
Decrease in the Ratio proBDNF/BDNF in the Urine of Aging Female Patients with OAB




Abstract

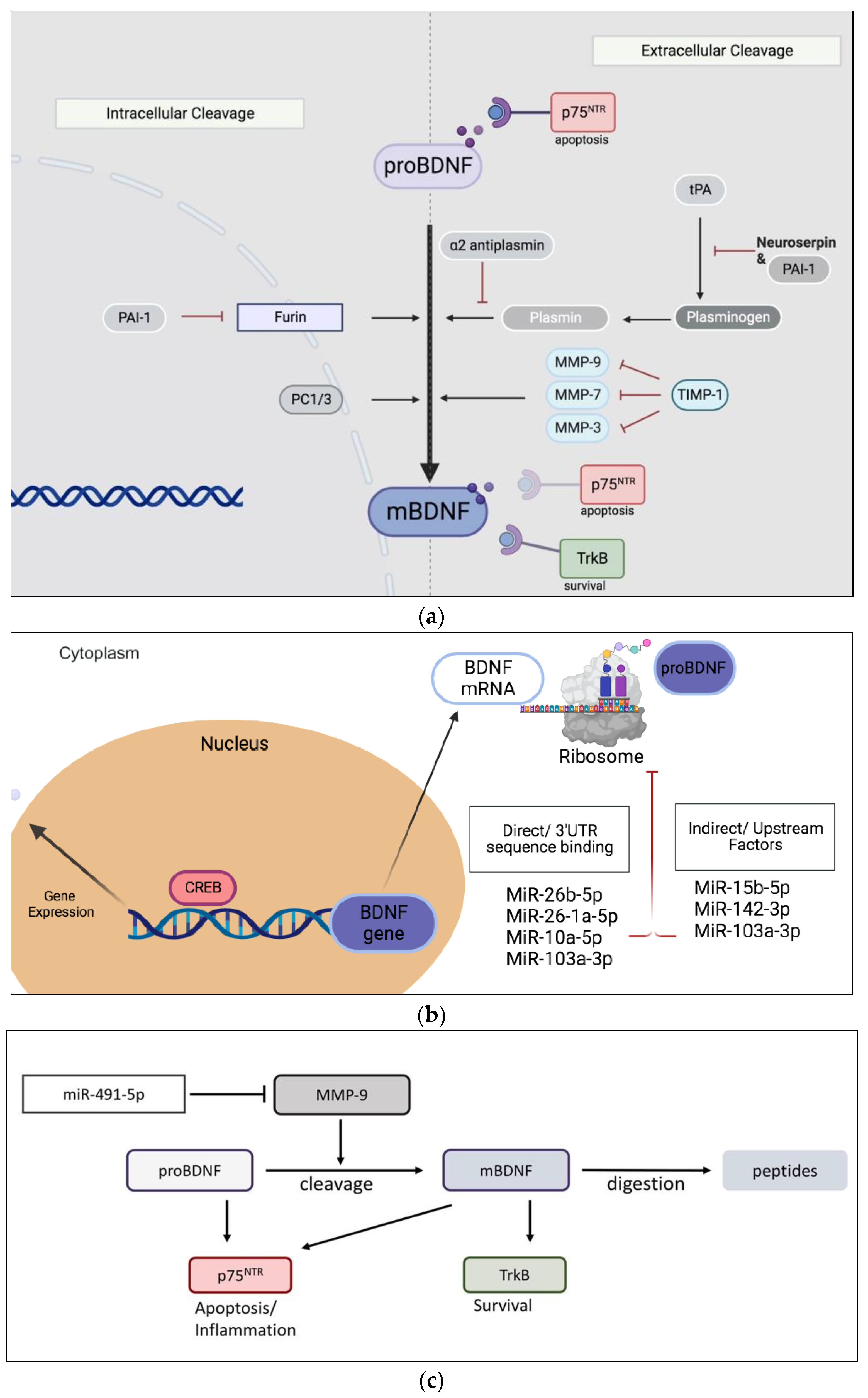
1. Introduction
2. Materials and Methods
2.1. Patient Profiles
2.2. Demographic and Clinical Differences
2.3. Collection and Preparation of Urine Samples
2.4. MiRNAs Isolation from Urine Samples
2.5. MicroRNA Poly-Adenylation and Synthesis of cDNA
2.6. Quantitative PCR (qPCR)
2.7. Enzyme-Linked Immunosorbent Assay (ELISA) and Enzymatic Kits
2.8. Statistics
3. Results
3.1. Subject Characteristics
3.2. Biochemical Urinalysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Haylen, B.T.; De Ridder, D.; Freeman, R.M.; Swift, S.E.; Berghmans, B.; Lee, J.; Monga, A.; Petri, E.; Rizk, D.E.; Sand, P.K. An International Urogynecological Association (IUGA)/International Continence Society (ICS) joint report on the terminology for female pelvic floor dysfunction. Neurourol. Urodyn. Off. J. Int. Cont. Soc. 2010, 29, 4–20. [Google Scholar] [CrossRef] [PubMed]
- Lightner, D.J.; Gomelsky, A.; Souter, L.; Vasavada, S.P. Diagnosis and Treatment of Overactive Bladder (Non-Neurogenic) in Adults: AUA/SUFU Guideline Amendment 2019. J. Urol. 2019, 202, 558–563. [Google Scholar] [CrossRef]
- Irwin, D.E.; Milsom, I.; Hunskaar, S.; Reilly, K.; Kopp, Z.; Herschorn, S.; Coyne, K.; Kelleher, C.; Hampel, C.; Artibani, W.; et al. Population-Based Survey of Urinary Incontinence, Overactive Bladder, and Other Lower Urinary Tract Symptoms in Five Countries: Results of the EPIC Study. Eur. Urol. 2006, 50, 1306–1315. [Google Scholar] [CrossRef]
- Reynolds, W.S.; Fowke, J.; Dmochowski, R. The burden of overactive bladder on US public health. Curr. Bladder Dysfunct. Rep. 2016, 11, 8–13. [Google Scholar] [CrossRef] [PubMed]
- Irwin, D.E.; Kopp, Z.S.; Agatep, B.; Milsom, I.; Abrams, P. Worldwide prevalence estimates of lower urinary tract symptoms, overactive bladder, urinary incontinence and bladder outlet obstruction. BJU Int. 2011, 108, 1132–1138. [Google Scholar] [CrossRef]
- Al Edwan, G.; Abdelazim, M.S.; Salhab, S.E.; Jamal, Y.M.; Soliman, M.A. The Prevalence of Overactive Bladder Symptoms in Women in Algeria, Egypt, Jordan and Lebanon: A Cross-Sectional Population-Based Survey. Adv. Ther. 2020, 38, 1155–1167. [Google Scholar] [CrossRef]
- Wang, Y.; Xu, K.; Hu, H.; Zhang, X.; Wang, X.; Na, Y.; Kang, X. Prevalence, risk factors, and impact on health related quality of life of overactive bladder in China. Neurourol. Urodyn. 2011, 30, 1448–1455. [Google Scholar] [CrossRef] [PubMed]
