
The Role of Plant Growth-Promoting Microorganisms (PGPMs) and Their Feasibility in Hydroponics and Vertical Farming


Abstract
1. Introduction
2. The Role of PGPMs in Growth and Productivity
3. The Role of PGPMs in Plant Detoxification
4. The Role of PGPMs in Abiotic Stress Tolerance
4.1. Tolerance to a Drought Environment
4.2. Tolerance to Extreme Temperatures
4.3. Tolerance to a Hypersaline Environment
5. Possibilities of Using PGPMs in Hydroponics and Vertical Farming
6. Conclusions
Funding
Acknowledgments
Conflicts of Interest
References
- Ma, Y. Biotechnological potential of plant-microbe interactions in environmental decontamination. Front. Plant Sci. 2019, 10, 1519. [Google Scholar]
- Dhawi, F.; Datta, R.; Ramakrishna, W. Mycorrhiza and heavy metal resistant bacteria enhance growth, nutrient uptake and alter metabolic profile of sorghum grown in marginal soil. Chemosphere 2016, 157, 33–41. [Google Scholar] [CrossRef] [PubMed]
- Dhawi, F.; Datta, R.; Ramakrishna, W. Proteomics provides insights into biological pathways altered by plant growth promoting bacteria and arbuscular mycorrhiza in sorghum grown in marginal soil. Biochim. Biophys. Acta Proteins Proteom. 2017, 1865, 243–251. [Google Scholar] [CrossRef] [PubMed]
- Dhawi, F.; Hess, A. Plant growth-prompting bacteria influenced metabolites of Zea mays var. amylacea and Pennisetum americanum p. in a species-specific manner. Adv. Biol. Chem. 2017, 7, 161. [Google Scholar] [CrossRef]
- Dhawi, F.; Hess, A. Poor-soil rhizosphere enriched with different microbial activities influence the availability of base elements. Open J. Ecol. 2017, 7, 495–502. [Google Scholar] [CrossRef]
- Dhawi, F.; Alsanie, S.I. So it is above, so it is below: Microbial Pathways Associated with Date Palm Trees. Ann. Rom. Soc. Cell Biol. 2021, 25, 2725–2740. [Google Scholar]
- Dhawi, F. Investigating Two Date Palm Cultivars Microbial Pathways. J. Hunan Univ. Nat. Sci. 2022, 49, 158–160. [Google Scholar]
- Dhawi, F.; Alsanie, S.I. Investigation of Microbial Community Structure and Diversity in the Rhizosphere of Date Palm (Phoenix dactylifera L.), Sukkari Cultivar. J. Hunan Univ. Nat. Sci. 2022, 4, 9–24. [Google Scholar]
- Mehmood, T.; Gaurav, G.K.; Cheng, L.; Klemeš, J.J.; Usman, M.; Bokhari, A.; Lu, J. A review on plant-microbial interactions, functions, mechanisms and emerging trends in bioretention system to improve multi-contaminated stormwater treatment. J. Environ. Manag. 2021, 294, 113108. [Google Scholar] [CrossRef]
- Conlon, R.; Wang, M.; Germaine, X.L.; Mali, R.; Dowling, D.; Germaine, K.J. Ecopiling: Beneficial Soil Bacteria, Plants, and Optimized Soil Conditions for Enhanced Remediation of Hydrocarbon Polluted Soil. Good Microbes Med. Food Prod. Biotechnol. Bioremediat. Agric. 2022, 337–347. [Google Scholar] [CrossRef]
- Augusta, A.C.; Bertha, E.E.; Eromosele, A.S. Plant-Microbe Interaction: Prospects and Applications in Sustainable Environmental Management. In Plant Hormones: Recent Advances. New Perspectives and Applications; IntechOpen: London, UK, 2022; p. 43. [Google Scholar]
- Schnitzler, W.H. Urban hydroponics–facts and vision. In SEAVEG 2012 High Value Vegetables in Southeast Asia: Production, Supply and Demand; AVRDC-World Vegetable Center: Tainan, Taiwan, 2013; p. 285. [Google Scholar]
- Despommier, D. Farming up the city: The rise of urban vertical farms. Trends Biotechnol. 2013, 31, 388–389. [Google Scholar]
- Kaur, G.; Chawla, P. All about Vertical Farming: A Review. Turk. J. Comput. Math. Educ. 2021, 12, 1–14. [Google Scholar]
