
Certain Tomato Root Exudates Induced by Pseudomonas stutzeri NRCB010 Enhance Its Rhizosphere Colonization Capability



Abstract
1. Introduction
2. Materials and Methods
2.1. PGPR Strain and Plant Materials
2.2. Effects of NRCB010 on Tomato Growth in Hydroponic Conditions
2.3. Tomato Root Exudates Collection and Component Analysis
2.3.1. Root Exudates Collection and Component Measurements
2.3.2. Root Exudates Component Identification
2.4. Effects of Root Exudates and Certain Metabolites on NRCB010 Growth
2.4.1. Root Exudates and Certain Metabolites Preparation
NRCB010 Biomass
Diameter of NRCB010 Swarming Area
Biofilm Formation
Chemotaxis
Colonization
2.5. Statistical Analyses
3. Results
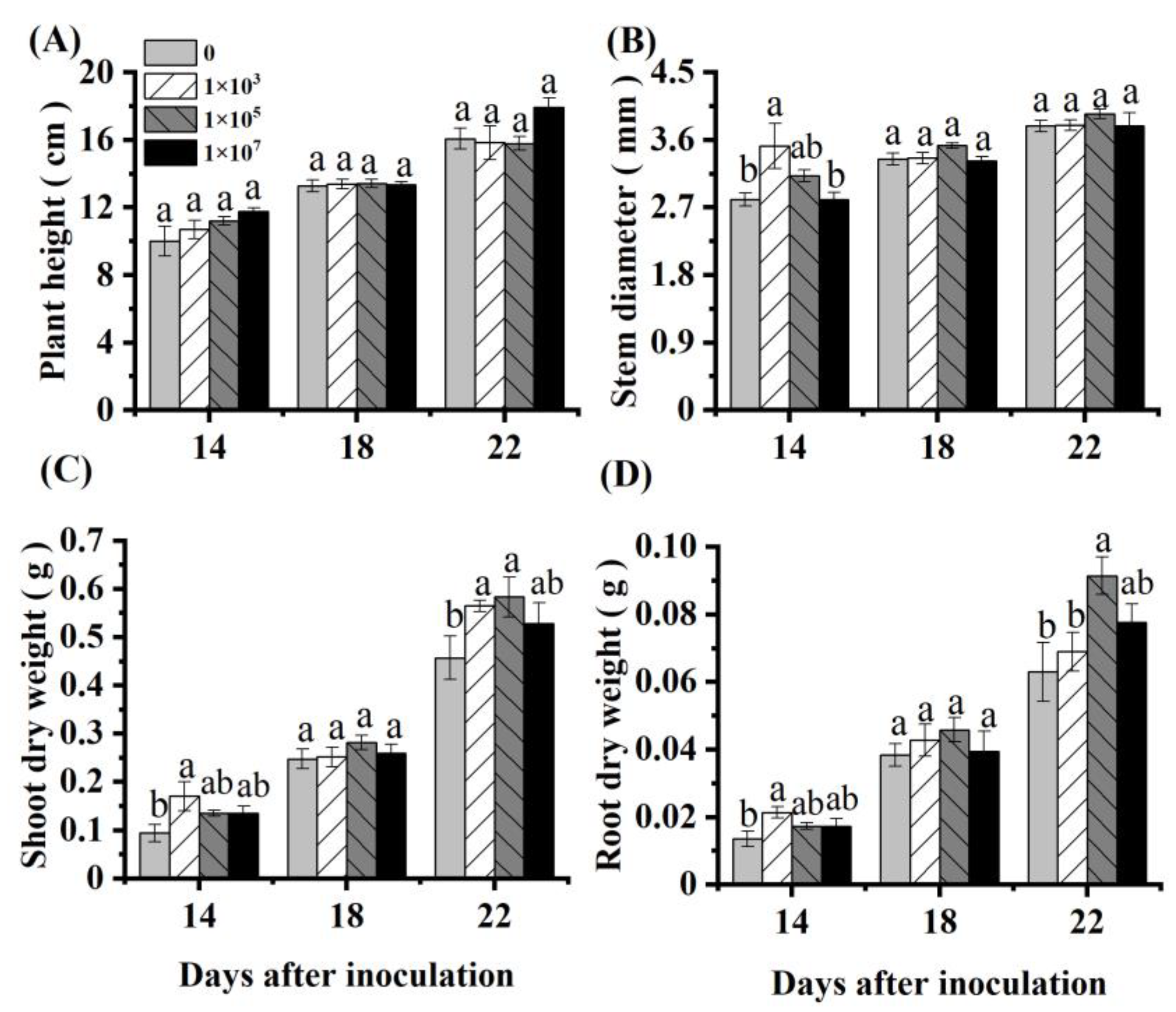
3.1. Effects of NRCB010 on Tomato Growth in Hydroponic Conditions
Effects of NRCB010 on Tomato Root Exudate (RE) Component
3.2. Effects of Root Exudates on NRCB010 Growth
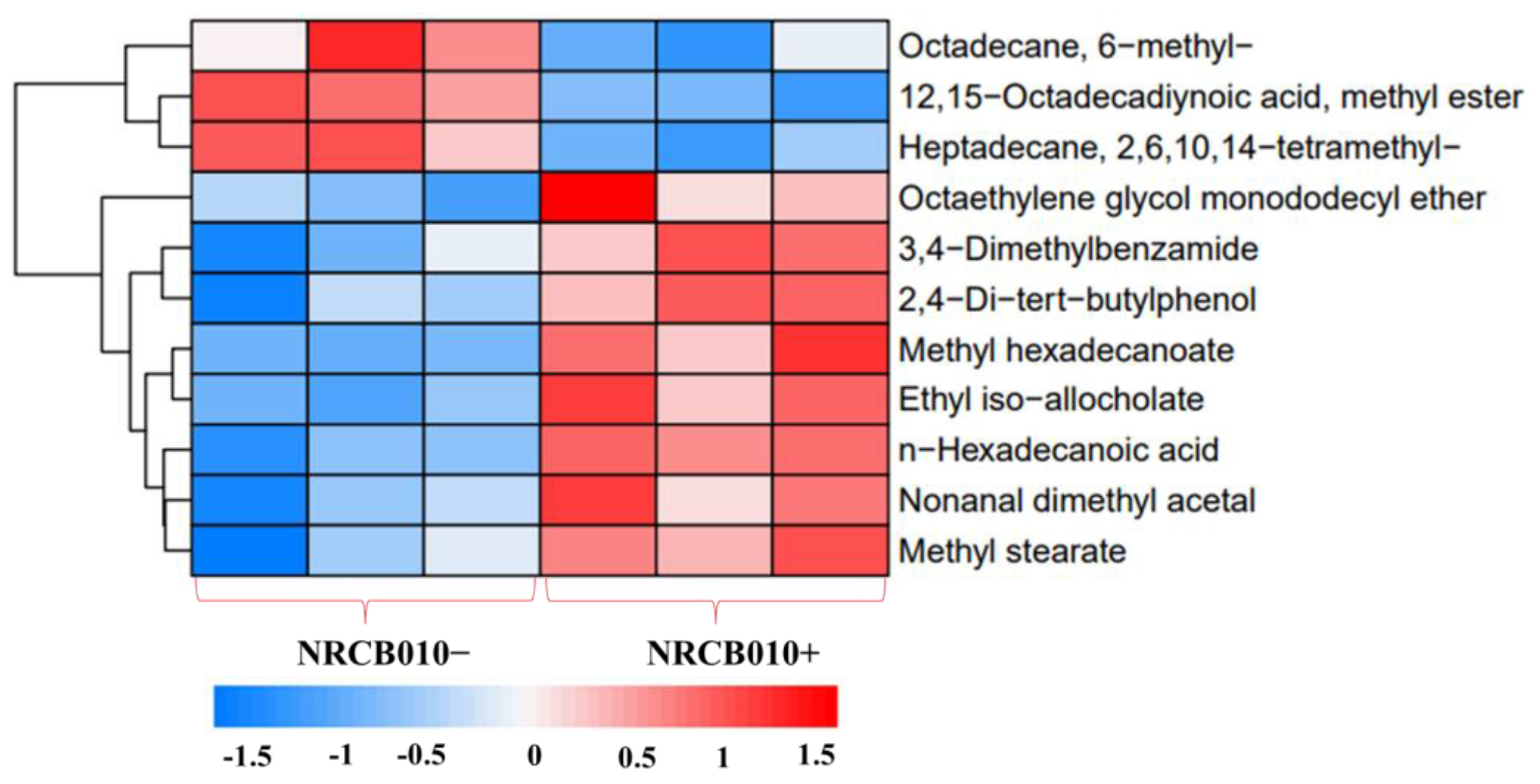
3.3. Changes in Tomato Root Exudate Profile in Response to NRCB010
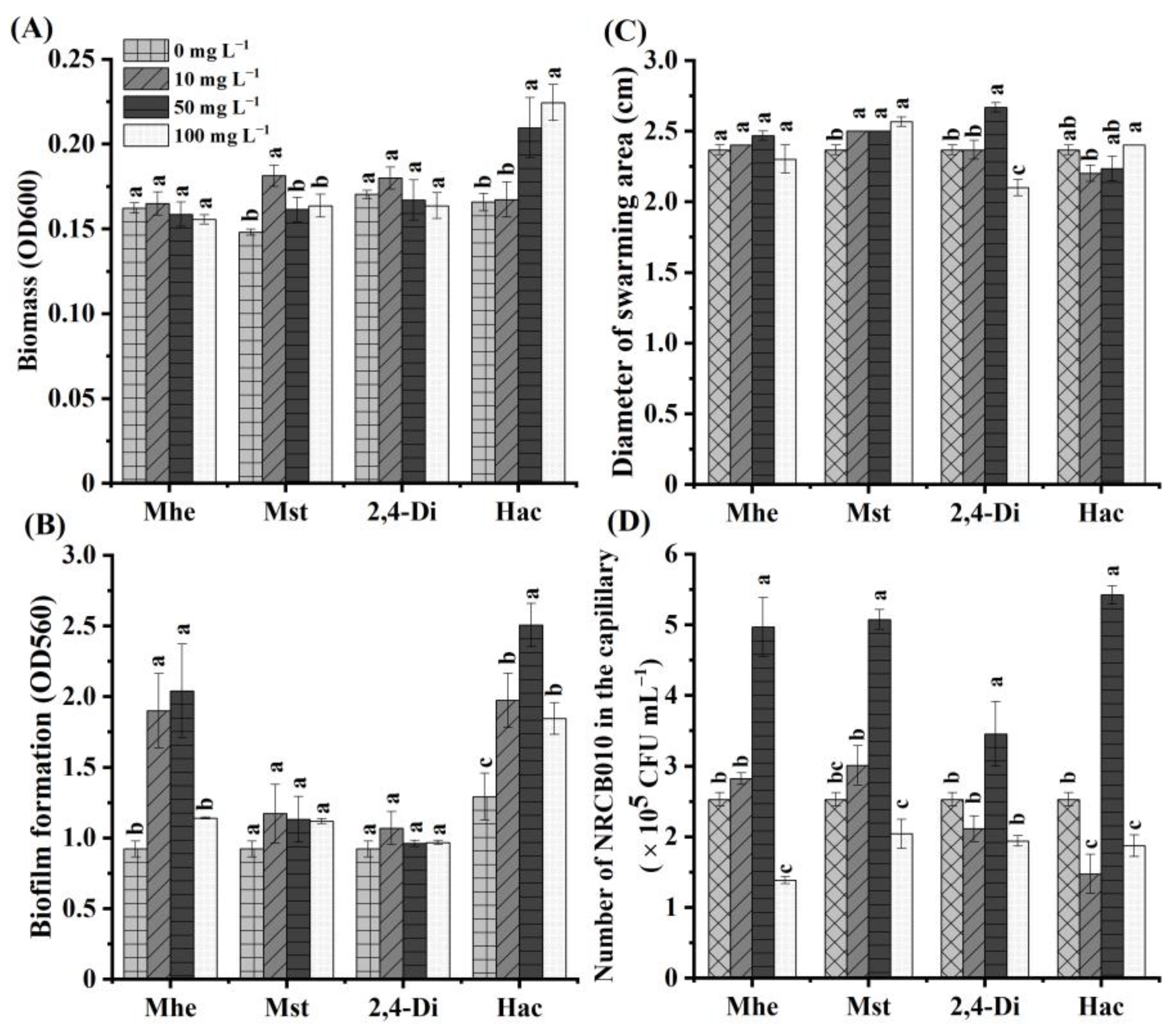
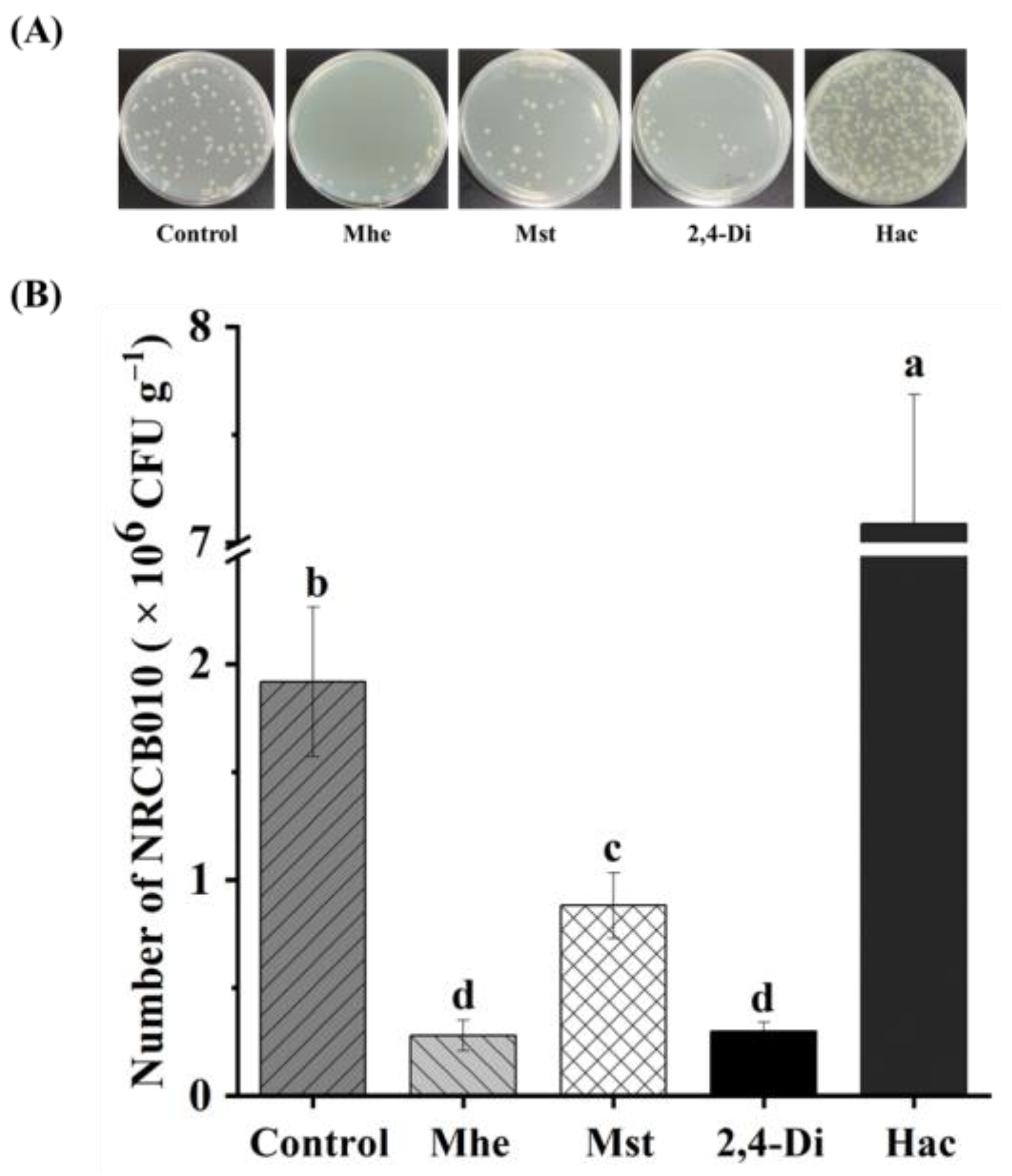
3.4. Effects of Four Root Exudate Metabolites on NRCB010 Growth
4. Discussion
5. Conclusions
6. Patents
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Trivedi, P.; Leach, J.E.; Tringe, S.G.; Sa, T.M.; Singh, B.K. Plant-microbiome interactions: From community assembly to plant health. Nat. Rev. Microbiol. 2020, 18, 607–621. [Google Scholar] [CrossRef] [PubMed]