- Lo, C.-W.; Wu, M.-Y.; Yang, S.S.-D.; Jaw, F.-S.; Chang, S.-J. Comparing the Efficacy of OnabotulinumtoxinA, Sacral Neuromodulation, and Peripheral Tibial Nerve Stimulation as Third Line Treatment for the Management of Overactive Bladder Symptoms in Adults: Systematic Review and Network Meta-Analysis. Toxins 2020, 12, 128. [Google Scholar] [CrossRef]
- Peyronnet, B.; Mironska, E.; Chapple, C.; Cardozo, L.; Oelke, M.; Dmochowski, R.; Amarenco, G.; Gamé, X.; Kirby, R.; Van Der Aa, F.; et al. A Comprehensive Review of Overactive Bladder Pathophysiology: On the Way to Tailored Treatment. Eur. Urol. 2019, 75, 988–1000. [Google Scholar] [CrossRef] [PubMed]
- Yoshimura, N.; Ogawa, T.; Miyazato, M.; Kitta, T.; Furuta, A.; Chancellor, M.B.; Tyagi, P. Neural Mechanisms Underlying Lower Urinary Tract Dysfunction. Korean J. Urol. 2014, 55, 81–90. [Google Scholar] [CrossRef]
- Gibon, J.; Barker, P.A. Neurotrophins and Proneurotrophins: Focus on Synaptic Activity and Plasticity in the Brain. Neurosci. 2017, 23, 587–604. [Google Scholar] [CrossRef]
- Lommatzsch, M.; Braun, A.; Mannsfeldt, A.; Botchkarev, V.A.; Botchkareva, N.V.; Paus, R.; Fischer, A.; Lewin, G.R.; Renz, H. Abundant Production of Brain-Derived Neurotrophic Factor by Adult Visceral Epithelia: Implications for Paracrine and Target-Derived Neurotrophic Functions. Am. J. Pathol. 1999, 155, 1183–1193. [Google Scholar] [CrossRef]
- Lee, R.; Kermani, P.; Teng, K.; Hempstead, B.L. Regulation of Cell Survival by Secreted Proneurotrophins. Science 2001, 294, 1945–1948. [Google Scholar] [CrossRef] [PubMed]
- Allen, S.J.; Dawbarn, D. Clinical relevance of the neurotrophins and their receptors. Clin. Sci. 2006, 110, 175–191. [Google Scholar] [CrossRef] [PubMed]
- Coelho, A.; Oliveira, R.; Antunes-Lopes, T.; Cruz, C.D. Partners in Crime: NGF and BDNF in Visceral Dysfunction. Curr. Neuropharmacol. 2019, 17, 1021–1038. [Google Scholar] [CrossRef]
- Kuzniewska, B.; Rejmak, E.; Malik, A.R.; Jaworski, J.; Kaczmarek, L.; Kalita, K. Brain-Derived Neurotrophic Factor Induces Matrix Metalloproteinase 9 Expression in Neurons via the Serum Response Factor/c-Fos Pathway. Mol. Cell. Biol. 2013, 33, 2149–2162. [Google Scholar] [CrossRef]
- Mossa, A.H.; Cammisotto, P.G.; Shamout, S.; Campeau, L. Imbalance of nerve growth factor metabolism in aging women with overactive bladder syndrome. World J. Urol. 2021, 39, 2055–2063. [Google Scholar] [CrossRef]
