationship with Glutathione S-Transferase Levels in Gonadal and Gill Tissues of Mytilus Galloprovincialis Exposed to Ni (II), Cu (II) and Cd (II). Rend. Fis. Acc. Lincei 2016, 27, 737–748. [Google Scholar] [CrossRef]
- Piscopo, M. Seasonal Dependence of Cadmium Molecular Effects on Mytilus Galloprovincialis (Lamarck, 1819) Protamine-like Protein Properties. Mol. Reprod. Dev. 2019, 86, 1418–1429. [Google Scholar] [CrossRef] [PubMed]
- Troisi, J.; Sarno, L.; Martinelli, P.; Di Carlo, C.; Landolfi, A.; Scala, G.; Rinaldi, M.; D’Alessandro, P.; Ciccone, C.; Guida, M. A Metabolomics-Based Approach for Non-Invasive Diagnosis of Chromosomal Anomalies. Metabolomics 2017, 13, 140. [Google Scholar] [CrossRef]
- Troisi, J.; Landolfi, A.; Sarno, L.; Richards, S.; Symes, S.; Adair, D.; Ciccone, C.; Scala, G.; Martinelli, P.; Guida, M. A Metabolomics-Based Approach for Non-Invasive Screening of Fetal Central Nervous System Anomalies. Metabolomics 2018, 14, 77. [Google Scholar] [CrossRef]
- Troisi, J.; Cavallo, P.; Richards, S.; Symes, S.; Colucci, A.; Sarno, L.; Landolfi, A.; Scala, G.; Adair, D.; Ciccone, C.; et al. Noninvasive Screening for Congenital Heart Defects Using a Serum Metabolomics Approach. Prenat. Diagn. 2021, 41, 743–753. [Google Scholar] [CrossRef]
- Sumner, L.W.; Amberg, A.; Barrett, D.; Beale, M.H.; Beger, R.; Daykin, C.A.; Fan, T.W.-M.; Fiehn, O.; Goodacre, R.; Griffin, J.L.; et al. Proposed Minimum Reporting Standards for Chemical Analysis Chemical Analysis Working Group (CAWG) Metabolomics Standards Initiative (MSI). Metabolomics 2007, 3, 211–221. [Google Scholar] [CrossRef]
- Li, J. The Exploration of the Approach to Data Preparation for Chinese Text Analysis Based on R Language. OALib 2021, 8, 1–8. [Google Scholar] [CrossRef]
- Wehrens, R.; Bloemberg, T.G.; Eilers, P.H.C. Fast Parametric Time Warping of Peak Lists. Bioinformatics 2015, 31, 3063–3065. [Google Scholar] [CrossRef]
- Sysi-Aho, M.; Katajamaa, M.; Yetukuri, L.; Orešič, M. Normalization Method for Metabolomics Data Using Optimal Selection of Multiple Internal Standards. BMC Bioinform. 2007, 8, 93. [Google Scholar] [CrossRef]
- Yi, X.; Xu, Y.; Hu, Q.; Krishnamoorthy, S.; Li, W.; Tang, Z. ASN-SMOTE: A Synthetic Minority Oversampling Method with Adaptive Qualified Synthesizer Selection. Complex Intell. Syst. 2022, 8, 2247–2272. [Google Scholar] [CrossRef]
- Bijlsma, S.; Bobeldijk, I.; Verheij, E.R.; Ramaker, R.; Kochhar, S.; Macdonald, I.A.; van Ommen, B.; Smilde, A.K. Large-Scale Human Metabolomics Studies: A Strategy for Data (Pre-) Processing and Validation. Anal. Chem. 2006, 78, 567–574. [Google Scholar] [CrossRef] [PubMed]
- Gautam, P.K.; Gautam, R.; Banerjee, S.; Chattopadhyaya, M.; Pandey, J. Heavy Metals in the Environment: Fate, Transport, Toxicity and Remediation Technologies. In Heavy Metals: Sources, Toxicity and Remediation Techniques; Nova Science Publishers, Inc.: Hauppauge, NY, USA, 2016; pp. 101–130. ISBN 978-1-63484-740-7. [Google Scholar]