- Kulak, M.; Graves, A.; Chatterton, J. Reducing greenhouse gas emissions with urban agriculture: A Life Cycle Assessment perspective. Landsc. Urban Plan 2013, 111, 68–78. [Google Scholar] [CrossRef]
- Koriesh, E.M.; Abo-Soud, I.H. Facing Climate Change: Urban Gardening and Sustainable Agriculture. In Climate Change Impacts on Agriculture and Food Security in Egypt; Springer: Cham, Switzerland, 2020; pp. 345–419. [Google Scholar]
- Rajan, P.; Lada, R.R.; MacDonald, M.T. Advancement in indoor vertical farming for microgreen production. Am. J. Plant Sci. 2019, 10, 1397. [Google Scholar]
- Singh, B.K.; Liu, H.; Trivedi, P. Eco-holobiont: A new concept to identify drivers of host-associated microorganisms. Environ. Microbiol. 2020, 22, 564–567. [Google Scholar]
- Zilber-Rosenberg, I.; Rosenberg, E. Role of microorganisms in the evolution of animals and plants: The hologenome theory of evolution. FEMS Microbiol. Rev. 2008, 32, 723–735. [Google Scholar]
- Kothe, E.; Turnau, K. Mycorrhizosphere communication: Mycorrhizal fungi and endophytic fungus-plant interactions. Front. Microbiol. 2018, 9, 3015. [Google Scholar]
- de la Fuente Cantó, C.; Simonin, M.; King, E.; Moulin, L.; Bennett, M.J.; Castrillo, G.; Laplaze, L. An extended root phenotype: The rhizosphere, its formation and impacts on plant fitness. Plant J. 2020, 103, 951–964. [Google Scholar]
- Dhawi, F. Plant growth promoting Rhizobacteria (PGPR) regulated Phyto and microbial beneficial protein interactions. Open Life Sci. 2020, 15, 68–78. [Google Scholar] [CrossRef]
- Ghitti, E.; Rolli, E.; Crotti, E.; Borin, S. Flavonoids Are Intra-and Inter-Kingdom Modulator Signals. Microorganisms 2022, 10, 2479. [Google Scholar]
- Pratush, A.; Kumar, A.; Hu, Z. Adverse effect of heavy metals (As, Pb, Hg, and Cr) on health and their bioremediation strategies: A review. Int. Microbiol. 2018, 21, 97–106. [Google Scholar] [CrossRef]
- Ma, Y.; Dias, M.C.; Freitas, H. Drought and salinity stress responses and microbe-induced tolerance in plants. Front. Plant Sci. 2020, 11, 591911. [Google Scholar] [CrossRef]
- Hacquard, S.; Garrido-Oter, R.; González, A.; Spaepen, S.; Ackermann, G.; Lebeis, S.; McHardy, A.C.; Dangl, J.L.; Knight, R.; Ley, R.; et al. Microbiota and host nutrition across plant and animal kingdoms. Cell Host Microbe 2015, 17, 603–616. [Google Scholar] [CrossRef]
- Ojuederie, O.B.; Babalola, O.O. Microbial and plant-assisted bioremediation of heavy metal polluted environments: A review. Int. J. Environ. Res. Public Health 2017, 14, 1504. [Google Scholar] [CrossRef] [PubMed]
- Baset, M.M.; Shamsuddin, Z.H.; Wahab, Z.; Marziah, M. Effect of plant growth promoting rhizobacterial (PGPR) inoculation on growth and nitrogen incorporation of tissue-cultured'musa'plantlets under nitrogen-free hydroponics condition. Aust. J. Crop Sci. 2010, 4, 85–90. [Google Scholar]
- Goswami, M.; Chakraborty, P.; Mukherjee, K.; Mitra, G.; Bhattacharyya, P.; Dey, S.; Tribedi, P. Bioaugmentation and biostimulation: A potential strategy for environmental remediation. J. Microbiol. Exp. 2018, 6, 223–231. [Google Scholar]
- Delaporte-Quintana, P.; Lovaisa, N.C.; Rapisarda, V.A.; Pedraza, R.O. The plant growth promoting bacteria Gluconacetobacter diazotrophicus and Azospirillum brasilense contribute to the iron nutrition of strawberry plants through siderophores production. Plant Growth Regul. 2020, 91, 185–199. [Google Scholar] [CrossRef]
- Phieler, R.; Merten, D.; Roth, M.; Büchel, G.; Kothe, E. Phytoremediation using microbially mediated metal accumulation in Sorghum bicolor. Environ. Sci. Pollut. Res. 2015, 22, 19408–19416. [Google Scholar] [CrossRef]