- Mossa, A.H.; Shamout, S.; Cammisotto, P.; Campeau, L. Urinary metabolomics predict the severity of overactive bladder syndrome in an aging female population. Int. Urogynecology J. 2020, 31, 1023–1031. [Google Scholar] [CrossRef]
- Wishart, D.S.; Knox, C.; Guo, A.C.; Eisner, R.; Young, N.; Gautam, B.; Hau, D.D.; Psychogios, N.; Dong, E.; Bouatra, S. HMDB: A knowledgebase for the human metabolome. Nucleic Acids Res. 2009, 37 (Suppl. S1), D603–D610. [Google Scholar] [CrossRef]
- Bouatra, S.; Aziat, F.; Mandal, R.; Guo, A.C.; Wilson, M.R.; Knox, C.; Bjorndahl, T.C.; Krishnamurthy, R.; Saleem, F.; Liu, P. The human urine metabolome. PLoS ONE 2013, 8, e73076. [Google Scholar] [CrossRef] [PubMed]
- Blaivas, J.G.; Panagopoulos, G.; Weiss, J.P.; Somaroo, C. Validation of the overactive bladder symptom score. J. Urol. 2007, 178, 543–547. [Google Scholar] [CrossRef]
- Uebersax, J.; Wyman, J.; Shumaker, S.; McClish, D.; Fantl, J. Continence Program for Women Research Group Short forms to assess life quality and symptom distress for urinary incontinence in women: The incontinence impact questionnaire and the urogenital distress inventory. Neurourol. Urodyn. 1995, 14, 131–139. [Google Scholar] [CrossRef] [PubMed]
- Braicu, C.; Buiga, R.; Cojocneanu, R.; Buse, M.; Raduly, L.; Pop, L.A.; Chira, S.; Budisan, L.; Jurj, A.; Ciocan, C.; et al. Connecting the dots between different networks: miRNAs associated with bladder cancer risk and progression. J. Exp. Clin. Cancer Res. 2019, 38, 433. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Su, J.; Kong, W.; Fang, Z.; Li, Y.; Huang, Z.; Wen, J.; Wang, Y. Roles of miR-10a-5p and miR-103a-3p, Regulators of BDNF Expression in Follicular Fluid, in the Outcomes of IVF-ET. Front. Endocrinol. 2021, 12, 637384. [Google Scholar] [CrossRef]
- Caputo, V.; Sinibaldi, L.; Fiorentino, A.; Parisi, C.; Catalanotto, C.; Pasini, A.; Cogoni, C.; Pizzuti, A. Brain Derived Neurotrophic Factor (BDNF) Expression Is Regulated by MicroRNAs miR-26a and miR-26b Allele-Specific Binding. PLoS ONE 2011, 6, e28656. [Google Scholar] [CrossRef] [PubMed]
- Gupta, N.; Jadhav, S.; Tan, K.-L.; Saw, G.; Mallilankaraman, K.B.; Dheen, S.T. miR-142-3p Regulates BDNF Expression in Activated Rodent Microglia Through Its Target CAMK2A. Front. Cell. Neurosci. 2020, 14, 132. [Google Scholar] [CrossRef] [PubMed]
