
9-Methoxyellipticine: Antibacterial Bioactive Compound Isolated from Ochrosia elliptica Labill. Roots

 , ,
, ,  , and
, and
Abstract
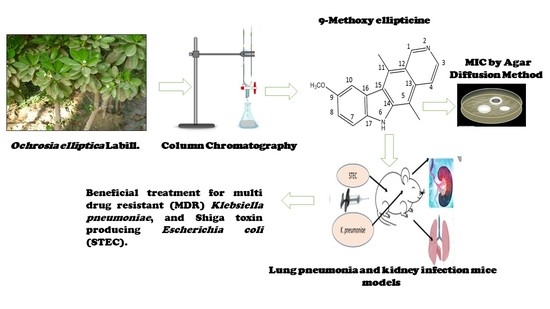
1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Extraction and Isolation
2.3. Biological Assays
2.3.1. Bacterial Strains
2.3.2. Culture Media
2.3.3. Screening for Multidrug-Resistant Bacteria
2.3.4. In Vitro Antibacterial Assay
Agar Disc Diffusion Method
Minimum Inhibitory Concentration (MIC)
2.3.5. In Vivo Study
Animals
Bacterial Strains
K. pneumoniae Mouse Model of Infection
E. coli STEC O157 Mouse Model of Infection
2.3.6. Ex Vivo Analysis
2.3.7. Quantification of Bacterial Shedding of STEC O157
3. Results and Discussion
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- WHO. New Report Calls for Urgent Action to Avert Antimicrobial Resistance Crisis. Available online: https://www.who.int/news/item/29-04-2019-new-report-calls-for-urgent-action-to-avert-antimicrobial-resistance-crisis (accessed on 20 December 2022).
- Nimer, N.A. Nosocomial Infection and Antibiotic-Resistant Threat in the Middle East. Infect. Drug Resist. 2022, 15, 631. [Google Scholar] [CrossRef] [PubMed]
- Asokan, G.V.; Ramadhan, T.; Ahmed, E.; Sanad, H. WHO global priority pathogens list: A bibliometric analysis of Medline-PubMed for knowledge mobilization to infection prevention and control practices in Bahrain. Oman Med. J. 2019, 34, 184. [Google Scholar] [CrossRef]
- Sayk, F.; Hauswaldt, S.; Nitschke, M.; Zuellich, H.; Buening, J.; Wellhoener, P.; Bohnet, S.; Haertel, C.; Steinhoff, J.; Lehnert, H. Successful management of STEC O157 diagnosis and infection control during a large outbreak caused by ESBL-producing STEC O157 (O104: H4). J. Med. Microbiol. 2011, 301, 37–38. [Google Scholar]
- Moulin-Schouleur, M.; Schouler, C.; Tailliez, P.; Kao, M.-R.; Brée, A.; Germon, P.; Oswald, E.; Mainil, J.; Blanco, M.; Blanco, J. Common virulence factors and genetic relationships between O18: K1: H7 Escherichia coli isolates of human and avian origin. J. Clin. Microbiol. 2006, 44, 3484–3492. [Google Scholar] [CrossRef]
- Venezia, S.; Kondratyeva, K.; Carattoli, A. Klebsiella pneumoniae: A major worldwide source and shuttle for antibiotic resistance. FEMS Microbiol. Rev. 2017, 41, 252–275. [Google Scholar] [CrossRef] [PubMed]
- Ramos-Castañeda, J.A.; Ruano-Ravina, A.; Barbosa-Lorenzo, R.; Paillier-Gonzalez, J.E.; Saldaña-Campos, J.C.; Salinas, D.F.; Lemos-Luengas, E.V. Mortality due to KPC carbapenemase-producing Klebsiella pneumoniae infections: Systematic review and meta-analysis: Mortality due to KPC Klebsiella pneumoniae infections. J. Infect. 2018, 76, 438–448. [Google Scholar] [CrossRef] [PubMed]
- Patil, S.A.; Patil, S.A.; Ble-González, E.A.; Isbel, S.R.; Hampton, S.M.; Bugarin, A. Carbazole Derivatives as Potential Antimicrobial Agents. Molecules 2022, 27, 6575. [Google Scholar] [CrossRef]