- Basu, S.; Rabara, R.C.; Negi, S.; Shukla, P. Engineering PGPMOs through gene editing and systems biology: A solution for phytoremediation? Trends Biotechnol. 2018, 36, 499–510. [Google Scholar] [PubMed]
- Abhilash, P.C.; Powell, J.R.; Singh, H.B.; Singh, B.K. Plant–microbe interactions: Novel applications for exploitation in multipurpose remediation technologies. Trends Biotechnol. 2012, 30, 416–420. [Google Scholar] [PubMed]
- Kuiper, I.; Lagendijk, E.L.; Bloemberg, G.V.; Lugtenberg, B.J. Rhizoremediation: A beneficial plant-microbe interaction. Mol. Plant-Microbe Interact. 2004, 17, 6–15. [Google Scholar] [CrossRef] [PubMed]
- Mendis, H.C.; Thomas, V.P.; Schwientek, P.; Salamzade, R.; Chien, J.T.; Waidyarathne, P.; Kloepper, J.; De La Fuente, L. Strain-specific quantification of root colonization by plant growth promoting rhizobacteria Bacillus firmus I-1582 and Bacillus amyloliquefaciens QST713 in non-sterile soil and field conditions. PLoS ONE 2018, 13, e0193119. [Google Scholar] [CrossRef] [PubMed]
- Jain, S.; Arnepalli, D.N. Biominerlisation as a remediation technique: A critical review. In Geotechnical Characterisation and Geoenvironmental Engineering; Springer: Singapore, 2019; pp. 155–162. [Google Scholar]
- Chibuike, G.U.; Obiora, S.C. Heavy metal polluted soils: Effect on plants and bioremediation methods. Appl. Environ. Soil Sci. 2014, 2014, 752708. [Google Scholar]
- Lacalle, R.G.; Aparicio, J.D.; Artetxe, U.; Urionabarrenetxea, E.; Polti, M.A.; Soto, M.; Garbisu, C.; Becerril, J.M. Gentle remediation options for soil with mixed chromium (VI) and lindane pollution: Biostimulation, bioaugmentation, phytoremediation and vermiremediation. Heliyon 2020, 6, e04550. [Google Scholar] [CrossRef] [PubMed]
- Monti, M.R.; Smania, A.M.; Fabro, G.; Alvarez, M.E.; Argarana, C.E. Engineering Pseudomonas fluorescens for biodegradation of 2,4-dinitrotoluene. Appl. Environ. Microbiol. 2005, 71, 8864–8872. [Google Scholar]
- Ren, X.; Zeng, G.; Tang, L.; Wang, J.; Wan, J.; Wang, J.; Deng, Y.; Liu, Y.; Peng, B. The potential impact on the biodegradation of organic pollutants from composting technology for soil remediation. Waste Manag. 2018, 72, 138–149. [Google Scholar]
- Kumar, A.; Chaturvedi, A.K.; Yadav, K.; Arunkumar, K.P.; Malyan, S.K.; Raja, P.; Kumar, R.; Khan, S.A.; Yadav, K.K.; Rana, K.L.; et al. Fungal phytoremediation of heavy metal-contaminated resources: Current scenario and future prospects. In Recent Advancement in White Biotechnology through Fungi; Springer: Cham, Switzerland, 2019; pp. 437–461. [Google Scholar]
- Bhantana, P.; Rana, M.S.; Sun, X.C.; Moussa, M.G.; Saleem, M.H.; Syaifudin, M.; Shah, A.; Poudel, A.; Pun, A.B.; Bhat, M.A.; et al. Arbuscular mycorrhizal fungi and its major role in plant growth, zinc nutrition, phosphorous regulation and phytoremediation. Symbiosis 2021, 84, 19–37. [Google Scholar]
- Sarwar, N.; Imran, M.; Shaheen, M.R.; Ishaque, W.; Kamran, M.A.; Matloob, A.; Rehim, A.; Hussain, S. Phytoremediation strategies for soils contaminated with heavy metals: Modifications and future perspectives. Chemosphere 2017, 171, 710–721. [Google Scholar]
- Göhre, V.; Paszkowski, U. Contribution of the arbuscular mycorrhizal symbiosis to heavy metal phytoremediation. Planta 2006, 223, 1115–1122. [Google Scholar] [CrossRef]
- Dhawi, F.; Datta, R.; Ramakrishna, W. Metabolomics, biomass and lignocellulosic total sugars analysis in foxtail millet (Setaria italica) inoculated with different combinations of plant growth promoting bacteria and mycorrhiza. Commun. Plant Sci. 2018, 8, 8–14. [Google Scholar]