- Boone, D.K.; Weisz, H.A.; Bi, M.; Falduto, M.T.; Torres, K.E.O.; Willey, H.E.; Volsko, C.M.; Kumar, A.M.; Micci, M.-A.; Dewitt, D.S.; et al. Evidence linking microRNA suppression of essential prosurvival genes with hippocampal cell death after traumatic brain injury. Sci. Rep. 2017, 7, 6645. [Google Scholar] [CrossRef]
- Guo, J.; Luo, C.; Yang, Y.; Dong, J.; Guo, Z.; Yang, J.; Lian, H.; Ye, C.; Liu, M. MiR-491-5p, as a Tumor Suppressor, Prevents Migration and Invasion of Breast Cancer by Targeting ZNF-703 to Regulate AKT/mTOR Pathway. Cancer Manag. Res. 2021, 13, 403–413. [Google Scholar] [CrossRef]
- Jeanneteau, F.; Chao, M. Are BDNF and glucocorticoid activities calibrated? Neuroscience 2013, 239, 173–195. [Google Scholar] [CrossRef]
- Meeker, R.B.; Crooks, A.M. The new wave of p75 neurotrophin receptor targeted therapies. Neural Regen. Res. 2022, 17, 95–96. [Google Scholar] [CrossRef]
- Luu, B.E.; Mossa, A.H.; Cammisotto, P.G.; Saragovi, H.U.; Campeau, L. Modulation of diabetic kidney disease markers by an antagonist of p75NTR in streptozotocin-treated mice. Gene 2022, 838, 146729. [Google Scholar] [CrossRef]
- Tsiapakidou, S.; Apostolidis, A.; Pantazis, K.; Grimbizis, G.F.; Mikos, T. The use of urinary biomarkers in the diagnosis of overactive bladder in female patients. A systematic review and meta-analysis. Int. Urogynecology J. 2021, 32, 3143–3155. [Google Scholar] [CrossRef] [PubMed]
- Rada, M.P.; Ciortea, R.; Măluţan, A.M.; Doumouchtsis, S.K.; Bucuri, C.E.; Clim, A.; Roman, A.; Mihu, D. The profile of urinary biomarkers in overactive bladder. Neurourol. Urodyn. 2020, 39, 2305–2313. [Google Scholar] [CrossRef] [PubMed]
- Utomo, E.; Barus, M.N.G.; Tala, M.R.Z. Brain-derived neurotrophic factor, nerve growth factor, and high sensitivity C-reactive protein levels in urine in overactive bladder patients: A meta-analysis. F1000Research 2021, 10, 747. [Google Scholar] [CrossRef]
- Antunes-Lopes, T.; Pinto, R.; Barros, S.C.; Botelho, F.; Silva, C.M.; Cruz, C.D.; Cruz, F. Urinary Neurotrophic Factors in Healthy Individuals and Patients with Overactive Bladder. J. Urol. 2013, 189, 359–365. [Google Scholar] [CrossRef]
- Wang, L.-W.; Han, X.-M.; Chen, C.-H.; Ma, Y.; Hai, B. Urinary brain-derived neurotrophic factor: A potential biomarker for objective diagnosis of overactive bladder. Int. Urol. Nephrol. 2014, 46, 341–347. [Google Scholar] [CrossRef]
- Farach, F.J.; Pruitt, L.D.; Jun, J.J.; Jerud, A.B.; Zoellner, L.A.; Roy-Byrne, P.P. Pharmacological treatment of anxiety disorders: Current treatments and future directions. J. Anxiety Disord. 2012, 26, 833–843. [Google Scholar] [CrossRef] [PubMed]
