
Identification of Antimicrobial Metabolites from the Egyptian Soil-Derived Amycolatopsis keratiniphila Revealed by Untargeted Metabolomics and Molecular Docking

 ,
,  ,
,  , ,
, ,  , and
, and
Abstract
1. Introduction
2. Experimental
2.1. Isolation of A. keratiniphila DPA04
2.2. 16S rRNA Gene Sequencing and Phylogenic Analysis of A. keratiniphila DPA04 Strain
2.3. Preparation of Crude Extracts
2.4. Antibacterial Activity
2.5. Determination of the Minimal Inhibitory Concentration (MIC)
2.6. Liquid Chromatography with Tandem Mass Spectrometry (LC-MS/MS) Analysis
2.7. MS/MS Data Preprocessing
2.8. Feature-Based Molecular Networking
2.9. Molecular Formula Prediction and Metabolites Identification
2.10. Molecular Docking Studies
3. Results
3.1. Taxonomic Identification of A. keratiniphila DPA04
3.2. Antibacterial Activity
3.3. Chemical Analysis
3.4. Docking Studies
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hutchings, M.I.; Truman, A.W.; Wilkinson, B. Antibiotics: Past, present and future. Curr. Opin. Microbiol. 2019, 51, 72–80. [Google Scholar] [CrossRef] [PubMed]
- Bhattarai, B.R.; Khadayat, K.; Aryal, N.; Aryal, B.; Lamichhane, U.; Bhattarai, K.; Rana, N.; Regmi, B.P.; Adhikari, A.; Thapa, S. Untargeted Metabolomics of S treptomyces Species Isolated from Soils of Nepal. Processes 2022, 10, 1173. [Google Scholar] [CrossRef]
- Lima, R.; Del Fiol, F.S.; Balcão, V.M. Prospects for the use of new technologies to combat multidrug-resistant bacteria. Front. Pharmacol. 2019, 10, 692. [Google Scholar] [CrossRef]
- Tavares, L.S.; Silva, C.S.; de Souza, V.C.; da Silva, V.L.; Diniz, C.G.; Santos, M.O. Strategies and molecular tools to fight antimicrobial resistance: Resistome, transcriptome, and antimicrobial peptides. Front. Microbiol. 2013, 4, 412. [Google Scholar] [CrossRef]
- World Health Organization. Antimicrobial Resistance. 2019. Available online: https://www.who.int/news-room/fact-sheets/detail/antimicrobial-resistance (accessed on 15 December 2022).
- Genilloud, O. Actinomycetes: Still a source of novel antibiotics. Nat. Prod. Rep. 2017, 34, 1203–1232. [Google Scholar] [CrossRef]
- Mohamed, O.G.; Khalil, Z.G.; Salim, A.A.; Cui, H.; Blumenthal, A.; Capon, R.J. Lincolnenins A–D: Isomeric bactericidal bianthracenes from Streptomyces lincolnensis. J. Org. Chem. 2020, 86, 11011–11018. [Google Scholar] [CrossRef] [PubMed]
- Mohamed, O.G.; Dorandish, S.; Lindow, R.; Steltz, M.; Shoukat, I.; Shoukat, M.; Chehade, H.; Baghdadi, S.; McAlister-Raeburn, M.; Kamal, A. Identification of a New Antimicrobial, Desertomycin H, Utilizing a Modified Crowded Plate Technique. Mar. Drugs 2021, 19, 424. [Google Scholar] [CrossRef]