- Eloutify, Y.T.; El-Shiekh, R.A.; Ibrahim, K.M.; Elshimy, R.; Avula, B.; Katragunta, K.; Khan, I.A.; Meselhy, M.R. Bioassay-Guided Isolation of Antimicrobial Components and LC/QToF Profile of Plumeria obtusa: Potential for the Treatment of Antimicrobial Resistance. ACS Omega 2023, 8, 6476–6491. [Google Scholar] [CrossRef]
- El-Shiekh, R.A.; Elhemely, M.; Naguib, I.A.; Bukhari, S.I.; Elshimy, R. Luteolin 4-Neohesperidoside Inhibits Clinically Isolated Resistant Bacteria In Vitro and In Vivo. Molecules 2023, 28, 2609. [Google Scholar] [CrossRef]
- El-Shiekh, R.A.; Abdelmohsen, U.R.; Ashour, H.M.; Ashour, R.M. Novel antiviral and antibacterial activities of Hibiscus schizopetalus. Antibiotics 2020, 9, 756. [Google Scholar] [CrossRef]
- Salem, M.A.; El-Shiekh, R.A.; Hashem, R.A.; Hassan, M. In vivo antibacterial activity of star anise (Illicium verum Hook.) Extract Using Murine MRSA skin infection model in relation to its metabolite profile. Infect. Drug Resist. 2021, 14, 33–48. [Google Scholar] [CrossRef] [PubMed]
- El-Shiekh, R.A.; Hassan, M.; Hashem, R.A.; Abdel-Sattar, E. Bioguided isolation of antibiofilm and antibacterial pregnane glycosides from Caralluma quadrangula: Disarming multidrug-resistant pathogens. Antibiotics 2021, 10, 811. [Google Scholar] [CrossRef] [PubMed]
- Ali, N.B.; El-Shiekh, R.A.; Ashour, R.M.; El-Gayed, S.H.; Abdel-Sattar, E.; Hassan, M. In Vitro and In Vivo Antibiofilm Activity of Red Onion Scales: An Agro-Food Waste. Molecules 2023, 28, 355. [Google Scholar] [CrossRef] [PubMed]
- El-shiekh, R.A.; Al-Mahdy, D.A.; Hifnawy, M.S.; Abdel-sattar, E.A. Pharmacognostical study of Ochrosia elliptica Labill.(Apocynaceae). J. Appl. Pharm. Sci. 2019, 9, 049–057. [Google Scholar]
- Chen, A.-H.; Liu, Q.-L.; Ma, Y.-L.; Jiang, Z.-H.; Tang, J.-Y.; Liu, Y.-P.; Chen, G.-Y.; Fu, Y.-H.; Xu, W. A new monoterpenoid indole alkaloid from Ochrosia elliptica. Nat. Prod. Res. 2017, 31, 1490–1494. [Google Scholar] [CrossRef]
- Chang, H.D.; Helbig, C.; Tykocinski, L.; Kreher, S.; Koeck, J.; Niesner, U.; Radbruch, A. Expression of IL-10 in Th memory lymphocytes is conditional on IL-12 or IL-4, unless the IL-10 gene is imprinted by GATA-3. Eur. J. Immunol. 2007, 37, 807–817. [Google Scholar] [CrossRef]
- El-shiekh, R.A.; Al-Mahdy, D.A.; Hifnawy, M.S.; Tzanova, T.; Evain-Bana, E.; Philippot, S.; Bagrel, D.; Abdelsattar, E.A. Chemical and biological investigation of Ochrosia elliptica Labill. cultivated in Egypt. Rec. Nat. Prod. 2017, 11, 552–557. [Google Scholar] [CrossRef]
- El-Shiekh, R.A.; Al-Mahdy, D.A.; Hifnawy, M.S.; Abdel-Sattar, E. Biological and Chemical Assessment of Ochrosia elliptica Labill Leaves. Arab. J. Sci. Eng. 2021, 46, 5247–5255. [Google Scholar] [CrossRef]