- Lanni, C.; Stanga, S.; Racchi, M.; Govoni, S. The Expanding Universe of Neurotrophic Factors: Therapeutic Potential in Aging and Age-Associated Disorders. Curr. Pharm. Des. 2010, 16, 698–717. [Google Scholar] [CrossRef]
- Berridge, M.J. Smooth muscle cell calcium activation mechanisms. J. Physiol. 2008, 586, 5047–5061. [Google Scholar] [CrossRef]
- Liao, D.; Zhang, C.; Liu, N.; Cao, L.; Wang, C.; Feng, Q.; Yao, D.; Long, M.; Jiang, P. Involvement of neurotrophic signaling in doxorubicin-induced cardiotoxicity. Exp. Ther. Med. 2020, 19, 1129–1135. [Google Scholar] [CrossRef] [PubMed]
- Ioannou, M.S.; Fahnestock, M. ProNGF, but Not NGF, Switches from Neurotrophic to Apoptotic Activity in Response to Reductions in TrkA Receptor Levels. Int. J. Mol. Sci. 2017, 18, 599. [Google Scholar] [CrossRef]
- Cechova, K.; Angelucci, F.; Markova, H.; Nikolai, T.; Matuskova, V.; Laczó, J.; Nedelska, Z.; Vyhnalek, M.; Hort, J. Ratio of serum proBDNF to BDNF and its association with cognitive performance and brain morphometry in mild cognitive impairment: Neuropsychology/Neuropsychological correlates of physiologic markers of cognitive decline/Dementia. Alzheimer’s Dement. 2020, 16, e046340. [Google Scholar] [CrossRef]
- Ridnour, L.A.; Windhausen, A.N.; Isenberg, J.S.; Yeung, N.; Thomas, D.D.; Vitek, M.P.; Roberts, D.D.; Wink, D.A. Nitric oxide regulates matrix metalloproteinase-9 activity by guanylyl-cyclase-dependent and -independent pathways. Proc. Natl. Acad. Sci. USA 2007, 104, 16898–16903. [Google Scholar] [CrossRef]
- Khan, K.M.F.; Kothari, P.; Du, B.; Dannenberg, A.J.; Falcone, D.J. Matrix Metalloproteinase-Dependent Microsomal Prostaglandin E Synthase-1 Expression in Macrophages: Role of TNF-α and the EP4 Prostanoid Receptor. J. Immunol. 2012, 188, 1970–1980. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| CTR | OAB Group | p Value | |
|---|---|---|---|
| Demographic and serum analysis: | |||
| Age (years) | 56.25 (5.22) | 68.9 (11.38) | <0.001 |
| BMI (kg/m2) | 29.75 (7.65) | 28.82 (5.45) | ns |
| eGFR (mL/min/1.73 m2) | 98.5 (14.52) | 76 (19.78) | <0.001 |
| HOMA-IR | 2.13 (1.03) | 3.11 (1.18) | 0.020 |
| Total Cholesterol/HDL | 3.50 (1.18) | 3.23 (0.81) | ns |
| Questionnaires’ scores: | |||
| OABSS (0–28) | 7.3 (3.56) | 17.45 (4.45) | <0.001 |
| ICIQ--SF (0–22) | 3.26 (3.98) | 8.05 (3.83) | <0.001 |