- Tiwari, K.; Gupta, R.K. Rare actinomycetes: A potential storehouse for novel antibiotics. Crit. Rev. Biotechnol. 2012, 32, 108–132. [Google Scholar] [CrossRef]
- Song, Z.; Xu, T.; Wang, J.; Hou, Y.; Liu, C.; Liu, S.; Wu, S. Secondary metabolites of the genus Amycolatopsis: Structures, bioactivities and biosynthesis. Molecules 2021, 26, 1884. [Google Scholar] [CrossRef] [PubMed]
- Rateb, M.E.; Houssen, W.E.; Harrison, W.T.; Deng, H.; Okoro, C.K.; Asenjo, J.A.; Andrews, B.A.; Bull, A.T.; Goodfellow, M.; Ebel, R. Diverse metabolic profiles of a Streptomyces strain isolated from a hyper-arid environment. J. Nat. Prod. 2011, 74, 1965–1971. [Google Scholar] [CrossRef]
- Pan, R.; Bai, X.; Chen, J.; Zhang, H.; Wang, H. Exploring structural diversity of microbe secondary metabolites using OSMAC strategy: A literature review. Front. Microbiol. 2019, 10, 294. [Google Scholar] [CrossRef]
- Tangerina, M.M.; Furtado, L.C.; Leite, V.M.; Bauermeister, A.; Velasco-Alzate, K.; Jimenez, P.C.; Garrido, L.M.; Padilla, G.; Lopes, N.P.; Costa-Lotufo, L.V. Metabolomic study of marine Streptomyces sp.: Secondary metabolites and the production of potential anticancer compounds. PLoS One 2020, 15, e0244385. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Carver, J.J.; Phelan, V.V.; Sanchez, L.M.; Garg, N.; Peng, Y.; Nguyen, D.D.; Watrous, J.; Kapono, C.A.; Luzzatto-Knaan, T. Sharing and community curation of mass spectrometry data with Global Natural Products Social Molecular Networking. Nat. Biotechnol. 2016, 34, 828–837. [Google Scholar] [CrossRef] [PubMed]
- Hussein, M.E.; Mohamed, O.G.; El-Fishawy, A.M.; El-Askary, H.I.; Hamed, A.A.; Abdel-Aziz, M.M.; Alnajjar, R.; Belal, A.; Naglah, A.M.; Almehizia, A.A.; et al. Anticholinesterase Activity of Budmunchiamine Alkaloids Revealed by Comparative Chemical Profiling of Two Albizia spp., Molecular Docking and Dynamic Studies. Plants 2022, 11, 3286. [Google Scholar] [CrossRef]
- Nothias, L.-F.; Petras, D.; Schmid, R.; Dührkop, K.; Rainer, J.; Sarvepalli, A.; Protsyuk, I.; Ernst, M.; Tsugawa, H.; Fleischauer, M. Feature-based molecular networking in the GNPS analysis environment. Nat. Methods 2020, 17, 905–908. [Google Scholar] [CrossRef] [PubMed]
- Mohamed, H.; Hassane, A.; Rawway, M.; El-Sayed, M.; Gomaa, A.E.-R.; Abdul-Raouf, U.; Shah, A.M.; Abdelmotaal, H.; Song, Y. Antibacterial and cytotoxic potency of thermophilic Streptomyces werraensis MI-S. 24-3 isolated from an Egyptian extreme environment. Arch. Microbiol. 2021, 203, 4961–4972. [Google Scholar] [CrossRef]
