Abstract
Baillonella toxisperma is a medicinal plant used in northern Gabon to treat microbial diseases. It is a plant well-known by local populations, but very few studies have focused on the molecules responsible for the antibacterial activities of B. toxisperma. This study proposes a dereplication strategy based on molecular networking generated from HPLC-ESI-Q/TOF data, allowing investigation of the molecules responsible for the antibacterial activity of B. toxisperma. From this strategy, eighteen compounds were putatively identified. All of these compounds belonged mainly to five families of natural compounds, including phenylpropanolamines, stilbenes, flavonoids, lignans and phenolic glycosides. The chemical study carried out from the bark of B. toxisperma allowed us to identify, for the first time, compounds such as resveratrol and derivatives, epicatechin, epigallocatechin and epigallocatechin gallate. In addition, antibacterial activity (diffusion method and microdilution) and cytotoxicity (Cell Counting Kit-8 (CCK-8 Assay)) in vitro were evaluated. The crude ethanolic extract, as well as the fractions of B. toxisperma, showed significant antibacterial activity. However, the ethanolic fractions F2 and F4 presented high antibacterial activity compared to the crude extract. Cytotoxicity studies on colon-cancer cells (Caco-2) and human keratinocyte cells (HaCaT) showed moderate cytotoxicity in both cell types. This study clearly shows the therapeutic potential of the ethanolic extract of the bark of B. toxisperma and provides information on the phytochemical composition and bioactive compounds of the plant.
1. Introduction
Baillonella toxisperma Pierre (B. toxisperma), commonly known as moabi in Cameroon and adzap in Gabon and Equatorial Guinea, is found wild in the forests of intertropical Africa [1]. Naturally durable, B. toxisperma is known for its pinkish-brown wood and the oil extracted from its fruit [2], which gives it multiple uses in traditional medicine. In the Cameroonian pharmacopoeia, decoctions of aqueous extracts of its bark are used in treatment of hemorrhoids, sexually transmitted infections, diarrhea, malaria, vaginal mycosis [3] and gastrointestinal and bronchopulmonary disorders [4]. In Gabon, this plant is used in traditional medicine in treatment of several pathologies, notably rheumatic pains, mycosis, wounds, ringworms and malaria. It is also used as an additive in other remedies [5]. B. toxisperma belongs to the Sapotaceae family, one of the most important plant families commonly distributed in the tropical forests of Africa, Australia and South America [6]. Indeed, Sapotaceae is one of the eight species of the richest family of angiosperms, it has about 1250 species and 58 genera [7,8]. Generally, the plants belonging to this family are known for their numerous pharmacological properties, as is the case for B. toxisperma. Since the 2000s, this plant has been the subject of several studies. The studies of Saha et al. showed strong antioxidant activity of the wood of the plant [4]. Other studies have also shown that the plant has antimicrobial and antifungal activities [9,10]. B. toxisperma also improves sexual performance and reproductive dysfunction induced by stress [3]. Although several biological studies have been conducted on the bark, fruit and leaves of B. toxisperma [11,12], very little work has been published on the compounds responsible for the biological activities of the plant. One of the few studies that isolated a compound (3-hydroxy uridine, a plant growth inhibitor) was performed by Hajime et al. [13]. Thus, in the present study, we propose a molecular networking (MN) dereplication approach based on high-resolution liquid chromatography/mass spectrometry (LC-HRMS/MS) data to more efficiently and rapidly target compounds present in the ethanolic extracts of B. toxisperma [14]. Molecular networking (MN) is a valuable tool for revealing the metabolomes of plants, humans, microorganisms and animals [15]. It can not only annotate compounds in complex matrices based on their tandem mass spectrometry (MS/MS) characteristics but also group them into molecular families based on their spectral similarities, thus facilitating visualization of structurally identical molecules within the same chemical family and structurally different molecules distributed in different chemical families [16,17]. This approach allows researchers to identify known compounds in order to focus on unknown compounds that may potentially be of biological interest [18]. The main purpose of our study was not only to have good visibility of the chemical composition of the bark of B. toxisperma but also to identify the molecules responsible for its antibacterial activity. Thus, a combined MN and LC-MS/MS approach was used. Considering the use of this plant in Gabonese traditional medicine, it was essential to test its innocuousness toward certain cell lines, including CaCo-2 cells and HaCaT cells. The structures of the compounds annotated in the ethanolic extract of B. toxisperma are mainly composed of phenolic compounds known for their antibacterial, antioxidant and antiproliferative activities on several cancer-cell lines [19,20,21,22]. To the best of our knowledge, the chemical composition and cytotoxic activities of B. toxisperma bark are described for the first time in this study.