- Vamerali, T.; Bandiera, M.; Mosca, G. Field crops for phytoremediation of metal-contaminated land. A review. Environ. Chem. Lett. 2010, 8, 1–17. [Google Scholar]
- Chen, B.D.; Li, X.L.; Tao, H.Q.; Christie, P.; Wong, M.H. The role of arbuscular mycorrhiza in zinc uptake by red clover growing in a calcareous soil spiked with various quantities of zinc. Chemosphere 2003, 50, 839–846. [Google Scholar] [CrossRef] [PubMed]
- Joner, E.J.; Leyval, C. Phytoremediation of organic pollutants using mycorrhizal plants: A new aspect of rhizosphere interactions. In Sustainable Agriculture; Springer: Dordrecht, The Netherlands, 2009; pp. 885–894. [Google Scholar]
- Arantza, S.J.; Hiram, M.R.; Erika, K.; Chávez-Avilés, M.N.; Valiente-Banuet, J.I.; Fierros-Romero, G. Bio- and phytoremediation: Plants and microbes to the rescue of heavy metal polluted soils. SN Appl. Sci. 2022, 4, 59. [Google Scholar] [CrossRef]
- Becerra-Castro, C.; Prieto-Fernández, A.; Álvarez-Lopez, V.; Monterroso, C.; Cabello-Conejo, M.I.; Acea, M.J.; Kidd, P.S. Nickel solubilizing capacity and characterization of rhizobacteria isolated from hyperaccumulating and non-hyperaccumulating subspecies of Alyssum serpyllifolium. Int. J. Phytoremediat. 2011, 13, 229–244. [Google Scholar] [PubMed]
- Gill, R.A.; Ahmar, S.; Ali, B.; Saleem, M.H.; Khan, M.U.; Zhou, W.; Liu, S. The role of membrane transporters in plant growth and development, and abiotic stress tolerance. Int. J. Mol. Sci. 2021, 22, 12792. [Google Scholar] [CrossRef] [PubMed]
- Malamy, J.E. Intrinsic and environmental response pathways that regulate root system architecture. Plant Cell Environ. 2005, 28, 67–77. [Google Scholar]
- Doornbos, R.F.; Geraats, B.P.; Kuramae, E.E.; Van Loon, L.C.; Bakker, P.A. Effects of jasmonic acid, ethylene, and salicylic acid signaling on the rhizosphere bacterial community of Arabidopsis thaliana. Mol. Plant-Microbe Interact. 2011, 24, 395–407. [Google Scholar] [PubMed]
- Osmont, K.S.; Sibout, R.; Hardtke, C.S. Hidden branches: Developments in root system architecture. Annu. Rev. Plant Biol. 2007, 58, 93–113. [Google Scholar] [CrossRef]
- Verbon, E.H.; Liberman, L.M. Beneficial microbes affect endogenous mechanisms controlling root development. Trends Plant science 2016, 21, 218–229. [Google Scholar] [CrossRef]
- Braud, A.; Jézéquel, K.; Bazot, S.; Lebeau, T. Enhanced phytoextraction of an agricultural Cr-and Pb-contaminated soil by bioaugmentation with siderophore-producing bacteria. Chemosphere 2009, 74, 280–286. [Google Scholar] [CrossRef]
- Chang, P.; Gerhardt, K.E.; Huang, X.D.; Yu, X.M.; Glick, B.R.; Gerwing, P.D.; Greenberg, B.M. Plant growth-promoting bacteria facilitate the growth of barley and oats in salt-impacted soil: Implications for phytoremediation of saline soils. Int. J. Phytoremediation 2014, 16, 1133–1147. [Google Scholar] [CrossRef] [PubMed]
- Babu, A.G.; Kim, J.D.; Oh, B.T. Enhancement of heavy metal phytoremediation by Alnus firma with endophytic Bacillus thuringiensis GDB-1. J. Hazard. Mater. 2013, 250, 477–483. [Google Scholar] [CrossRef] [PubMed]
- Agnello, A.C.; Bagard, M.; van Hullebusch, E.D.; Esposito, G.; Huguenot, D. Comparative bioremediation of heavy metals and petroleum hydrocarbons co-contaminated soil by natural attenuation, phytoremediation, bioaugmentation and bioaugmentation-assisted phytoremediation. Sci. Total Environ. 2016, 563, 693–703. [Google Scholar] [CrossRef] [PubMed]