- Lu, H.; Liu, M.; Lu, W.; Wang, C.; Wang, G.; Dong, W.; Wang, X.; Chen, H.; Tan, C. Repurposing ellipticine hydrochloride to combat colistin-resistant extraintestinal pathogenic E. coli (ExPEC). Front. Microbiol. 2020, 11, 806. [Google Scholar] [CrossRef]
- Vehar, B.; Hrast, M.; Kovač, A.; Konc, J.; Mariner, K.; Chopra, I.; O’Neill, A.; Janežič, D.; Gobec, S. Ellipticines and 9-acridinylamines as inhibitors of D-alanine: D-alanine ligase. Bioorg. Med. Chem. 2011, 19, 5137–5146. [Google Scholar] [CrossRef]
- Mahmud, P.I.; Yaacob, W.; Ibrahim, N. Antibacterial activity of alkaloid extracts from Ochrosia oppositifolia. Sains Malays. 2017, 46, 1279–1284. [Google Scholar] [CrossRef]
- Ho, T.L.; Hsieh, S.Y. Regioselective synthesis of ellipticine. Helv. Chim. Acta 2006, 89, 111–116. [Google Scholar] [CrossRef]
- Elshimy, R.; Zedan, H.; Elmorsy, T.H.; Khattab, R.A. A Study on Multidrug-Resistant Escherichia coli Clinical Isolates from Different Hospitals in Greater Cairo. Microb. Drug Resist. 2021, 27, 1420–1432. [Google Scholar] [CrossRef]
- Rana, E.; Rania, A.K.; Hamdallah, Z.; Alaa, E.-D.S.H.; Tarek, H.E. Study on prevalence and genetic discrimination of methicillin-resistant Staphylococcus aureus (MRSA) in Egyptian hospitals. Afr. J. Microbiol. Res. 2018, 12, 629–646. [Google Scholar] [CrossRef]
- Rajapaksha, P.; Elbourne, A.; Gangadoo, S.; Brown, R.; Cozzolino, D.; Chapman, J. A review of methods for the detection of pathogenic microorganisms. Analyst 2019, 144, 396–411. [Google Scholar] [CrossRef] [PubMed]
- Sabino, C.P.; Wainwright, M.; Ribeiro, M.S.; Sellera, F.P.; Dos Anjos, C.; da Silva Baptista, M.; Lincopan, N. Global priority multidrug-resistant pathogens do not resist photodynamic therapy. J. Photochem. Photobiol. B Biol. 2020, 208, 111893. [Google Scholar] [CrossRef]
- Bauer, A. Antibiotic susceptibility testing by a standardized single disc method. Am. J. Clin. Pathol. 1966, 45, 149–158. [Google Scholar] [CrossRef]
- Wiegand, I.; Hilpert, K.; Hancock, R. Agar and broth dilution methods to determine the minimal inhibitory concentration (MIC) of antimicrobial substances. Nat. Protoc. 2007, 3, 163–175. [Google Scholar] [CrossRef]
- Parihar, M.; Rakshit, A.; Meena, V.S.; Gupta, V.K.; Rana, K.; Choudhary, M.; Tiwari, G.; Mishra, P.K.; Pattanayak, A.; Bisht, J.K. The potential of arbuscular mycorrhizal fungi in C cycling: A review. Arch. Microbiol. 2020, 202, 1581–1596. [Google Scholar] [CrossRef]
- Tan, S.; Gao, J.; Li, Q.; Guo, T.; Dong, X.; Bai, X.; Yang, J.; Hao, S.; He, F. Synergistic effect of chlorogenic acid and levofloxacin against Klebsiella pneumonia infection in vitro and in vivo. Sci. Rep. 2020, 10, 20013. [Google Scholar] [CrossRef]
- Malorny, B.; Hoorfar, J.; Hugas, M.; Heuvelink, A.; Fach, P.; Ellerbroek, L.; Bunge, C.; Dorn, C.; Helmuth, R. Interlaboratory diagnostic accuracy of a Salmonella specific PCR-based method. Int. J. Food Microbiol. 2003, 89, 241–249. [Google Scholar] [CrossRef] [PubMed]