- Bais, H.P.; Weir, T.L.; Perry, L.G.; Gilroy, S.; Vivanco, J.M. The role of root exudates in rhizosphere interactions with plants and other organisms. Annu. Rev. Plant Biol. 2006, 57, 233–266. [Google Scholar] [CrossRef] [PubMed]
- Vives-Peris, V.; de Ollas, C.; Gomez-Cadenas, A.; Perez-Clemente, R.M. Root exudates: From plant to rhizosphere and beyond. Plant Cell Rep. 2020, 39, 3–17. [Google Scholar] [CrossRef] [PubMed]
- Chaparro, J.M.; Badri, D.V.; Bakker, M.G.; Sugiyama, A.; Manter, D.K.; Vivanco, J.M. Root exudation of phytochemicals in Arabidopsis follows specific patterns that are developmentally programmed and correlate with soil microbial functions. PLoS ONE 2013, 8, e55731. [Google Scholar] [CrossRef]
- Haldar, S.; Sengupta, S. Plant-microbe Cross-talk in the Rhizosphere: Insight and Biotechnological Potential. Open Microbiol. J. 2015, 9, 1–7. [Google Scholar] [CrossRef]
- Ling, N.; Raza, W.; Ma, J.H.; Huang, Q.W.; Shen, Q.R. Identification and role of organic acids in watermelon root exudates for recruiting Paenibacillus polymyxa SQR-21 in the rhizosphere. Eur. J. Soil Biol. 2011, 47, 374–379. [Google Scholar] [CrossRef]
- Tan, S.Y.; Yang, C.L.; Mei, X.L.; Shen, S.Y.; Raza, W.; Shen, Q.R.; Xu, Y.C. The effect of organic acids from tomato root exudates on rhizosphere colonization of Bacillus amyloliquefaciens T-5. Appl. Soil Ecol. 2013, 64, 15–22. [Google Scholar] [CrossRef]