- Dong, R.; Gu, L.; Guo, C.; Xun, F.; Liu, J. Effect of PGPR Serratia marcescens BC-3 and AMF Glomus intraradices on phytoremediation of petroleum contaminated soil. Ecotoxicology 2014, 23, 674–680. [Google Scholar] [CrossRef]
- DalCorso, G.; Fasani, E.; Manara, A.; Visioli, G.; Furini, A. Heavy metal pollutions: State of the art and innovation in phytoremediation. Int. J. Mol. Sci. 2019, 20, 3412. [Google Scholar] [CrossRef]
- Ma, Y.; Oliveira, R.S.; Freitas, H.; Zhang, C. Biochemical and molecular mechanisms of plant-microbe-metal interactions: Relevance for phytoremediation. Front. Plant Sci. 2016, 7, 918. [Google Scholar]
- Nasfi, Z.; Busch, H.; Kehraus, S.; Linares-Otoya, L.; König, G.M.; Schäberle, T.F.; Bachoual, R. Soil bacteria isolated from tunisian arid areas show promising antimicrobial activities against gram-negatives. Front. Microbiol. 2018, 9, 2742. [Google Scholar]
- Eida, A.A.; Ziegler, M.; Lafi, F.F.; Michell, C.T.; Voolstra, C.R.; Hirt, H.; Saad, M.M. Desert plant bacteria reveal host influence and beneficial plant growth properties. PLoS ONE 2018, 13, e0208223. [Google Scholar]
- He, M.; He, C.Q.; Ding, N.Z. Abiotic stresses: General defenses of land plants and chances for engineering multistress tolerance. Front. Plant Sci. 2018, 9, 1771. [Google Scholar]
- Saleem, M.; Arshad, M.; Hussain, S.; Bhatti, A.S. Perspective of plant growth promoting rhizobacteria (PGPR) containing ACC deaminase in stress agriculture. J. Ind. Microbiol. Biotechnol. 2007, 34, 635–648. [Google Scholar]
- Glick, B.R.; Penrose, D.M.; Li, J. A model for the lowering of plant ethylene concentrations by plant growth-promoting bacteria. J. Theor. Biol. 1998, 190, 63–68. [Google Scholar] [PubMed]
- Rai, A.; Borpatragohain, B.; Sahoo, S. Role of plant-microbe interactions on abiotic stress tolerance in plants: A review. Int. J. Agric. Plant Sci. 2019, 1, 25–31. [Google Scholar]
- Ait Barka, E.; Nowak, J.; Clément, C. Enhancement of chilling resistance of inoculated grapevine plantlets with a plant growth-promoting rhizobacterium, Burkholderia phytofirmans strain PsJN. Appl. Environ. Microbiol. 2006, 72, 7246–7252. [Google Scholar] [PubMed]
- Lesk, C.; Rowhani, P.; Ramankutty, N. Influence of extreme weather disasters on global crop production. Nature 2016, 529, 84–87. [Google Scholar] [CrossRef] [PubMed]
- Grover, M.; Farrugia, G.; Lurken, M.S.; Bernard, C.E.; Faussone–Pellegrini, M.S.; Smyrk, T.C.; Parkman, H.P.; Abell, T.L.; Snape, W.J.; Hasler, W.L.; et al. Cellular changes in diabetic and idiopathic gastroparesis. Gastroenterology 2011, 140, 1575–1585. [Google Scholar] [CrossRef]
- Comas, L.H.; Becker, S.R.; Cruz, V.M.; Byrne, P.F.; Dierig, D.A. Root traits contributing to plant productivity under drought. Front. Plant Sci. 2013, 4, 442. [Google Scholar] [CrossRef]
- Seleiman, M.F.; Al-Suhaibani, N.; Ali, N.; Akmal, M.; Alotaibi, M.; Refay, Y.; Dindaroglu, T.; Abdul-Wajid, H.H.; Battaglia, M.L. Drought stress impacts on plants and different approaches to alleviate its adverse effects. Plants 2021, 10, 259. [Google Scholar] [CrossRef]
- Vardharajula, S.; Zulfikar Ali, S.; Grover, M.; Reddy, G.; Bandi, V. Drought-tolerant plant growth promoting Bacillus spp.: Effect on growth, osmolytes, and antioxidant status of maize under drought stress. J. Plant Interact. 2011, 6, 1–4. [Google Scholar] [CrossRef]
- Abobatta, W.F. Drought adaptive mechanisms of plants—A review. Adv. Agric. Environ. Sci. 2019, 2, 62–65. [Google Scholar]