- Mousavi, A.; Pourakbar, L.; Siavash Moghaddam, S. Effects of Malic Acid and EDTA on Oxidative Stress and Antioxidant Enzymes of Okra (Abelmoschus esculentus L.) Exposed to Cadmium Stress. Ecotoxicol. Environ. Saf. 2022, 248, 114320. [Google Scholar] [CrossRef]
- Guo, H.; Chen, H.; Hong, C.; Jiang, D.; Zheng, B. Exogenous Malic Acid Alleviates Cadmium Toxicity in Miscanthus Sacchariflorus through Enhancing Photosynthetic Capacity and Restraining ROS Accumulation. Ecotoxicol. Environ. Saf. 2017, 141, 119–128. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, R.; Ishimaru, Y.; Nakanishi, H.; Nishizawa, N.K. Role of the Iron Transporter OsNRAMP1 in Cadmium Uptake and Accumulation in Rice. Plant Signal Behav. 2011, 6, 1813–1816. [Google Scholar] [CrossRef]
- Xu, P.; Leng, Y.; Zeng, G.; Huang, D.; Lai, C.; Zhao, M.; Wei, Z.; Li, N.; Huang, C.; Zhang, C.; et al. Cadmium Induced Oxalic Acid Secretion and Its Role in Metal Uptake and Detoxification Mechanisms in Phanerochaete Chrysosporium. Appl. Microbiol. Biotechnol. 2015, 99, 435–443. [Google Scholar] [CrossRef]
- Eiss, M.; Schieferstein, G.; Wahl, R. Oxalate in Human Seminal Plasma: Possible Significance and Problems of Oxalate Determination. Fertil. Steril. 2000, 73, 961–966. [Google Scholar] [CrossRef] [PubMed]
- Jiang, M.; Jiang, J.; Li, S.; Li, M.; Tan, Y.; Song, S.; Shu, Q.; Huang, J. Glutamate Alleviates Cadmium Toxicity in Rice via Suppressing Cadmium Uptake and Translocation. J. Hazard. Mater. 2020, 384, 121319. [Google Scholar] [CrossRef]
- Susetyarini, R.E. The Level of Glutamic Acid in the Semen of Male White Rat (Ratus Norwegicus) after Being Treated with Tannin of Pluchea Indica. Procedia Chem. 2015, 14, 152–156. [Google Scholar] [CrossRef][Green Version]
- Juyena, N.S.; Stelletta, C. Seminal Plasma: An Essential Attribute to Spermatozoa. J. Androl. 2012, 33, 536–551. [Google Scholar] [CrossRef] [PubMed]
- Lavanya, M.; Swathi, D.; Archana, S.S.; Ramya, L.; Ranjithkumaran, R.; Krishnaswamy, N.; Singh, S.K.; Krishnappa, B.; Rajendran, D.; Kumar, H.; et al. Supraphysiological Concentration of Urea Affects the Functional Competence of Holstein-Friesian (Bos taurus) Sperm. Theriogenology 2021, 176, 104–114. [Google Scholar] [CrossRef] [PubMed]
- Allahkarami, S.; Atabakhsh, M.; Moradi, M.N.; Ghasemi, H.; Bahmanzadeh, M.; Tayebinia, H. Correlation of Uric Acid, Urea, Ammonia and Creatinine of Seminal Plasma With Semen Parameters and Fertilization Rate in Infertile Couples. Avicenna J. Med. Biochem. 2017, 5, 76–80. [Google Scholar] [CrossRef]
- Calogero, A.E.; Gullo, G.; La Vignera, S.; Condorelli, R.A.; Vaiarelli, A. Myoinositol Improves Sperm Parameters and Serum Reproductive Hormones in Patients with Idiopathic Infertility: A Prospective Double-Blind Randomized Placebo-Controlled Study. Andrology 2015, 3, 491–495. [Google Scholar] [CrossRef] [PubMed]