- Elsayed, T.R.; Galil, D.F.; Sedik, M.Z.; Hassan, H.; Sadik, M.W. Antimicrobial and anticancer activities of actinomycetes isolated from Egyptian soils. Int. J. Curr. Microbiol. Appl. Sci 2020, 9, 2020. [Google Scholar] [CrossRef]
- Weisburg, W.G.; Barns, S.M.; Pelletier, D.A.; Lane, D.J. 16S ribosomal DNA amplification for phylogenetic study. J. Bacteriol. 1991, 173, 697–703. [Google Scholar] [CrossRef]
- Heuer, H.; Wieland, G.; Schönfeld, J.; Schönwälder, A.; Gomes, N.; Smalla, K. Bacterial community profiling using DGGE or TGGE analysis. Environ. Mol. Microbiol. Protoc. Appl. 2001, 9, 177–190. [Google Scholar]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar]
- Larkin, M.A.; Blackshields, G.; Brown, N.P.; Chenna, R.; McGettigan, P.A.; McWilliam, H.; Valentin, F.; Wallace, I.M.; Wilm, A.; Lopez, R. Clustal W and Clustal X version 2.0. Bioinformatics 2007, 23, 2947–2948. [Google Scholar]
- Tamura, K.; Dudley, J.; Nei, M.; Kumar, S. MEGA4: Molecular evolutionary genetics analysis (MEGA) software version 4.0. Mol. Biol. Evol. 2007, 24, 1596–1599. [Google Scholar]
- Okunji, C.; Okeke, C.; Gugnani, H.; Iwu, M. An antifungal spirostanol saponin from fruit pulp of Dracaena mannii. Int. J. Crude Drug Res. 1990, 28, 193–199. [Google Scholar] [CrossRef]
- Rios, J.-L.; Recio, M.C.; Villar, A. Screening methods for natural products with antimicrobial activity: A review of the literature. J. Ethnopharmacol. 1988, 23, 127–149. [Google Scholar] [CrossRef] [PubMed]
- Hamed, A.A.; El-Shiekh, R.A.; Mohamed, O.G.; Aboutabl, E.A.; Fathy, F.I.; Fawzy, G.A.; Al-Taweel, A.M.; Elsayed, T.R.; Tripathi, A.; Al-Karmalawy, A.A. Cholinesterase Inhibitors from an Endophytic Fungus Aspergillus niveus Fv-er401: Metabolomics, Isolation and Molecular Docking. Molecules 2023, 28, 2559. [Google Scholar] [CrossRef] [PubMed]
- Chambers, M.C.; Maclean, B.; Burke, R.; Amodei, D.; Ruderman, D.L.; Neumann, S.; Gatto, L.; Fischer, B.; Pratt, B.; Egertson, J. A cross-platform toolkit for mass spectrometry and proteomics. Nat. Biotechnol. 2012, 30, 918–920. [Google Scholar] [CrossRef]
- Pluskal, T.; Castillo, S.; Villar-Briones, A.; Orešič, M. MZmine 2: Modular framework for processing, visualizing, and analyzing mass spectrometry-based molecular profile data. BMC Bioinform. 2010, 11, 1–11. [Google Scholar] [CrossRef]