2. Materials and Methods
2.1. Plant Material
Fresh trunk bark of B. toxisperma was collected in January 2019 from the village of Nvane Essabeigne in the Woleu-Ntem province of northern Gabon (0°43′0″ N, 11°37′60″ E). The botanical identification was confirmed by Professor Henri Paul Bourobou. A specimen (M.E.M 002) was deposited at the National Herbarium of Gabon (Libreville, Gabon).
2.2. Extraction and Fractionation
B. toxisperma bark was dried and then ground. From this powder (400 g), extraction by maceration under agitation was carried out for 24 h with 1500 mL of 70% ethanol. The extract obtained was filtered under a vacuum and then concentrated with a rotary evaporator to obtain 12.5 g of ethanolic crude extract. The crude extract was fractionated with Flash chromatography on an 80 g Reveleris® Grace Silica PF-50SIHC-F0080 cartridge (Rheinland-Pfalz, Germany) using a CH2Cl2 gradient, increasing the proportions of MeOH (100:0 to 0:100) with a fixed flow rate 60 mL/min [17]. The fractions obtained in the Flash chromatography were pooled with TLC thin-layer chromatography and were then analyzed in HPLC-QTOF-HRES to search for potential new compounds.
2.3. Data-Dependent LC-HR-ESI-MS2 Analysis
Data-dependent LC-HR-ESI-MS2 analysis was performed using a previously described method [17]. The extracts were analyzed using high-performance liquid chromatography coupled to a Sunfire® C18 analytical column (150 × 2.1 mm, 3.5 µm particle diameters; Waters, Milford, MA, USA). A standard elution gradient was used, with mobile phases A:H2O and B:MeOH, each containing 0.1% formic acid (HCOOH). Both solvents were eluted at a flow rate of 0.25 mL/min using a standard gradient of MeOH:H2O + 0.1% HCOOH (5:95 to 100:0 for 40 min). The temperature of the column oven was set to 40 °C and the injection volume to 5 µL. Detection was performed with a diode-array UV detector connected in series with an Agilent 6530 hybrid quadrupole time-of-flight mass spectrometer (Agilent Technologies, Massy, France) equipped with an ESI source, which was compatible with analytes with various polarity ranges. The ionization parameters were a 320 °C capillary temperature, a source voltage of 3500, a 10 L/min sheath-gas flow rate, 180 V fragmentation potential and 35 psi nebulizer pressure. The bypass valve was set to waste for the first 3 min. The analyzer provided satisfactory accuracy in the measurements, with a mass resolution of 11,000 at m/z 922, and the high-resolution measurements provided significant certainty in the calculated raw formulas for the observed fragment ions, this quality of data is necessary for the molecular network approach employed. MS scans were measured in the full-scan mode of m/z, between 100 and 1700, with a scan time of 0.1 s. The MS1 scan was followed by an MS2 scan of the three most intense ions above an absolute threshold of 5000 counts [17]. The selected parent ions were fragmented with a collision energy of 45 eV and an isolation window of 1.3 u.m.a [23]. The calibration solution contained two internal reference masses (Purine, C5H4N4, m/z 121.050873 and HP-921 [hexakis-(1H,1H,3H-tetrafluoropentoxy) phosphazene], C18H18O6N3P3F24, m/z 922.0098). A permanent MS/MS exclusion list criterion was established to avoid oversampling of the internal calibrant. LC-UV and MS data acquisition and processing were performed using MassHunter Workstation software (Agilent Technologies, Massy, France) [17]. The raw LC-HR-ESI-MS/MS data in .d format (Agilent) were converted to mzXML format with MSConvert software [24]. With MZmine 2 V32 software [25], all mzXML formats were processed. Then, the .mgf spectral data file and its .csv metadata file were exported to the GNPS web platform. A molecular network was created using the spectral clustering algorithm, with a cosine score of 0.65 and a minimum of six matched peaks. The molecular network data were analyzed and visualized using Cytoscape software (ver. 3.9.0) [26].
2.4. Bacterial Germs Tested
The bacterial carrier used in our study consisted of two reference bacterial strains (Escherichia coli ATCC 25922, Escherichia coli ATCC 8739) and three clinical strains, including multi-drug-resistant Klebsiella pneumoniae, extended-spectrum beta-lactamase-producing Escherichia coli and Salmonella enterica. All of these bacterial strains were obtained at the Centre Interdisciplinaire de Recherches Médicales de Franceville (CIRMF) in Gabon.