- Hu, L.F.; Robert, C.A.M.; Cadot, S.; Zhang, X.; Ye, M.; Li, B.B.; Manzo, D.; Chervet, N.; Steinger, T.; van der Heijden, M.G.A.; et al. Root exudate metabolites drive plant-soil feedbacks on growth and defense by shaping the rhizosphere microbiota. Nat. Commun. 2018, 9, 2738. [Google Scholar] [CrossRef]
- Gao, T.; Wang, X.; Qin, Y.; Ren, Z.; Zhao, X. Watermelon Root Exudates Enhance Root Colonization of Bacillus amyloliquefaciens TR2. Curr. Microbiol. 2023, 80, 110. [Google Scholar] [CrossRef]
- Lin, Q.; Li, M.M.; Wang, Y.; Xu, Z.X.; Li, L. Root exudates and chemotactic strains mediate bacterial community assembly in the rhizosphere soil of Casuarina equisetifolia L. Front. Plant Sci. 2022, 13, 988442. [Google Scholar] [CrossRef]
- Feng, H.C.; Zhang, N.; Du, W.B.; Zhang, H.H.; Liu, Y.P.; Fu, R.X.; Shao, J.H.; Zhang, G.S.; Shen, Q.R.; Zhang, R.F. Identification of Chemotaxis Compounds in Root Exudates and Their Sensing Chemoreceptors in Plant-Growth-Promoting Rhizobacteria Bacillus amyloliquefaciens SQR9. Mol. Plant-Microbe Interact. 2018, 31, 995–1005. [Google Scholar] [CrossRef]
- Ankati, S.; Rani, T.S.; Podile, A.R. Changes in Root Exudates and Root Proteins in Groundnut-Pseudomonas sp. Interaction Contribute to Root Colonization by Bacteria and Defense Response of the Host. J. Plant Growth Regul. 2019, 38, 523–538. [Google Scholar] [CrossRef]
- Wang, H.W.; Ma, C.Y.; Xu, F.J.; Lu, F.; Zhang, W.; Dai, C.C. Root endophyte-enhanced peanut-rhizobia interaction is associated with regulation of root exudates. Microbiol. Res. 2021, 250, 126765. [Google Scholar] [CrossRef]