- Paul, S.; Andrew, M.; Owen, O. Baliga Nitin S, Wang Jonathan T, Ramage Daniel, Amin Nada, Schwikowski Benno, Ideker Trey. Cytoscape: A software environment for integrated models of biomolecular interaction networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar]
- Dührkop, K.; Fleischauer, M.; Ludwig, M.; Aksenov, A.A.; Melnik, A.V.; Meusel, M.; Dorrestein, P.C.; Rousu, J.; Böcker, S. SIRIUS 4: A rapid tool for turning tandem mass spectra into metabolite structure information. Nat. Methods 2019, 16, 299–302. [Google Scholar] [CrossRef]
- Dührkop, K.; Shen, H.; Meusel, M.; Rousu, J.; Böcker, S. Searching molecular structure databases with tandem mass spectra using CSI: FingerID. Proc. Natl. Acad. Sci. USA 2015, 112, 12580–12585. [Google Scholar] [CrossRef]
- van Santen, J.A.; Poynton, E.F.; Iskakova, D.; McMann, E.; Alsup, T.A.; Clark, T.N.; Fergusson, C.H.; Fewer, D.P.; Hughes, A.H.; McCadden, C.A. The Natural Products Atlas 2.0: A database of microbially-derived natural products. Nucleic Acids Res. 2022, 50, D1317–D1323. [Google Scholar] [CrossRef]
- Cao, L.; Guler, M.; Tagirdzhanov, A.; Lee, Y.-Y.; Gurevich, A.; Mohimani, H. MolDiscovery: Learning mass spectrometry fragmentation of small molecules. Nat. Commun. 2021, 12, 1–13. [Google Scholar] [CrossRef]
- Goodman, J. Computer Software Review: Reaxys; ACS Publications: Washington, DC, USA, 2009. [Google Scholar]
- Radchenko, M.; Symersky, J.; Nie, R.; Lu, M. Structural basis for the blockade of MATE multidrug efflux pumps. Nat. Commun. 2015, 6, 1–11. [Google Scholar] [CrossRef]
- Chemical Computing Group Inc. Molecular Operating Environment (MOE); Chemical Computing Group Inc.: Quebec, QC, Canada, 2016; Volume 1010. [Google Scholar]
- Elshal, M.; Eid, N.; El-Sayed, I.; El-Sayed, W.; Al-Karmalawy, A.A. Concanavalin-A Shows Synergistic Cytotoxicity with Tamoxifen viaInducing Apoptosis in Estrogen Receptor-Positive Breast Cancer: In Vitro and Molecular Docking Studies. Pharm. Sci. 2021, 28, 76–85. [Google Scholar]
- Ma, C.; Taghour, M.S.; Belal, A.; Mehany, A.B.; Mostafa, N.; Nabeeh, A.; Eissa, I.H.; Al-Karmalawy, A.A. Design and synthesis of new quinoxaline derivatives as potential histone deacetylase inhibitors targeting hepatocellular carcinoma: In silico, in vitro, and SAR studies. Front. Chem. 2021, 9, 725135. [Google Scholar] [CrossRef] [PubMed]
- Khattab, M.; Al-Karmalawy, A.A. Computational repurposing of benzimidazole anthelmintic drugs as potential colchicine binding site inhibitors. Future Med. Chem. 2021, 13, 1623–1638. [Google Scholar] [CrossRef] [PubMed]