- Festy, B.; Poisson, J.; Paoletti, C. A new DNA intercalating drug: Methoxy-9-ellipticine. FEBS Lett. 1971, 17, 321–323. [Google Scholar] [CrossRef] [PubMed]
- Okdah, L.; Le Page, S.; Olaitan, A.O.; Dubourg, G.; Hadjadj, L.; Rolain, J.-M. New therapy from old drugs: Synergistic bactericidal activity of sulfadiazine with colistin against colistin-resistant bacteria, including plasmid-mediated colistin-resistant mcr-1 isolates. Int. J. Antimicrob. Agents 2018, 51, 775–783. [Google Scholar] [CrossRef] [PubMed]
- Pfalzgraff, A.; Weindl, G. Intracellular lipopolysaccharide sensing as a potential therapeutic target for sepsis. Trends Pharmacol. Sci. 2019, 40, 187–197. [Google Scholar] [CrossRef] [PubMed]
- Butler, T.; Rahman, H.; Al-Mahmud, K.; Islam, M.; Bardhan, P.; Kabir, I.; Rahman, M. An animal model of haemolytic--uraemic syndrome in shigellosis: Lipopolysaccharides of Shigella dysenteriae I and S. flexneri produce leucocyte-mediated renal cortical necrosis in rabbits. Br. J. Exp. Pathol. 1985, 66, 7. [Google Scholar] [PubMed]
- Al-Mathkhury, H.J.F.; Flaih, M.T.; Tawfiq, H.K. Histopathological effects of S. xylosus peptidoglycan in comparison to E. coli lipopolysaccharide in the urinary tract of mice. Turk. J. Med. Sci. 2012, 42, 1278–1285. [Google Scholar] [CrossRef]
- Güran, M.; Şanlıtürk, G.; Kerküklü, N.R.; Altundağ, E.M.; Yalçın, A.S. Combined effects of quercetin and curcumin on anti-inflammatory and antimicrobial parameters in vitro. Eur. J. Pharmacol. 2019, 859, 172486. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Liu, J.; Zhuang, Y.; Bai, L.-P.; Yuan, Q.; Zheng, S.; Liao, K.; Khan, M.A.; Wu, Q.; Luo, C. Identification of an IKKβ inhibitor for inhibition of inflammation in vivo and in vitro. Pharmacol. Res. 2019, 149, 104440. [Google Scholar] [CrossRef] [PubMed]
- Sappati, S.; Hassanali, A.; Gebauer, R.; Ghosh, P. Nuclear quantum effects in a HIV/cancer inhibitor: The case of ellipticine. J. Chem. Phys. 2016, 145, 205102. [Google Scholar] [CrossRef]
- Tian, L.-X.; Li, X.-Y.; Tang, X.; Zhou, X.-Y.; Luo, L.; Ma, X.-Y.; Tang, W.-Q.; Yu, J.; Ma, W.; Yang, X. Ellipticine conveys protective effects to lipopolysaccharide-activated macrophages by targeting the JNK/AP-1 signaling pathway. Inflammation 2020, 43, 231–240. [Google Scholar] [CrossRef]
- Wen, H.-l.; Yang, G.; Dong, Q.-r. Ellipticine inhibits the proliferation and induces apoptosis in rheumatoid arthritis fibroblast-like synoviocytes via the STAT3 pathway. Immunopharmacol. Immunotoxicol. 2017, 39, 219–224. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Ye, C.; Mulati, M.; Sun, L.; Qian, F. Ellipticine blocks synergistic effects of IL-17A and TNF-α in epithelial cells and alleviates severe acute pancreatitis-associated acute lung injury. Biochem. Pharmacol. 2020, 177, 113992. [Google Scholar] [CrossRef] [PubMed]