2.5. Antibacterial Assays
The diffusion method was used to study sensitivity of microorganisms to plant extracts. Bacterial colonies were used to prepare the inoculum in order to have a density equivalent to that of 0.5 McFarland. Flood inoculation of the microbial suspension was performed, the agar was allowed to dry for 10 min. After drying, sterilized Wattman 1 paper disks impregnated with 20 µL of crude extract or fraction prepared at a concentration of 100 μg/mL (diluted in 1% DMSO) were placed on the Petri plates. Then, the latter were incubated for 18–24 h at 37 °C. Ticarcillin, tetracycline and gentamicin were used as positive controls.
The minimum inhibitory concentrations (MICs) of the crude extracts and fractions were determined with the microdilution method on 96-well microplates [27]. A series of seven dilutions of each extract (double dilutions ranging from 0.0049 to 5 mg/mL) were made in Muller–Hinton Nutrient Broth (Liofilchem, Roseto, Italy). To determine the minimum bactericidal concentrations (MBCs), nutrient agar was inoculated with 100 μL of the contents of the wells (concentrations greater than or equal to the MIC). The MBC was determined after a 24 h incubation at 37 °C. Antibacterial samples were considered to be bactericidal with MBC/MIC ratios equivalent to 1 or 2 and bacteriostatic if the MBC/MIC ratio was equivalent to 4 or 16 [27].
2.6. Cell Culture
Human colon epithelial cells, Caco-2 (accession No 86010202, European collection of authenticated cell cultures (ECACC), Salisbury, UK), were grown in monolayers in Dulbecco’s Modified Eagle’s Medium (DMEM), supplemented with 10% inactivated calcium-free fetal calf serum (Gibco Laboratories, Grand Island, NY, USA), 100 U/mL penicillin/0.1 mg/mL streptomycin (PanBiotech GmbH, Aidenbach, Germany) and 2 mM L- glutamine (PanBiotech GmbH, Germany), at 37 °C and in an atmosphere containing 5% CO2 (CO2 Incubator; Thermo Scientific, Marietta, OH, USA) [28]. HaCaT (immortalized human keratinocyte) cells (accession No. 300493, Cell Lines Service GmbH, Eppelheim, Germany) were cultured in DMEM medium that was low in calcium (0.03 mM) and supplemented with 10% calcium-free fetal calf serum, 100 U/mL penicillin/0.1 mg/mL streptomycin and 2 mM L-glutamine (PanBiotech GmbH, Germany).
2.7. Cell Viability CCK-8 Assays
Cell viability was determined with the Cell Counting Kit-8 (CCK-8) assay (Dojindo Laboratories, Kumamoto, Japan) according to the manufacturer’s protocol [28]. Cells were seeded in 96-well plates at densities of 2 × 104 cells/cm2 and 1.2 × 104 cells/cm2 for the Caco-2 and HaCaT cells, respectively. After 4 days of culture for the HaCaT cells and 6 days of culture for the Caco-2 cells, the samples were diluted in DMEM (at increasing concentrations ranging from 100 to 500 µg/mL) and contacted with cells for 24 h at 37 °C (CO2 Incubator, Thermo Scientific, Marietta, USA) at a rate of 100 µL/well. WST-8 was then added to the final concentration of 5% for 2 h at 37 °C in the dark. The concentration of reduced WST-8 was measured at a 450 nm wavelength using the spectrophotometer (SpectraMax® iD3, Molecular Devices, San Jose, CA, USA) against “blanks or counter assays” containing all reagents for each condition except cells. Results are presented as percent viability compared to the control (DMEM 2% DMSO). Cell viability was calculated relative to the percentage of live cells in the DMSO control on GraphPad Prism 9.3.0 (463) using the following formula: % Cell viability = Mean OD of treated cells/Mean OD of DMSO control cells. The data were expressed as the mean ± standard deviation (SD) of triplicate independent experiments (N = 3 three independent experiments) for each line, in triplicate for each experiment (n = 9), and analyzed using a one-way analysis of variance (ANOVA), the Kruskal-Wallis test and Dunn’s post hoc test. * p = 0.0304; *** p = 0.0007 and **** p < 0.0001 were considered to be statistically significant.
3. Results and Discussion
3.1. Metabolite Profiling of B. toxisperma Bark from LC-MS/MS Analysis
Figure 1 shows the BPC (Base Peak Chromatogram) of the ethanolic extract of B. toxisperma obtained with HPLC-UV-MS in positive ion mode. This chromatogram allowed us to visualize the majority of the chromatographic peaks on broad polarity ranging from 5 to 100% of organic solvent.