- Kushawaha, B.; Yadav, R.S.; Swain, D.K.; Kumari, P.; Kumar, A.; Yadav, B.; Anand, M.; Yadav, S.; Singh, D.; Garg, S.K. Collapsed Mitochondrial Cristae in Goat Spermatozoa Due to Mercury Result in Lethality and Compromised Motility along with Altered Kinematic Patterns. Sci. Rep. 2021, 11, 646. [Google Scholar] [CrossRef] [PubMed]
- Condorelli, R.A.; Barbagallo, F.; Calogero, A.E.; Cannarella, R.; Crafa, A.; La Vignera, S. D-Chiro-Inositol Improves Sperm Mitochondrial Membrane Potential: In Vitro Evidence. J. Clin. Med. 2020, 9, 1373. [Google Scholar] [CrossRef]
- Metcalf, E.S.; Dideon, B.A.; Blehr, R.; Schlimgen, T.; Bertrand, W.; Varner, D.D.; Teague, S.R.; Hausman, M.S. Effects of DMSO and L-Ergothioneine on Post-Thaw Semen Parameters in Stallions: Preliminary Results. Anim. Reprod. Sci. 2008, 107, 332–333. [Google Scholar] [CrossRef]
- Hunter, T. Why Nature Chose Phosphate to Modify Proteins. Philos. Trans. R. Soc. B Biol. Sci. 2012, 367, 2513–2516. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Zhang, Y.; Wu, X.; Wan, D.; Yin, Y. Effects of Dietary Serine Supplementation on Intestinal Integrity, Inflammation and Oxidative Status in Early-Weaned Piglets. Cell. Physiol. Biochem. 2018, 48, 993–1002. [Google Scholar] [CrossRef]
- Zhou, X.; He, L.; Wu, C.; Zhang, Y.; Wu, X.; Yin, Y. Serine Alleviates Oxidative Stress via Supporting Glutathione Synthesis and Methionine Cycle in Mice. Mol. Nutr. Food Res. 2017, 61. [Google Scholar] [CrossRef]
- Alfaro, A.C.; Young, T. Showcasing Metabolomic Applications in Aquaculture: A Review. Rev. Aquac. 2018, 10, 135–152. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lettieri, G.; Marinaro, C.; Notariale, R.; Perrone, P.; Lombardi, M.; Trotta, A.; Troisi, J.; Piscopo, M. Impact of Heavy Metal Exposure on Mytilus galloprovincialis Spermatozoa: A Metabolomic Investigation. Metabolites 2023, 13, 943. https://doi.org/10.3390/metabo13080943
Lettieri G, Marinaro C, Notariale R, Perrone P, Lombardi M, Trotta A, Troisi J, Piscopo M. Impact of Heavy Metal Exposure on Mytilus galloprovincialis Spermatozoa: A Metabolomic Investigation. Metabolites. 2023; 13(8):943. https://doi.org/10.3390/metabo13080943
Chicago/Turabian StyleLettieri, Gennaro, Carmela Marinaro, Rosaria Notariale, Pasquale Perrone, Martina Lombardi, Alessio Trotta, Jacopo Troisi, and Marina Piscopo. 2023. "Impact of Heavy Metal Exposure on Mytilus galloprovincialis Spermatozoa: A Metabolomic Investigation" Metabolites 13, no. 8: 943. https://doi.org/10.3390/metabo13080943
APA StyleLettieri, G., Marinaro, C., Notariale, R., Perrone, P., Lombardi, M., Trotta, A., Troisi, J., & Piscopo, M. (2023). Impact of Heavy Metal Exposure on Mytilus galloprovincialis Spermatozoa: A Metabolomic Investigation. Metabolites, 13(8), 943. https://doi.org/10.3390/metabo13080943