- Ankati, S.; Podile, A.R. Metabolites in the root exudates of groundnut change during interaction with plant growth promoting rhizobacteria in a strain-specific manner. J. Plant Physiol. 2019, 243, 153057. [Google Scholar] [CrossRef] [PubMed]
- Xiong, Y.W.; Li, X.W.; Wang, T.T.; Gong, Y.; Zhang, C.M.; Xing, K.; Qin, S. Root exudates-driven rhizosphere recruitment of the plant growth-promoting rhizobacterium Bacillus flexus KLBMP 4941 and its growth-promoting effect on the coastal halophyte Limonium sinense under salt stress. Ecotoxicol. Environ. Saf. 2020, 194, 110374. [Google Scholar] [CrossRef] [PubMed]
- Marchese, A.; Barbieri, R.; Coppo, E.; Orhan, I.E.; Daglia, M.; Nabavi, S.F.; Izadi, M.; Abdollahi, M.; Nabavi, S.M.; Ajami, M. Antimicrobial activity of eugenol and essential oils containing eugenol: A mechanistic viewpoint. Crit. Rev. Microbiol. 2017, 43, 668–689. [Google Scholar] [CrossRef] [PubMed]
- Vance, C.P.; Uhde-Stone, C.; Allan, D.L. Phosphorus acquisition and use: Critical adaptations by plants for securing a nonrenewable resource. New Phytol. 2003, 157, 423–447. [Google Scholar] [CrossRef] [PubMed]
- Almeida, D.S.; Delai, L.B.; Sawaya, A.; Rosolem, C.A. Exudation of organic acid anions by tropical grasses in response to low phosphorus availability. Sci. Rep. 2020, 10, 16955. [Google Scholar] [CrossRef]