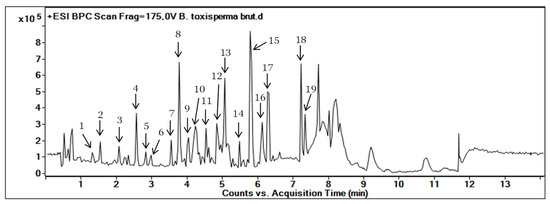
Figure 1.
Base Peak Chromatogram (BPC) of the ethanolic extract of B. toxisperma obtained with HPLC-UV-MS/MS. The number on each peak corresponds to the numbers in Table 1.
The step of identification of the metabolites present in the ethanolic crude extract of the B. toxisperma trunk bark was carried out using a double identification (LC-MS/MS and molecular networks). These two identification steps allowed us not only to determine the elemental composition of the molecules present in the plant but also to have an overview of the families of molecules grouped in the plant. These two steps were made possible by exploiting high-resolution mass spectrometry data. The datasets were then annotated with the theoretical precise masses of the compounds contained in the public databases already described above. As in Table 1, from the crude extract of B. toxisperma, a total of nineteen compounds were putatively annotated. These metabolites mainly belong to five families of natural compounds: phenylpropanolamines, flavonoids, stilbenes, lignans and phenolic glycosides. Some of the major peaks were tentatively identified manually based on exact masses through comparison on public databases such as the Dictionary of Natural Products: https://dnp.chemnetbase.com (accessed on 22 October 2022) [29], PubChem and ChemSpider. Some of these compounds have also been identified in several other plants belonging to the family Sapotaceae [30]. Thus, on this basis, only compounds 1, 3, 4, 7, 8, 9, 10 and 11 were identified, as epigallocatechin; kelampayoside A; epigallocatechin gallate; resveratrol; β-conidendrin; 3,5-O-dimethyl resveratrol; methyl resveratrol and isorhapontin, respectively.

Table 1.
LC-HRMS determination of compounds in the ethanolic crude extract of B. toxisperma in positive ionization mode.
The confidence levels of the compound annotations were performed with respect to the guidelines published by the Metabolomics Society [31]. Level 0, Unambiguous 3D Structure: the isolated, pure compound, including full stereochemistry; Level 1, Confident 2D Structure: uses reference standard-matching or full 2D structure elucidation; Level 2, Probable Structure: matched to literature data or databases with diagnostic evidence; Level 3, Possible Structure or Class: the most likely structure, isomers possible, substance class or substructure match; Level 4, Unknown Feature of Interest.
3.2. MS/MS–Molecular Networking-Based Dereplication
The analysis of the ethanolic crude extract of B. toxisperma was performed using the MN approach combined with LC-MS/MS as proposed on the GNPS website: http://gnps.ucsd.edu (accessed on 19 April 2022). All obtained HRESIMS/MS spectra were processed using MZmine2 software, thus, two types of files were obtained, in .mgf and _quant.csv formats. From these two files, the MS/MS spectra were searched in the spectral libraries hosted by the GNPS platform and the results were annotated as part of this dereplication strategy. To refine the structural information of the molecular network, an in silico structure annotation from GNPS Library Search was incorporated into the network using the MolNetEnhancer workflow [32].
The molecular network was visualized using Cytoscape 3.9.0 software. This molecular network contained one large cluster and sixteen small clusters (number of nodes n ≥ 3) that could be linked into molecular families or groups (Figure 2). As well as the manual exploitation of the MS/MS data, these results highlighted the presence of five major molecular families in the extract of B. toxisperma: phenylpropanolamines (cluster A), stilbenes (cluster B), flavonoids (cluster C and C’), lignans (cluster D) and phenolic glycosides (cluster E). Cluster B was composed of seven nodes, three of which had m/z 259.097 (C15H14O3); m/z 229.086 (C14H12O3) and m/z 243.102 (C16H8O3), corresponding to three stilbenes, respectively: 3,5-O-dimethyl resveratrol, resveratrol and methyl resveratrol. On the other hand, clusters C and C’ revealed six compounds of the flavonoid family, they are epicatechin (m/z 291.086; C15H14O6); epigallocatechin (m/z 307.081; C15H14O7); epigallocatechin gallate (m/z 459.092; C22H18O11); leucocyanidin (m/z 306.2064; C15H14O7); ampelopsin, still known as dihydromyricetin (m/z 320.2220; C15H12O8); and leucodelphinidin (m/z 322.2379; C15H14O8). Cluster-C compounds have been described by Baky et al. (2016) to be in several plants of the Sapotaceae family [30], and those of cluster C’ described by Ma et al. (2011) in a Fabaceae used in traditional Chinese medicine [33], which generally confirms the presence of these compounds at the level of angiosperms and, in particular, at the level of B. toxisperma (Figure 2 and Figure 3). Finally, phenylpropanolamines (cluster A) such as norephedrine (m/z 152.107; C9H13NO); N-benzoyl-L-phenylalaninol (m/z 256.133; C16H17NO2); lignans (cluster D), such as the example of conidendrin (m/z 357.369; C20H20O6) and phenolic glycosides (cluster E), such as kelampayoside A (m/z 496.203; C20H30O13), were also identified. Consistently with these results, the phenylpropanolamines appeared to be the largest family, with many associated nodes. Flavonoids, stilbenes and lignans were also well-represented in this ethanolic extract, with a fairly large cluster (number of nodes n ≥ 5), whereas phenolic glycosides were less numerous. Several compounds in cluster A were fragmented in the source, the fragment ions were then fragmented in MS/MS, which explains the redundancy of the information generated for this cluster with several nodes. Indeed, since the fragment ions generated in the source had MS/MS spectra very similar to the MS/MS spectrum of the parent ion, they were linked with a high degree of similarity (Table 2). As for the other clusters, they were not fragmented in the source, therefore, they contained nodes that represented only the molecular ions without information redundancy. We also noted the presence of four clusters where n ≥ 5, named zone F in Figure 2. In fact, the majority of the ion fragments present within these clusters could not be identified. This absence suggests that they are potentially new compounds.