- Nurhayati, A.P.D.; Santoso, M.; Setiawan, E.; Cintyasari, A.; Abdillah, S.; Lianingsih, F.; Indiani, A.M.; Ghaissani, S.S. In vitro Test of Deazaelliptisina 1 Compound on Phagocytotic activity of Macrophage Cell. Sustain. Nat. Resour. Res. 2021, 27, 6–12. [Google Scholar]
- Wang, C.; Lu, H.; Liu, M.; Wang, G.; Li, X.; Lu, W.; Dong, W.; Wang, X.; Chen, H.; Tan, C. Effective antibacterial and antihemolysin activities of ellipticine hydrochloride against Streptococcus suis in a mouse model. Appl. Environ. Microbiol. 2021, 87, e03165-20. [Google Scholar] [CrossRef] [PubMed]
- Melton-Celsa, A.R. Shiga toxin (Stx) classification, structure, and function. Microbiol. Spectr. 2014, 2, 2–4. [Google Scholar] [CrossRef]
- Sulaiman, M.; Jannat, K.; Nissapatorn, V.; Rahmatullah, M.; Paul, A.K.; de Lourdes Pereira, M.; Rajagopal, M.; Suleiman, M.; Butler, M.S.; Break, M.K.B. Antibacterial and Antifungal Alkaloids from Asian Angiosperms: Distribution, Mechanisms of Action, Structure-Activity, and Clinical Potentials. Antibiotics 2022, 11, 1146. [Google Scholar] [CrossRef]
- Porras, G.; Chassagne, F.; Lyles, J.T.; Marquez, L.; Dettweiler, M.; Salam, A.M.; Samarakoon, T.; Shabih, S.; Farrokhi, D.R.; Quave, C.L. Ethnobotany and the role of plant natural products in antibiotic drug discovery. Chem. Rev. 2020, 121, 3495–3560. [Google Scholar] [CrossRef]
- Liang, M.; Ge, X.; Xua, H.; Ma, K.; Zhang, W.; Zan, Y.; Efferth, T.; Xue, Z.; Hua, X. Phytochemicals with activity against methicillin-resistant Staphylococcus aureus. Phytomedicine 2022, 100, 154073. [Google Scholar] [CrossRef]
- Dabrovolskas, K.; Jonuškienė, I.; Sutkuvienė, S.; Gudeika, D. Synthesis and evaluation of antibacterial and antioxidative activities of carbazole derivatives. Chemija 2020, 31, 42–51. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Zone of Inhibition in mm (Mean ± SD) | |||
|---|---|---|---|---|
| STEC O157 | MDR Klebsiella pneumoniae | Bacillus cereus | Methicillin-Resistant Staphylococcus aureus (MRSA) | |
| Negative control | 0.0 ± 0.0 * | 0.0 ± 0.0 * | 0.0 ± 0.0 * | 9.0 ± 0.0 * |
| PEN | 0.0 ± 0.0 * | 6.0 ± 0.0 * | 0.0 ± 0.0 * | 0.0 ± 0.0 * |
| GEN | 20.0 ± 1.00 # | 9.0 ± 1.00 * | ||
| VAN | - | - | 9.0 ± 1.00 * | 18.0 ± 1.00 # |
| 9-methoxyellipticine | 18 ± 1 # | 21 ± 0.577 # | 5 ± 0.577 * | 3 ± 1.00 * |
| PEN + 9-methoxyellipticine | 23.0 ± 0.577 # | 22.0 ± 0.577 # | 6 ± 0.577 * | 11 ± 0.577 * |
| GEN + 9-methoxyellipticine | 25.0 ± 1.00 # | 23.0 ± 1.00 # | - | - |
| VAN + 9-methoxyellipticine | - | - | 18.0 ± 1.00 * | 23.0 ± 1.00 # |
| Sample | MIC (µg/mL) | |||
|---|---|---|---|---|
| Shiga-Toxin-Producing Escherichia coli O157 (STEC O157) | MDR Klebsiella pneumoniae | Methicillin-Resistant Staphylococcus aureus (MRSA) | MDR Bacillus cereus | |