| IIQ-7 (0–100) | 2.4 (5.2) | 28.9 (23.2) | <0.001 |
| Voiding diary parameters: | |||
| 24 h frequency | 9.15 (2.28) | 11.4 (3.03) | 0.012 |
| Daytime frequency | 8.5 (2.04) | 9.5 (2.09) | ns |
| Night frequency | 0.65 (0.81) | 1.9 (1.71) | 0.005 |
| 24 h voiding volume (mL) | 2705 (2346.02) | 1859.6 (865.37) | ns |
| Night voiding volume (mL) | 495.25 (253.88) | 449.75 (270.77) | ns |
| Mean voided volume (mL) | 322.25 (311.1) | 167.36 (75.2) | 0.037 |
| Maximum voided volume (mL) | 480.75 (193.44) | 327.25 (126.7) | 0.005 |
| Ctl Group | OAB Group | p Value | |
|---|---|---|---|
| BDNF (pg/mg creat) | 19.67 ± 4.25 | 30.92 ± 6.84 | 0.265 |
| proBDNF (pg/mg creat) | 1543.5 ± 381.7 | 733.1 ± 131.8 | 0.231 |
| proBDNF/BDNF | 27.24 ± 3.09 | 16.89 ± 3.02 | 0.023 |
| MMP-9 (ng/mg creat) | 0.325 ± 0.124 | 1.802 ± 0.481 | 0.035 |
| proMMP-9 (ng/mg creat) | 0.391 ± 0.162 | 0.646 ± 0.216 | 0.301 |
| MMP-9/pro-MMP-9 | 2.401 ± 0.60 | 3.446 ± 0.61 | 0.251 |
| Confounders | CTR | OAB Group | p Value | |
|---|---|---|---|---|
| BDNF (pg/mg creat) | Age | 27.51 (20.63–34.39) | 26.87 (19.99–33.76) | 0.952 |
| HOMA-IR | 20.78 (13.95–27.6) | 27.66 (20.59–34.74) | 0.509 | |
| eGFR | 22.97 (16.1–29.9) | 31.41 (13.99–48.84) | 0.431 | |
| proBDNF (pg/mg creat) | Age | 1528 (1202–1853) | 749 (424–1074) | 0.133 |
| HOMA-IR | 1753 (1390–2117) | 729 (352–1105) | 0.072 | |
| eGFR | 1488 (1169–1807) | 789 (470–1108) | 0.162 | |
| proBDNF/BDNF (mol/mol) | Age | 24.81 (21.44–28.18) | 19.31 (15.94–22.68) | 0.302 |
| HOMA-IR | 29.45 (26.08–32.82) | 18.89 (15.4–22.38) | 0.047 | |
| eGFR | 27.59 (24.18–31.01) | 16.53 (13.11–19.95) | 0.042 | |
| MMP-9 (ng/mg creat) | Age | 0.530 (0.082–0.978) | 1.645 (1.225–2.065) | 0.111 |
| HOMA-IR | 0.581 (0.108–1.054) | 1.677 (1.204–2.150) | 0.128 | |
| eGFR | 0.445 (0.014–0.876) | 1.722 (1.316–2.128) | 0.054 | |
| proMMP-9 (ng/mg creat) | Age | 0.583 (0.374–0.792) | 0.455 (0.246–0.664) | 0.697 |
| HOMA-IR | 0.387 (0.170–0.604) | 0.539 (0.314–0.764) | 0.645 | |
| eGFR | 0.331 (0.117–0.545) | 0.707 (0.376–1.038) | 0.262 | |
| MMP-9/proMMP-9 | Age | 2.419 (1.682–3.156) | 3.431 (2.740–4.122) | 0.367 |
| HOMA-IR | 2.727 (1.889–3.565) | 3.764 (2.957–4.571) | 0.405 | |
| eGFR | 3.010 (2.342–3.678) | 2.900 (2.271–3.529) | 0.913 |
| Correlation | p Value | ||
|---|---|---|---|
| BDNF (pg/mg creat) | OABSS | 0.035 | 0.828 |
| ICIQ-SF | 0.165 | 0.314 | |
| IIQ-7 | 0.144 | 0.377 | |