- Redman, R.S.; Sheehan, K.B.; Stout, R.G.; Rodriguez, R.J.; Henson, J.M. Thermotolerance generated by plant/fungal symbiosis. Science 2002, 298, 1581. [Google Scholar]
- Bui, L.; Luo, H.; Gunther, W.R.; Román-Leshkov, Y. Domino reaction catalyzed by zeolites with Brønsted and Lewis acid sites for the production of γ-valerolactone from furfural. Angew. Chem. 2013, 125, 8180–8183. [Google Scholar]
- Bashan, Y.; de-Bashan, L.E.; Prabhu, S.R.; Hernandez, J.P. Advances in plant growth-promoting bacterial inoculant technology: Formulations and practical perspectives (1998–2013). Plant Soil 2014, 378, 1–33. [Google Scholar] [CrossRef]
- Wu, J. Advances in K-Means Clustering: A Data Mining Thinking; Springer Science & Business Media: Berlin, Germany, 2012. [Google Scholar]
- Egamberdieva, D.; Wirth, S.J.; Alqarawi, A.A.; Abd_Allah, E.F.; Hashem, A. Phytohormones and beneficial microbes: Essential components for plants to balance stress and fitness. Front. Microbiol. 2017, 8, 2104. [Google Scholar]
- Grover, P.; Sahai, A. Shannon meets Tesla: Wireless information and power transfer. In Proceedings of the 2010 IEEE International Symposium on Information Theory, Austin, TX, USA, 13–18 June 2010; pp. 2363–2367. [Google Scholar]
- Sen, S.; Chandrasekhar, C.N. Effect of PGPR on growth promotion of rice (Oryza sativa L.) under salt stress. Asian J. Plant Sci. Res. 2014, 4, 62–67. [Google Scholar]
- Stegelmeier, A.A.; Rose, D.M.; Joris, B.R.; Glick, B.R. The Use of PGPB to Promote Plant Hydroponic Growth. Plants 2022, 11, 2783. [Google Scholar] [CrossRef] [PubMed]
- Barker, K.R.; Koenning, S.R. Developing sustainable systems for nematode management. Annu. Rev. Phytopathol. 1998, 36, 165–205. [Google Scholar]
- Zapata-Vahos, I.C.; Rojas-Rodas, F.; David, D.; Gutierrez-Monsalve, J.A.; Castro-Restrepo, D. Comparison of antioxidant contents of green and red leaf lettuce cultivated in hydroponic systems in greenhouses and conventional soil cultivation. Rev. Fac. Nac. Agron. Medellín 2020, 73, 9077–9088. [Google Scholar] [CrossRef]
- Nair, A.G.; Chacko, A.; Mohan, G.; Francis, T.K. Smart vertical farming using hydroponics. J. Electr. Electron. Eng. 2015, 1, 14–17. [Google Scholar]
- Panno, S.; Davino, S.; Caruso, A.G.; Bertacca, S.; Crnogorac, A.; Mandi´c, A.; Noris, E.; Mati´c, S. A review of the most common and economically important diseases that undermine the cultivation of tomato crop in the mediterranean basin. Agronomy 2021, 11, 2188. [Google Scholar] [CrossRef]
- O’Callaghan, M. Microbial Inoculation of Seed for Improved Crop Performance: Issues and Opportunities. Appl. Microbiol. Biotechnol. 2016, 100, 5729–5746. [Google Scholar] [CrossRef]
- John, C.J.; Kumar, S.; Ge, M. Probiotic Prospects of PGPR for Green and Sustainable Agriculture. Arch. Phytopathol. Plant Prot. 2020, 53, 899–914. [Google Scholar] [CrossRef]
- Danish, S.; Zafar-ul-Hye, M. Co-Application of ACC-Deaminase Producing PGPR and Timber-Waste Biochar 1034 Improves Pigments Formation, Growth and Yield of Wheat under Drought Stress. Sci. Rep. 2019, 9, 1–13. [Google Scholar]
- Korir, H.; Mungai, N.W.; Thuita, M.; Hamba, Y.; Masso, C. Co-Inoculation Effect of Rhizobia and Plant Growth Promoting Rhizobacteria on Common Bean Growth in a Low Phosphorus Soil. Front. Plant Sci. 2017, 8, 141. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, M.; Zahir, Z.A.; Asghar, H.N.; Arshad, M. The combined application of rhizobial strains and plant growth promoting rhizobacteria improves growth and productivity of mung bean (Vigna radiata L.) under salt-stressed conditions. Ann. Microbiol. 2012, 62, 1321–1330. [Google Scholar]