- Elmaaty, A.A.; Darwish, K.M.; Chrouda, A.; Boseila, A.A.; Tantawy, M.A.; Elhady, S.S.; Shaik, A.B.; Mustafa, M.; Al-karmalawy, A.A. In Silico and In Vitro Studies for Benzimidazole Anthelmintics Repurposing as VEGFR-2 Antagonists: Novel Mebendazole-Loaded Mixed Micelles with Enhanced Dissolution and Anticancer Activity. ACS Omega 2022, 7, 875–899. [Google Scholar] [CrossRef]
- Mahmoud, D.B.; Bakr, M.M.; Al-Karmalawy, A.A.; Moatasim, Y.; El Taweel, A.; Mostafa, A. Scrutinizing the feasibility of nonionic surfactants to form isotropic bicelles of curcumin: A potential antiviral candidate against COVID-19. AAPS PharmSciTech 2022, 23, 1–12. [Google Scholar] [CrossRef]
- Abd-Alla, H.I.; Kutkat, O.; Sweelam, H.-T.M.; Eldehna, W.M.; Mostafa, M.A.; Ibrahim, M.T.; Moatasim, Y.; GabAllah, M.; Al-Karmalawy, A.A. Investigating the Potential Anti-SARS-CoV-2 and Anti-MERS-CoV Activities of Yellow Necklacepod among Three Selected Medicinal Plants: Extraction, Isolation, Identification, In Vitro, Modes of Action, and Molecular Docking Studies. Metabolites 2022, 12, 1109. [Google Scholar] [CrossRef] [PubMed]
- Ezz Eldin, R.R.; Saleh, M.A.; Alotaibi, M.H.; Alsuair, R.K.; Alzahrani, Y.A.; Alshehri, F.A.; Mohamed, A.F.; Hafez, S.M.; Althoqapy, A.A.; Khirala, S.K.; et al. Ligand-based design and synthesis of N’-Benzylidene-3,4-dimethoxybenzohydrazide derivatives as potential antimicrobial agents; evaluation by in vitro, in vivo, and in silico approaches with SAR studies. J. Enzym. Inhib. Med. Chem. 2022, 37, 1098–1119. [Google Scholar] [CrossRef]
- Hammoud, M.M.; Khattab, M.; Abdel-Motaal, M.; Van der Eycken, J.; Alnajjar, R.; Abulkhair, H.; Al-Karmalawy, A.A. Synthesis, structural characterization, DFT calculations, molecular docking, and molecular dynamics simulations of a novel ferrocene derivative to unravel its potential antitumor activity. J. Biomol. Struct. Dyn. 2022, 1–18. [Google Scholar] [CrossRef]
- Kim, S.-H.; Lu, W.; Ahmadi, M.K.; Montiel, D.; Ternei, M.A.; Brady, S.F. Atolypenes, tricyclic bacterial sesterterpenes discovered using a multiplexed in vitro Cas9-TAR gene cluster refactoring approach. ACS Synth. Biol. 2018, 8, 109–118. [Google Scholar] [CrossRef]
- Shi, Y.; Zhang, J.; Tian, X.; Wu, X.; Li, T.; Lu, C.; Shen, Y. Isolation of 11, 12-seco-Rifamycin W derivatives reveals a cleavage pattern of the rifamycin ansa chain. Org. Lett. 2019, 21, 900–903. [Google Scholar] [CrossRef] [PubMed]
- Lu, C.; Liu, X.; Li, Y.; Shen, Y. Two 18-membered epothilones from Sorangium cellulosum So0157-2. J. Antibiot. 2010, 63, 571–574. [Google Scholar] [CrossRef]