- Sasse, J.; Martinoia, E.; Northen, T. Feed Your Friends: Do Plant Exudates Shape the Root Microbiome? Trends Plant Sci. 2018, 23, 25–41. [Google Scholar] [CrossRef]
- Gupta, V.; Kumar, G.N.; Buch, A. Colonization by multi-potential Pseudomonas aeruginosa P4 stimulates peanut (Arachis hypogaea L.) growth, defence physiology and root system functioning to benefit the root-rhizobacterial interface. J. Plant Physiol. 2020, 248, 153144. [Google Scholar] [CrossRef]
- Zhang, H.H.; Shen, W.S.; Ma, C.Y.; Li, S.S.; Chen, J.; Mou, X.F.; Cheng, W.W.; Lei, P.; Xu, H.; Gao, N.; et al. Simultaneous Nitrogen Removal and Plant Growth Promotion Using Salt-tolerant Denitrifying Bacteria in Agricultural Wastewater. Microbes Environ. 2022, 37, ME22025. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Z.Y.; Zhang, H.H.; Leng, J.; Niu, H.Q.; Chen, X.C.; Liu, D.; Chen, Y.; Gao, N.; Ying, H.J. Isolation and characterization of plant growth-promoting rhizobacteria and their effects on the growth ofMedicago sativaL. under salinity conditions. Antonie Van Leeuwenhoek Int. J. Gen. 2020, 113, 1263–1278. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Kronzucker, H.J.; Shi, W. Stigmasterol root exudation arising from Pseudomonas inoculation of the duckweed rhizosphere enhances nitrogen removal from polluted waters. Environ. Pollut. 2021, 287, 117587. [Google Scholar] [CrossRef] [PubMed]
- Shen, H.; Yan, X.L.; Zhao, M.; Zheng, S.L.; Wang, X.R. Exudation of organic acids in common bean as related to mobilization of aluminum- and iron-bound phosphates. Environ. Exp. Bot. 2002, 48, 1–9. [Google Scholar] [CrossRef]