- Htwe, A.Z.; Moh, S.M.; Soe, K.M.; Moe, K.; Yamakawa, T. Effects of biofertilizer produced from Bradyrhizobium and Streptomyces griseoflavus on plant growth, nodulation, nitrogen fixation, nutrient uptake, and seed yield of mung bean, cowpea, and soybean. Agronomy 2019, 9, 77. [Google Scholar] [CrossRef]
- Wiggins, Z.; Akaeze, O.; Nandwani, D.; Witcher, A. Substrate properties and fertilizer rates on yield responses of lettuce in a vertical growth system. Sustainability 2020, 12, 6465. [Google Scholar] [CrossRef]
- Vejan, P.; Abdullah, R.; Khadiran, T.; Ismail, S.; Nasrulhaq Boyce, A. Role of plant growth promoting rhizobacteria in agricultural sustainability—A review. Molecules 2016, 21, 573. [Google Scholar] [CrossRef] [PubMed]
- Moncada, A.; Vetrano, F.; Miceli, A. Alleviation of salt stress by plant growth-promoting bacteria in hydroponic leaf lettuce. Agronomy 2020, 10, 1523. [Google Scholar]
- Settanni, L.; Miceli, A.; Francesca, N.; Cruciata, M.; Moschetti, G. Microbiological Investigation of Raphanus sativus L. Grown Hydroponically in Nutrient Solutions Contaminated with Spoilage and Pathogenic Bacteria. Int. J. Food Microbiol. 2013, 160, 344–352. [Google Scholar]
- Stouvenakers, G.; Dapprich, P.; Massart, S.; Jijakli, M.H. Plant Pathogens and Control Strategies in Aquaponics. In Aquaponics Food Production Systems; Springer: Cham, Switzerland, 2019; p. 353. [Google Scholar]
- Bjarnsholt, T.; Van Gennip, M.; Jakobsen, T.H.; Christensen, L.D.; Jensen, P.Ø.; Givskov, M. In Vitro Screens for Quorum Sensing Inhibitors and in vivo Confirmation of Their Effect. Nat. Protoc. 2010, 5, 282–293. [Google Scholar] [CrossRef]
- Defoirdt, T. Quorum-Sensing Systems as Targets for Antivirulence Therapy. Trends Microbiol. 2018, 26, 313–328. [Google Scholar] [CrossRef] [PubMed]
- Ye, T.; Zhou, T.; Li, Q.; Xu, X.; Fan, X.; Zhang, L.; Chen, S. Cupriavidus sp. HN-2, a Novel Quorum Quenching Bacterial Isolate, Is a Potent Biocontrol Agent against Xanthomonas campestris pv. campestris. Microorganisms 2020, 8, 45. [Google Scholar]
- Sambo, P.; Nicoletto, C.; Giro, A.; Pii, Y.; Valentinuzzi, F.; Mimmo, T.; Lugli, P.; Orzes, G.; Mazzetto, F.; Astolfi, S. Hydroponic Solutions for Soilless Production Systems: Issues and Opportunities in a Smart Agriculture Perspective. Front. Plant Sci. 2019, 10, 923. [Google Scholar]
- Gravel, V.; Martinez, C.; Antoun, H.; Tweddell, R.J. Control of greenhouse tomato root rot [Pythium ultimum] in hydroponic systems, using plant-growth-promoting microorganisms. Can. J. Plant Pathol. 2006, 28, 475–483. [Google Scholar] [CrossRef]
- Paradiso, R.; Arena, C.; De Micco, V.; Giordano, M.; Aronne, G.; De Pascale, S. Changes in leaf anatomical traits enhanced photosynthetic activity of soybean grown in hydroponics with plant growth-promoting microorganisms. Front. Plant Sci. 2017, 8, 674. [Google Scholar] [PubMed]
- Varia, J.; Kamaleson, C.; Lerer, L. Biostimulation with phycocyanin-rich Spirulina extract in hydroponic vertical farming. Sci. Hortic. 2022, 299, 111042. [Google Scholar]


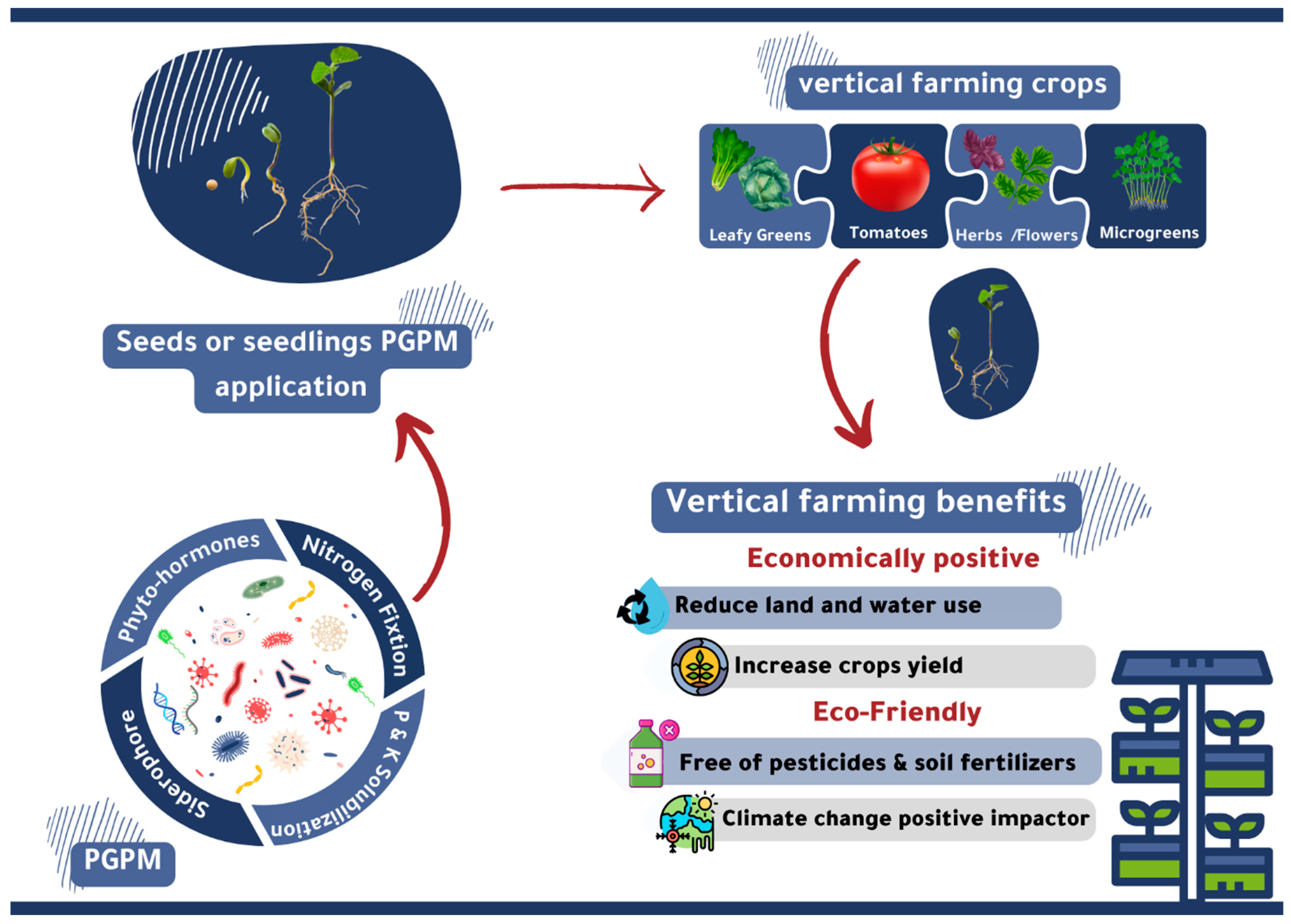{kind=link}
| Plant | Mode of PGPM Application | Type of PGPM | Influence | References |
|---|---|---|---|---|
| Banana Berangan’ (Musa spp. dessert type) | Seedling inoculation with microbial suspension | Bacillus sphaericus and azospirillum | Increase in root formation, leaf area, chlorophyll content, and consequently, total biomass | [28] |
| Strawberries | Siderophores added to hydroponic medium | Gluconacetobacter diazotrophicus and azospirillum brasilense | Increased the nutrition of iron | [30] |