- Bewley, C.A.; Debitus, C.; Faulkner, D.J. Microsclerodermins A and B. Antifungal cyclic peptides from the lithistid sponge Microscleroderma sp. J. Am. Chem. Soc. 1994, 116, 7631–7636. [Google Scholar] [CrossRef]
- Xu, L.-Y.; Quan, X.-S.; Wang, C.; Sheng, H.-F.; Zhou, G.-X.; Lin, B.-R.; Jiang, R.-W.; Yao, X.-S. Antimycins A19 and A20, two new antimycins produced by marine actinomycete Streptomyces antibioticus H74-18. J. Antibiot. 2011, 64, 661–665. [Google Scholar] [CrossRef] [PubMed]
- Zhuravleva, O.; Afiyatullov, S.; Ermakova, S.; Nedashkovskaya, O.; Dmitrenok, P.; Denisenko, V.; Kuznetsova, T. New C14-surfactin methyl ester from the marine bacterium Bacillus pumilus KMM 456. Russ. Chem. Bull. 2010, 59, 2137–2142. [Google Scholar] [CrossRef]
- Tareq, F.S.; Lee, M.A.; Lee, H.-S.; Lee, J.-S.; Lee, Y.-J.; Shin, H.J. Gageostatins A–C, antimicrobial linear lipopeptides from a marine Bacillus subtilis. Mar. Drugs 2014, 12, 871–885. [Google Scholar] [CrossRef]
- Zhang, L.; Zhang, J.; Ren, B.; Lu, W.; Hou, C.; Wang, J.; Ma, X.; Ma, R.; Liu, M.; Liu, Z. Characterization of anti-BCG benz [α] anthraquinones and new siderophores from a Xinjiang desert–isolated rare actinomycete Nocardia sp. XJ31. Appl. Microbiol. Biotechnol. 2020, 104, 8267–8278. [Google Scholar] [CrossRef]
- Schneider, K.; Nachtigall, J.; Hänchen, A.; Nicholson, G.; Goodfellow, M.; Süssmuth, R.D.; Fiedler, H.-P. Lipocarbazoles, secondary metabolites from Tsukamurella pseudospumae Acta 1857 with antioxidative activity. J. Nat. Prod. 2009, 72, 1768–1772. [Google Scholar] [CrossRef]
- Qian-Cutrone, J.; Ueki, T.; Huang, S.; Mookhtiar, K.A.; Ezekiel, R.; Kalinowski, S.S.; Brown, K.S.; Golik, J.; Lowe, S.; Pirnik, D.M. Glucolipsin A and B, two new glucokinase activators produced by Streptomyces purpurogeniscleroticus and Nocardia vaccinii. J. Antibiot. 1999, 52, 245–255. [Google Scholar] [CrossRef]
- Banskota, A.H.; McAlpine, J.B.; Sørensen, D.; Ibrahim, A.; Aouidate, M.; Piraee, M.; Alarco, A.-M.; Farnet, C.M.; Zazopoulos, E. Genomic analyses lead to novel secondary metabolites. J. Antibiot. 2006, 59, 533–542. [Google Scholar] [CrossRef]
- Mondal, H.; Thomas, J. Isolation and Characterization of a Novel Actinomycete Isolated from Marine Sediments and Its Antibacterial Activity against Fish Pathogens. Antibiotics 2022, 11, 1546. [Google Scholar] [CrossRef] [PubMed]
- Gamaleldin, N.M.; Bakeer, W.; Sayed, A.M.; Shamikh, Y.I.; El-Gendy, A.O.; Hassan, H.M.; Horn, H.; Abdelmohsen, U.R.; Hozzein, W.N. Exploration of chemical diversity and antitrypanosomal activity of some red sea-derived actinomycetes using the OSMAC approach supported by LC-MS-based metabolomics and molecular modelling. Antibiotics 2020, 9, 629. [Google Scholar] [CrossRef] [PubMed]