- Oburger, E.; Dell’mour, M.; Hann, S.; Wieshammer, G.; Puschenreiter, M.; Wenzel, W.W. Evaluation of a novel tool for sampling root exudates from soil-grown plants compared to conventional techniques. Environ. Exp. Bot. 2013, 87, 235–247. [Google Scholar] [CrossRef]
- Liang, C.; Schimel, J.P.; Jastrow, J.D. The importance of anabolism in microbial control over soil carbon storage. Nat. Microbiol. 2017, 2, 17105. [Google Scholar] [CrossRef] [PubMed]
- Mada, R.J.; Bagyaraj, D.J. Root exudation from Leucaena leucocephala in relation to mycorrhizal colonization. World J. Microbiol. Biotechnol. 1993, 9, 342–344. [Google Scholar] [CrossRef]
- Deutscher, J. The mechanisms of carbon catabolite repression in bacteria. Curr. Opin. Microbiol. 2008, 11, 87–93. [Google Scholar] [CrossRef]
- Yuan, J.; Wu, Y.C.; Zhao, M.L.; Wen, T.; Huang, Q.W.; Shen, Q.R. Effect of phenolic acids from banana root exudates on root colonization and pathogen suppressive properties of Bacillus amyloliquefaciens NJN-6. Biol. Control 2018, 125, 131–137. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Soluble Sugar (mg g−1) | Soluble Protein (mg g−1) | Free Amino Acids (mg g−1) |
|---|---|---|---|
| RE/NRCB010− | 8.1 ± 1.5 ** | 66.5 ± 4.0 | 0.93 ± 0.06 |
| RE/NRCB010+ | 4.7 ± 0.4 | 126.4 ± 29.0 ** | 0.97 ± 0.07 |
| Treatment | Biomass (OD600) | Diameter of Swarming Area (cm) | Biofilm Formation (OD560) |