| 9-methoxyellipticine | 512 (1853) * | 256 (926.52) * | >2048 | >2048 |
| GEN | 2 (4.18) * | 10 (20.93) * | - | - |
| VAN | - | - | 2 | 1 |
| Normal Control Group | Model Group | 9-Methoxyellipticine-Treated Group | Gentamycin (GEN)-Treated Group | |
|---|---|---|---|---|
| Thickened interalveolar septa | − | +++ | + | + |
| Perivascular edema | − | +++ | + | − |
| Hemorrhage | − | +++ | − | − |
| RBCs in alveolar lamina | − | +++ | + | − |
| Normal Control Group | Model Group | 9-Methoxyellipticine-Treated Group | Gentamycin (GEN)-Treated Group | |
|---|---|---|---|---|
| Necrobiotic changes in tubular epithelium | − | +++ | + | + |
| Interstitial inflammatory cell infiltration | − | + | − | + |
| Interstitial hemorrhage | − | + | − | − |
| Subcapsular hemorrhage | − | +++ | − | − |
| Renal cast | − | − | + | − |
| Negative Control | Positive Control STEC O157 | STEC O157 and 9-Methoxyellipticine | STEC O157 and GEN | Negative Control | |
|---|---|---|---|---|---|
| Day zero | 2 × 104 | 3 × 106 | 4 × 104 | 4 × 106 | 36 × 101 |
| Day 4 (before challenge and after treatment with streptomycin) | 2 × 101 | 1 × 102 | 0 | 1 × 101 | 10 × 101 |
| Day 7 (after challenge) | 2 × 101 | Death | 9 × 107 | 1 × 107 | 10 × 106 |
| Day10 (after treatment for 3 successive days with GEN i.p., once daily) | 2 × 101 | Death | 10 × 103 | 2 × 103 | 7 × 108 |
| Day 17 (after treatment for 7 days with 9-methoxyellipticine orally, once daily) | 2 × 101 | Death | 5 × 101 | NA | NA |
| Kidney | zero | Death | zero | 1 × 101 | zero |
| Normal Control | Model Control K. pneumoniae | 9-Methoxyellipticine-Treated Group | GEN-Treated Group | |
|---|---|---|---|---|
| Lung | zero | Death | 2 × 101 | 6 × 102 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Elshimy, R.; Khawagi, W.Y.; Naguib, I.A.; Bukhari, S.I.; El-Shiekh, R.A. 9-Methoxyellipticine: Antibacterial Bioactive Compound Isolated from Ochrosia elliptica Labill. Roots. Metabolites 2023, 13, 643. https://doi.org/10.3390/metabo13050643
Elshimy R, Khawagi WY, Naguib IA, Bukhari SI, El-Shiekh RA. 9-Methoxyellipticine: Antibacterial Bioactive Compound Isolated from Ochrosia elliptica Labill. Roots. Metabolites. 2023; 13(5):643. https://doi.org/10.3390/metabo13050643
Chicago/Turabian StyleElshimy, Rana, Wael Y. Khawagi, Ibrahim A. Naguib, Sarah I. Bukhari, and Riham A. El-Shiekh. 2023. "9-Methoxyellipticine: Antibacterial Bioactive Compound Isolated from Ochrosia elliptica Labill. Roots" Metabolites 13, no. 5: 643. https://doi.org/10.3390/metabo13050643
APA StyleElshimy, R., Khawagi, W. Y., Naguib, I. A., Bukhari, S. I., & El-Shiekh, R. A. (2023). 9-Methoxyellipticine: Antibacterial Bioactive Compound Isolated from Ochrosia elliptica Labill. Roots. Metabolites, 13(5), 643. https://doi.org/10.3390/metabo13050643