| Triticum aestivum | Seedling inoculation with microbial suspension | Calothrix sp., anabaena cylindrica, chryseobacterium balustinum, pseudomonas simiae, and pseudomonas fluorescen | Increased the growth, plant height, dry shoot mass, total nutrients, and the ability to produce indole acetic acid | [31] |
| Lettuce | Biostimulant extract added to hydroponic medium | Bacillus spp. | Minimized salt stress | [96] |
| Tomato | Seedling inoculation with microbial suspension | Penicillium brevicompactum, penicillium solitum strain 1, pseudomonas fluorescens subgroup g strain 2, pseudomonas marginalis, pseudomonas putida subgroup b strain 1, pseudomonas syringae strain 1, and trichoderma atroviride | Plant growth and development in the absence of pathogens (antagonistic activity against Pythium ultimum) | [103] |
| Glycine max (L.) Merr. | Seedling inoculation with microbial suspension | Bacteria, yeasts, mycorrhiza, and Trichoderma | Higher density of smaller stomata, thicker palisade parenchyma, larger intercellular spaces in the mesophyll; increased photosynthetic traits, growth and seed production | [104] |
| Lettuce (Salanova® Lactuca sativa and Salanova® Red Crisp). | Bio-stimulant extract added to hydroponic medium | Phycocyanin-rich spirulina extract | Reduced time from seed to harvest by 6 days, increased yield by 12.5%, and improved antioxidant flavonoid levels | [105] |
| Hydroponic Types | Definition | Suggested Mode of PGPM Application |
|---|---|---|
| Wicking System | Small-scale production plants (small plants, e.g., herbs and leafy greens) | Pre-treatment seedling inoculation with microbial suspension |
| Ebb and Flow (Flood and Drain) | Seedling cultivation in commercial settings | Pre-treatment seedling inoculation with microbial suspension |
| Drip Irrigation | Commercial production for larger fruiting crops such as tomatoes, cucumbers, peppers, and strawberries | Synchronize treatment: Bio-stimulant extract added to hydroponic medium |
| Nutrient Film Technique | In commercial use for smaller leafy crops such as lettuce | Pre-treatment seedling inoculation with microbial suspension |
| Deep Water Culture | In commercial settings deep water culture systems are used for small leafy plants such as lettuce or herbs | Pre-treatment seedling inoculation with microbial suspension |
| Aeroponics | Commercial growers to produce small leafy plants and potato mini-tubers | Pre-treatment seedling inoculation with microbial suspension |
| Aquaponics | Growing fish and plants in the same system | Pre-treatment seedling inoculation with microbial suspension |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dhawi, F. The Role of Plant Growth-Promoting Microorganisms (PGPMs) and Their Feasibility in Hydroponics and Vertical Farming. Metabolites 2023, 13, 247. https://doi.org/10.3390/metabo13020247
Dhawi F. The Role of Plant Growth-Promoting Microorganisms (PGPMs) and Their Feasibility in Hydroponics and Vertical Farming. Metabolites. 2023; 13(2):247. https://doi.org/10.3390/metabo13020247
Chicago/Turabian StyleDhawi, Faten. 2023. "The Role of Plant Growth-Promoting Microorganisms (PGPMs) and Their Feasibility in Hydroponics and Vertical Farming" Metabolites 13, no. 2: 247. https://doi.org/10.3390/metabo13020247
APA StyleDhawi, F. (2023). The Role of Plant Growth-Promoting Microorganisms (PGPMs) and Their Feasibility in Hydroponics and Vertical Farming. Metabolites, 13(2), 247. https://doi.org/10.3390/metabo13020247