|---|---|---|---|
| Methanol control | 0.10 ± 0.01 b | 1.57 ± 0.03 c | 1.14 ± 0.08 c |
| RE/NRCB010− | 0.13 ± 0.00 ab | 1.67 ± 0.03 b | 1.92 ± 0.12 b |
| RE/NRCB010+ | 0.15 ± 0.01 a | 1.80 ± 0.00 a | 2.24 ± 0.03 a |
| No. | Retention Time | Compounds | Relative Area (%) | |
|---|---|---|---|---|
| (min) | NRCB010− | NRCB010+ | ||
| 1 | 7.973 | 12,15-Octadecadiynoic acid, methyl ester | 0.35 ** | 0.16 |
| 2 | 9.599 | Nonanal dimethyl acetal | 0.33 | 0.44 * |
| 3 | 10.425 | Octaethylene glycol monododecyl ether | 0.22 | 0.30 * |
| 4 | 10.544 | 3,4-Dimethylbenzamide | 1.96 | 3.00 * |
| 5 | 11.275 | Octadecane, 6-methyl- | 0.37 * | 0.28 |
| 6 | 12.071 | Heptadecane, 2,6,10,14-tetramethyl- | 0.75 ** | 0.46 |
| 7 | 12.738 | 2,4-Di-tert-butylphenol | 6.23 | 7.60 * |
| 8 | 17.248 | Methyl hexadecanoate | 8.50 | 13.59 ** |
| 9 | 17.503 | n-Hexadecanoic acid | 0.51 | 0.88 ** |
| 10 | 18.928 | Ethyl iso-allocholate | 0.24 | 0.31 ** |
| 11 | 19.152 | Methyl stearate | 5.24 | 11.45 * |
| No. | Metabolite | Biomass (OD600) | Diameter of Swarming Area (cm) | Biofilm Formation (OD560) |
|---|---|---|---|---|
| 1 | 12,15-Octadecadiynoic acid, methyl ester | −0.803 | −0.929 ** | −0.898 * |
| 2 | Nonanal dimethyl acetal | 0.648 | 0.931 ** | 0.857 * |
| 3 | Octaethylene glycol monododecyl ether | 0.525 | 0.643 | 0.457 |
| 4 | 3,4-Dimethylbenzamide | 0.720 | 0.934 ** | 0.963 ** |
| 5 | Octadecane, 6-methyl- | −0.660 | −0.572 | −0.492 |