| proBDNF (pg/mg creat) | OABSS | −0.336 | 0.034 |
| ICIQ-SF | −0.176 | 0.285 | |
| IIQ-7 | −0.267 | 0.096 | |
| proBDNF/BDNF (mol/mol) | OABSS | −0.392 | 0.012 |
| ICIQ-SF | −0.290 | 0.073 | |
| IIQ-7 | −0.391 | 0.013 | |
| MMP-9 (ng/mg creat) | OABSS | 0.259 | 0.117 |
| ICIQ-SF | 0.307 | 0.065 | |
| IIQ-7 | 0.207 | 0.212 |
| Ctl | OAB | p Value | |
| miR-26b-5p | 0.891 (0.423, 1.864) | 1.865 (0.0830, 2.842) | 0.99 |
| miR-26a-5p | 0.808 (0.495, 1.911) | 1.634 (0.0852, 2.911) | 0.989 |
| miR-10a-5p | 0.884 (0.420, 3.489) | 0.344 (0.101, 1.083) | 0.108 |
| miR-103a-3p | 1.200 (0.500, 2.156) | 0.592 (0.0570, 1.105) | 0.102 |
| miR-15b-5p | 0.825 (0.494, 3.076) | 1.504 (0.307, 4.070) | 0.841 |
| miR-142-3p | 0.438 (0.0728, 1.511) | 0.449 (0.267, 1.930) | 0.369 |
| miR-202-3p | 1.300 (0.318, 2.750) | 0.257 (0.146, 2.531) | 0.211 |
| miR-124-5p | 0.939 (0.382, 4.121) | 1.356 (0.259, 2.234) | 0.813 |
| miR-152-5p | 1.362 (1.166, 3.902) | 0.721 (0.298, 5.568) | 0.792 |
| miR-491-5p | 0.533 (0.302, 1.643) | 0.122 (0.0221, 0.392) | 0.008 |
| MMP-7 (ng/mg creat) | 0.232 (0.140, 0.455) | 0.443 (0.228, 1.02) | 0.079 |
| MMP-3 (mU/mg creat) | 0.0147 (0.00584, 0.0262) | 0.0104 (0.00038, 0.235) | 0.583 |
| Plasmin (ng/mg creat) | 18.1 (14.57, 30.90) | 27.65 (15.65, 38.40) | 0.512 |
| Cortisol (ng/mg creat) | 38.18 (29.72, 53.74) | 35.29 (17.89, 56.70) | 0.529 |
| p75ECD (ng/mg creat) | 2.471 (2.149, 2.855) | 2.851 (2.623, 3.593) | 0.035 |
| Sortilin (pg/mg creat) | 1710 (737, 2984) | 1752 (1286, 3266) | 0.398 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Covarrubias, C.; Cammisotto, P.G.; Shamout, S.; Campeau, L. Decrease in the Ratio proBDNF/BDNF in the Urine of Aging Female Patients with OAB. Metabolites 2023, 13, 723. https://doi.org/10.3390/metabo13060723
Covarrubias C, Cammisotto PG, Shamout S, Campeau L. Decrease in the Ratio proBDNF/BDNF in the Urine of Aging Female Patients with OAB. Metabolites. 2023; 13(6):723. https://doi.org/10.3390/metabo13060723
Chicago/Turabian StyleCovarrubias, Claudia, Philippe G Cammisotto, Samer Shamout, and Lysanne Campeau. 2023. "Decrease in the Ratio proBDNF/BDNF in the Urine of Aging Female Patients with OAB" Metabolites 13, no. 6: 723. https://doi.org/10.3390/metabo13060723
APA StyleCovarrubias, C., Cammisotto, P. G., Shamout, S., & Campeau, L. (2023). Decrease in the Ratio proBDNF/BDNF in the Urine of Aging Female Patients with OAB. Metabolites, 13(6), 723. https://doi.org/10.3390/metabo13060723