- Sorensen, D. Genome scanning technology reveals an antibacterial compound (ECO-0501) of a new structural class from the vancomycin-producer Amycolatopsis orientalis. In Proceedings of the 44th ICAAC, Washington, DC, USA, 30 October–2 November 2004. [Google Scholar]
- Desai, H.; Wong, R.; Pasha, A.K. A novel way of treating multidrug-resistant enterococci. N. Am. J. Med. Sci. 2016, 8, 229. [Google Scholar] [CrossRef] [PubMed]
- Aristoff, P.A.; Garcia, G.A.; Kirchhoff, P.D.; Showalter, H.H. Rifamycins–obstacles and opportunities. Tuberculosis 2010, 90, 94–118. [Google Scholar] [CrossRef]
- Palma Esposito, F.; Giugliano, R.; Della Sala, G.; Vitale, G.A.; Buonocore, C.; Ausuri, J.; Galasso, C.; Coppola, D.; Franci, G.; Galdiero, M. Combining OSMAC approach and untargeted metabolomics for the identification of new glycolipids with potent antiviral activity produced by a marine Rhodococcus. Int. J. Mol. Sci. 2021, 22, 9055. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Mean Diameter of Inhibition Zone (MDIZ)/mm | |||||||
|---|---|---|---|---|---|---|---|
| SNA | ISP-2 | M1 | R2YE | Czapek | Positive Control | Negative Control | |
| Staphylococcus aureus | - | 24 | 20 | - | - | 22 | - |
| Methicillin-resistant Staphylococcus aureus (MRSA) | - | 22 | 19 | - | - | 24 | - |
| Bacillus cereus | - | 24 | 22 | - | - | 24 | - |
| Salmonella typhimurium | - | - | - | - | - | 18 | - |
| Pseudomonas aeruginosa | - | - | - | - | - | 14 | - |
| Listeria monocytogenes | - | 32 | 27 | - | - | 24 | - |
| E. coli O157:H7 | - | - | - | - | - | 14 | - |
| Minimal Inhibitory Concentration (MIC, µg/mL) | |||||
|---|---|---|---|---|---|
| SNA | ISP-2 | M1 | R2YE | Czapek | |
| Staphylococcus aureus | - | 39 | 39 | - | - |
| Methicillin-resistant Staphylococcus aureus (MRSA) | - | 39 | 39 | - | - |
| Bacillus cereus | - | 19.5 | 19.5 | - | - |
| Salmonella typhimurium | - | - | - | - | - |
| Pseudomonas aeruginosa | - | - | - | - | - |
| Listeria monocytogenes | - | 19.5 | 39 | - | - |
| E. coli O157:H7 | - | - | - | - | - |
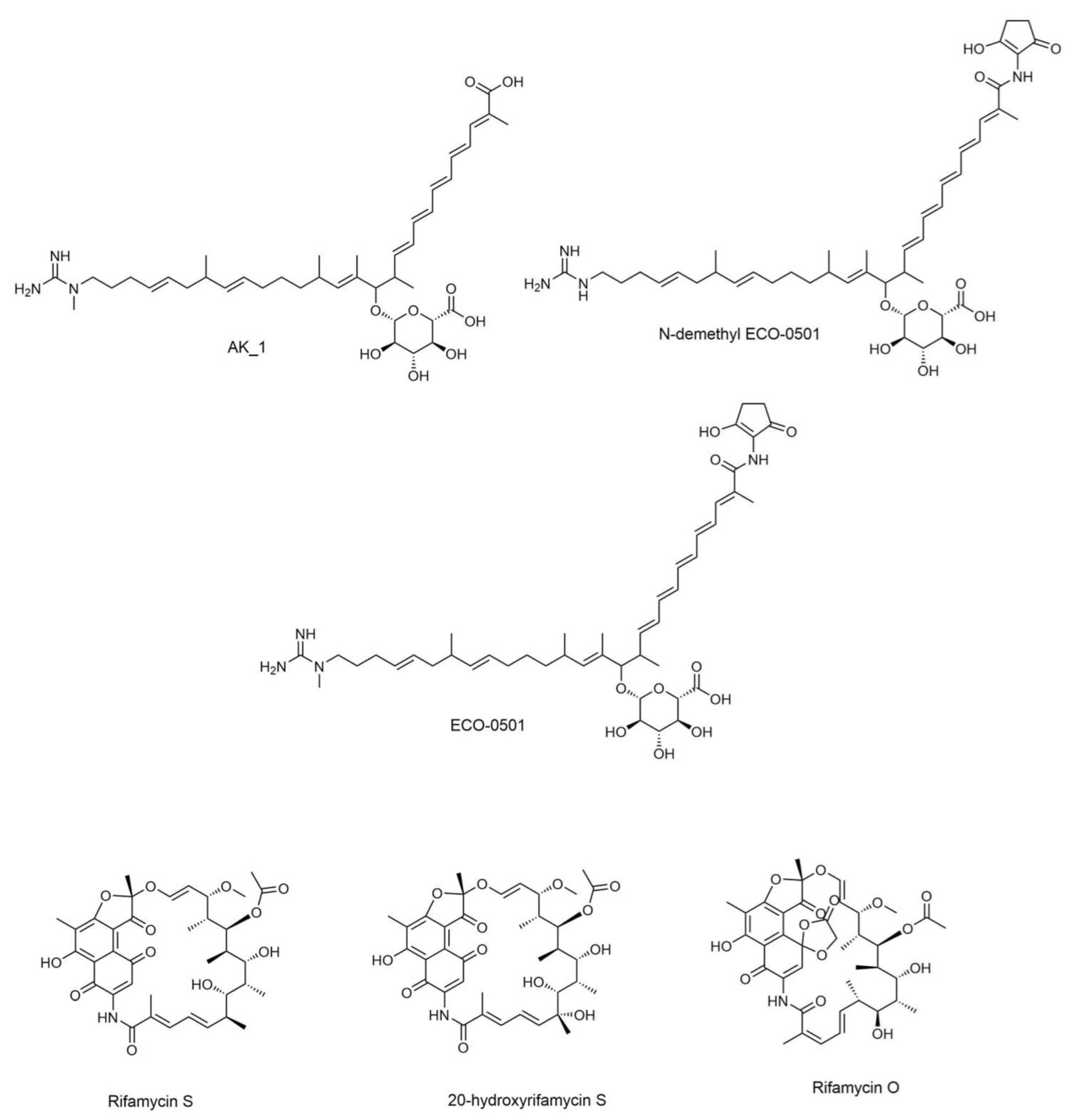
| No. | Rt (Min) | Compound Name | Adduct | Precursor Mass | Molecular Formula | MS/MS Fragmentation Product Ions | Chemical Class |
|---|---|---|---|---|---|---|---|
| 1 | 2.90 | 20-Hydroxyrifamycin S | M-H | 711.289094 | C37H45NO13 | 123.045, 180.1031, 221.0471, 253.0712 | Macrocyclic lactams |