| 6 | Heptadecane, 2,6,10,14-tetramethyl- | −0.741 | −0.894 * | −0.870 * |
| 7 | 2,4-Di-tert-butylphenol | 0.548 | 0.969 ** | 0.901 * |
| 8 | Methyl hexadecanoate | 0.816 * | 0.841 | 0.784 |
| 9 | n-Hexadecanoic acid | 0.678 | 0.973 ** | 0.891 * |
| 10 | Ethyl iso-allocholate | 0.812 * | 0.841 * | 0.788 |
| 11 | Methyl stearate | 0.673 | 0.969 ** | 0.965 * |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, H.; Zheng, D.; Hu, C.; Cheng, W.; Lei, P.; Xu, H.; Gao, N. Certain Tomato Root Exudates Induced by Pseudomonas stutzeri NRCB010 Enhance Its Rhizosphere Colonization Capability. Metabolites 2023, 13, 664. https://doi.org/10.3390/metabo13050664
Zhang H, Zheng D, Hu C, Cheng W, Lei P, Xu H, Gao N. Certain Tomato Root Exudates Induced by Pseudomonas stutzeri NRCB010 Enhance Its Rhizosphere Colonization Capability. Metabolites. 2023; 13(5):664. https://doi.org/10.3390/metabo13050664
Chicago/Turabian StyleZhang, Huanhuan, Donghui Zheng, Chun Hu, Wenwen Cheng, Peng Lei, Hong Xu, and Nan Gao. 2023. "Certain Tomato Root Exudates Induced by Pseudomonas stutzeri NRCB010 Enhance Its Rhizosphere Colonization Capability" Metabolites 13, no. 5: 664. https://doi.org/10.3390/metabo13050664
APA StyleZhang, H., Zheng, D., Hu, C., Cheng, W., Lei, P., Xu, H., & Gao, N. (2023). Certain Tomato Root Exudates Induced by Pseudomonas stutzeri NRCB010 Enhance Its Rhizosphere Colonization Capability. Metabolites, 13(5), 664. https://doi.org/10.3390/metabo13050664