| 2 | 3.21 | Rifamycin S | M-H | 695.294179 | C37H45NO12 | 123.0453, 153.0559, 180.1034, 221.0462 | Macrocyclic lactams |
| 3 | 3.85 | Rifamycin O | M-H | 753.299659 | C39H47NO14 | 119.0862, 154.0744, 163.0772, 192.1029 | Macrocyclic lactams |
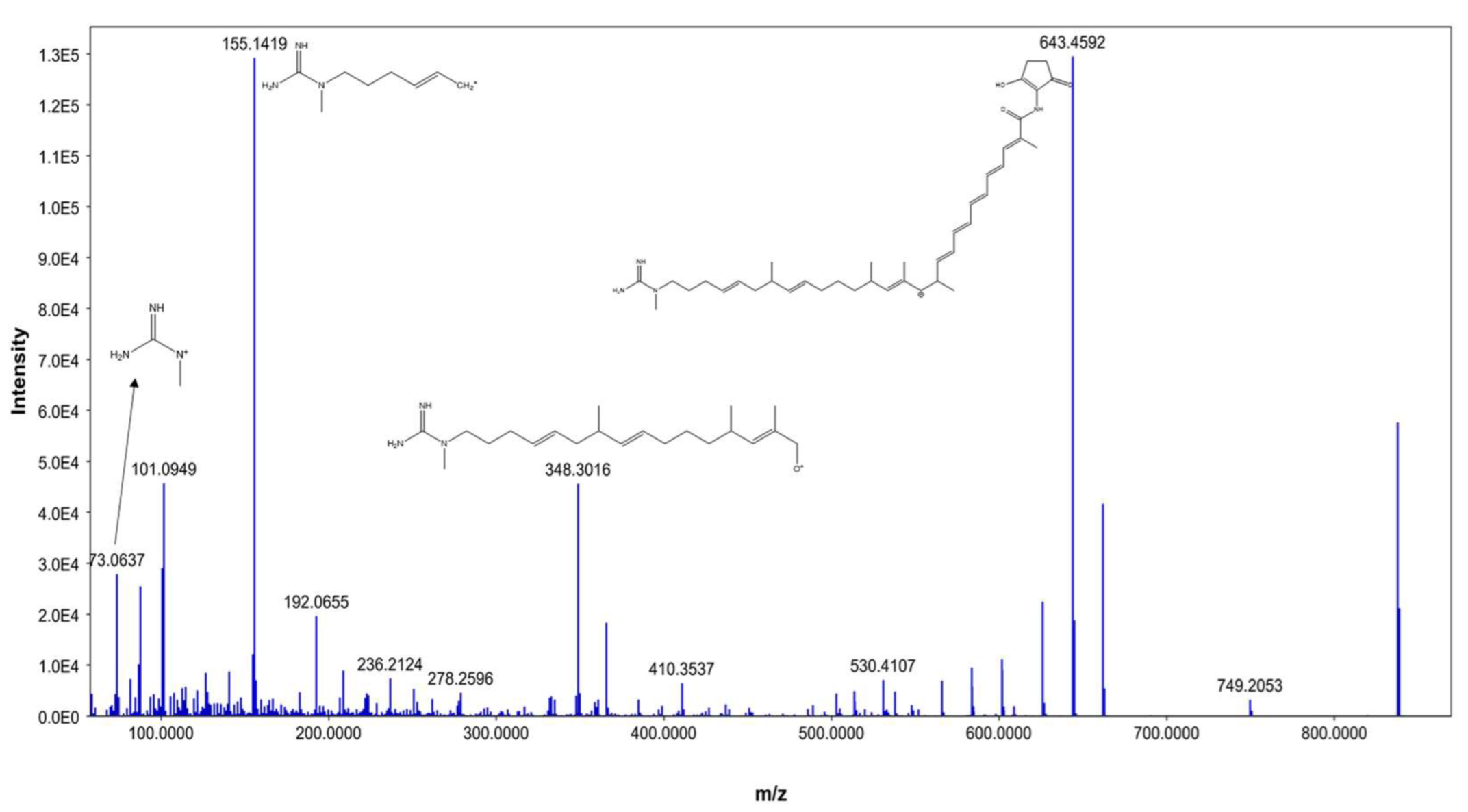
| 4 | 4.13 | AK_1 | M+H | 741.456432 | C41H63N3O9 | 73.056, 101.0861, 155.1322, 548.415 | Linear polyketides |
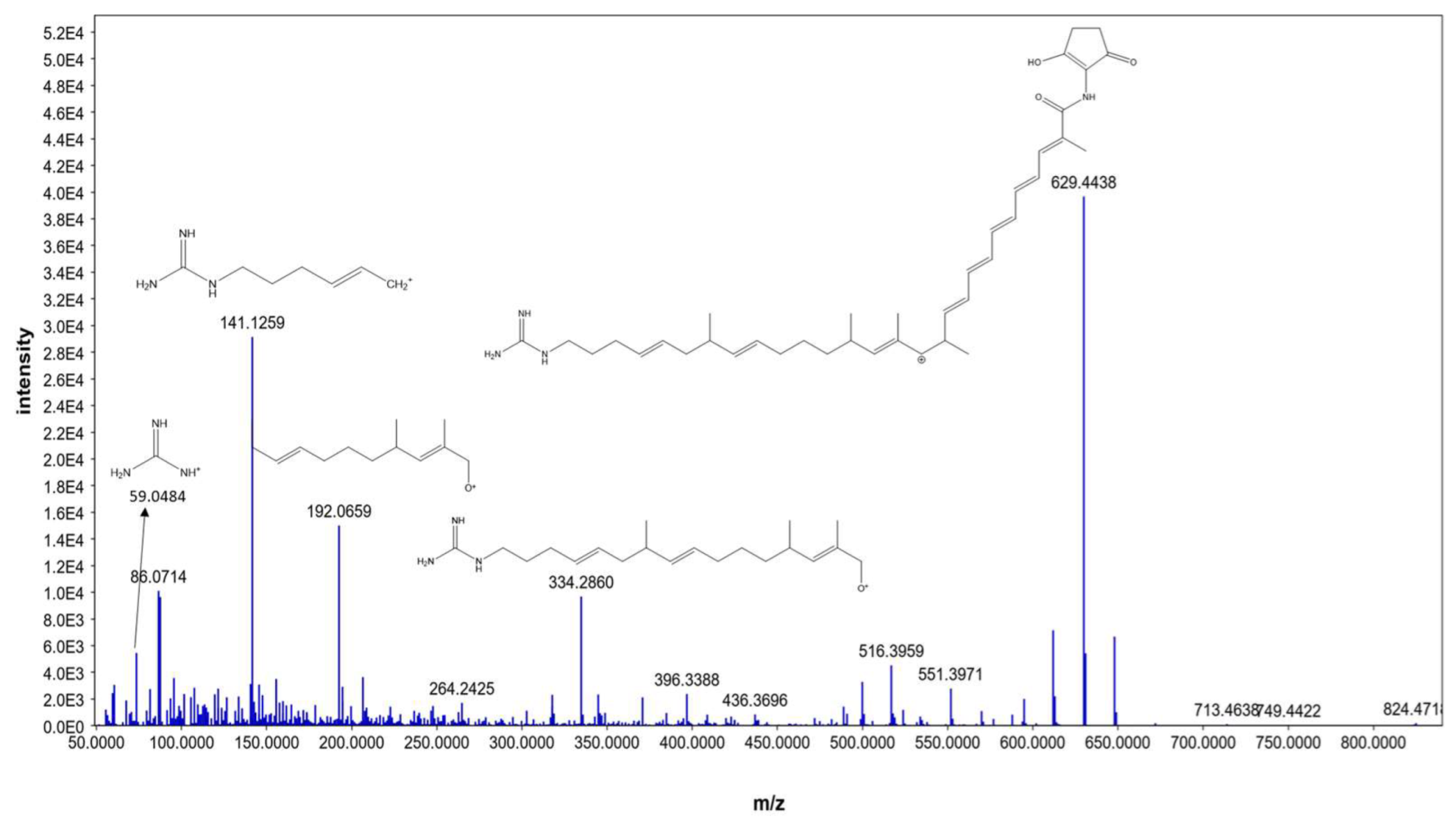
| 5 | 4.41 | N-demethyl ECO-0501 | M+H | 822.477896 | C45H66N4O10 | 86.0714, 141.1259, 192.0659, 629.4438 | Linear polyketides |
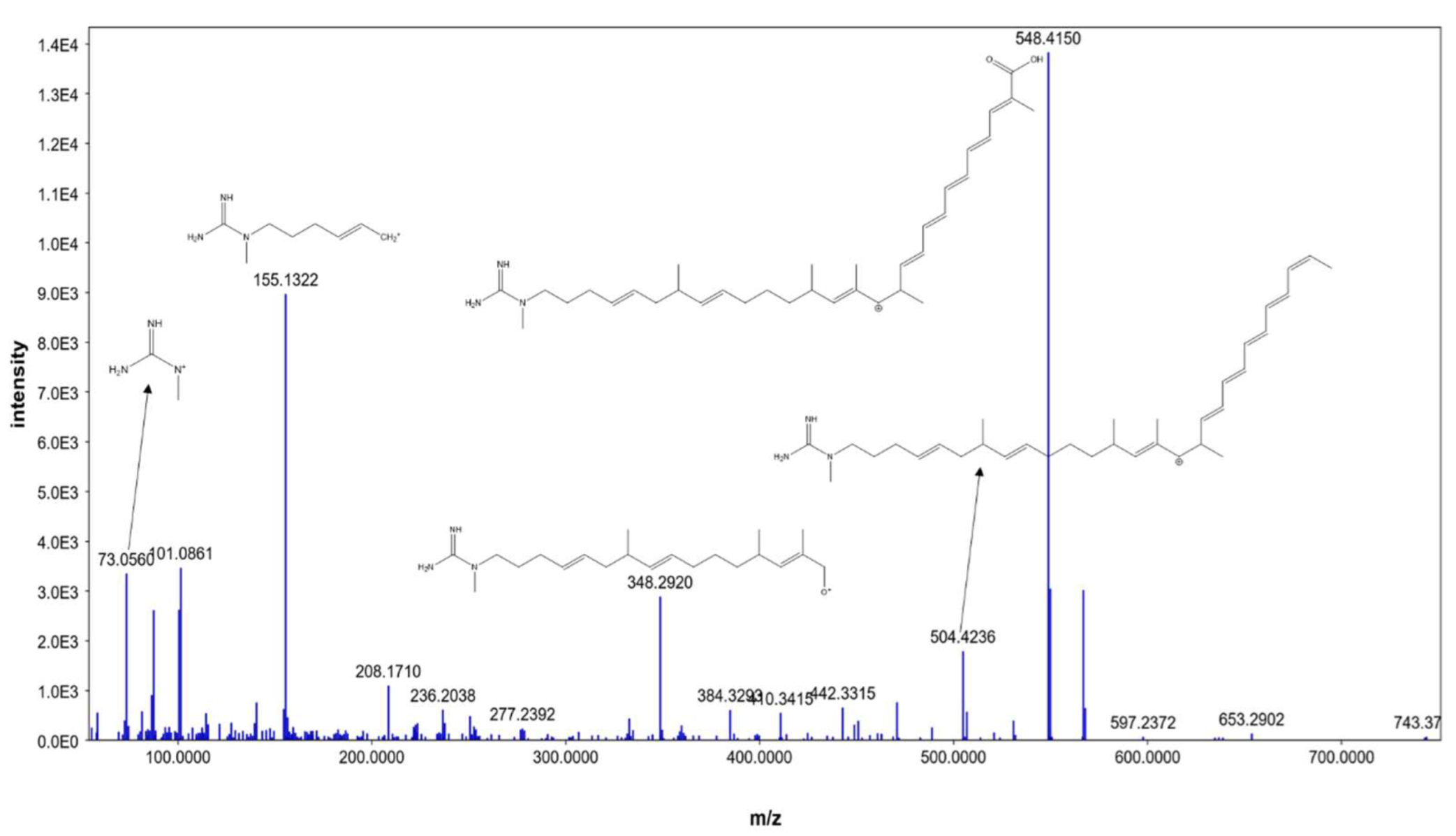
| 6 | 4.48 | ECO-0501 | M+H | 836.493545 | C46H68N4O10 | 101.0949, 155.1419, 348.3016, 643.4592 | Linear polyketides |
| Compound | 3D Interactions | 3D Positioning |
|---|---|---|
| ECO-0501 |  |  |
| AK_1 |  |  |
| N-demethyl ECO-0501 |  |  |
| 4YH |  |  |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hamed, A.A.; Mohamed, O.G.; Aboutabl, E.A.; Fathy, F.I.; Fawzy, G.A.; El-Shiekh, R.A.; Al-Karmalawy, A.A.; Al-Taweel, A.M.; Tripathi, A.; Elsayed, T.R. Identification of Antimicrobial Metabolites from the Egyptian Soil-Derived Amycolatopsis keratiniphila Revealed by Untargeted Metabolomics and Molecular Docking. Metabolites 2023, 13, 620. https://doi.org/10.3390/metabo13050620
Hamed AA, Mohamed OG, Aboutabl EA, Fathy FI, Fawzy GA, El-Shiekh RA, Al-Karmalawy AA, Al-Taweel AM, Tripathi A, Elsayed TR. Identification of Antimicrobial Metabolites from the Egyptian Soil-Derived Amycolatopsis keratiniphila Revealed by Untargeted Metabolomics and Molecular Docking. Metabolites. 2023; 13(5):620. https://doi.org/10.3390/metabo13050620
Chicago/Turabian StyleHamed, Ahmed A., Osama G. Mohamed, Elsayed A. Aboutabl, Fify I. Fathy, Ghada A. Fawzy, Riham A. El-Shiekh, Ahmed A. Al-Karmalawy, Areej M. Al-Taweel, Ashootosh Tripathi, and Tarek R. Elsayed. 2023. "Identification of Antimicrobial Metabolites from the Egyptian Soil-Derived Amycolatopsis keratiniphila Revealed by Untargeted Metabolomics and Molecular Docking" Metabolites 13, no. 5: 620. https://doi.org/10.3390/metabo13050620
APA StyleHamed, A. A., Mohamed, O. G., Aboutabl, E. A., Fathy, F. I., Fawzy, G. A., El-Shiekh, R. A., Al-Karmalawy, A. A., Al-Taweel, A. M., Tripathi, A., & Elsayed, T. R. (2023). Identification of Antimicrobial Metabolites from the Egyptian Soil-Derived Amycolatopsis keratiniphila Revealed by Untargeted Metabolomics and Molecular Docking. Metabolites, 13(5), 620. https://doi.org/10.3390/metabo13050620