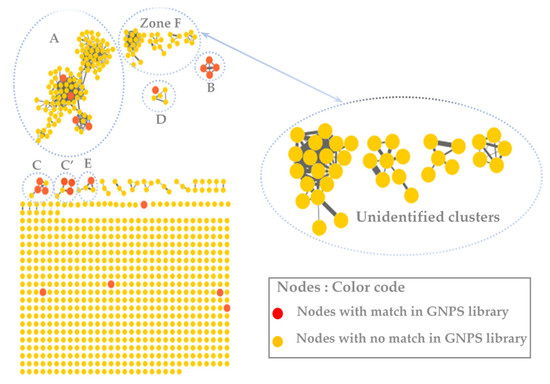
Figure 2.
The molecular network of the ethanolic crude extract of B. toxisperma obtained with the GNPS platform and visualized with Cytoscape 3.9.0 software. The letters A, B, C, C,’ D and E represent different clusters with matches on the GNPS databases.
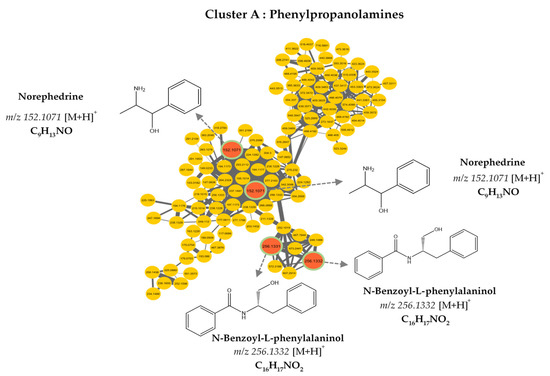
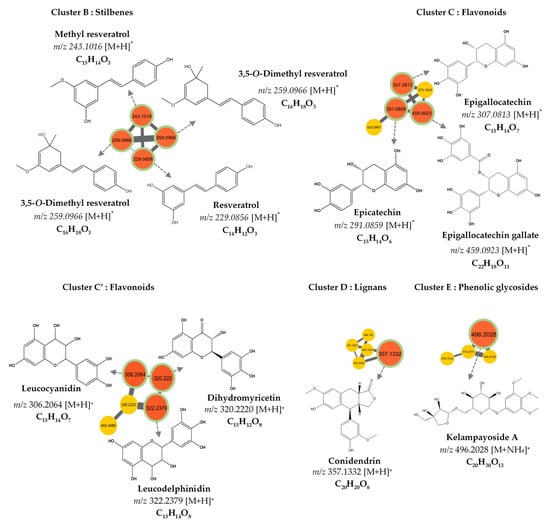
Figure 3.
Molecular network analysis of the ethanolic crude extract of B. toxisperma. Nodes of clusters A (phenylpropanolamines), B (stilbenes), C, and C’ (flavonoids), D (lignans) and E (phenolic glycosides) were putatively annotated from the GNPS spectral library. Nodes in red represent nodes with a match in the GNPS library and those in yellow represent nodes with no match.
Table 2.
Compounds putatively identified in the molecular network from the ethanolic crude extract of B. toxisperma.
3.3. Antibacterial Activity
The antibacterial activity of the ethanolic extract of Baillonella toxisperma was evaluated with five bacterial strains (Escherichia coli ATCC 25992, Escherichia coli ATCC 8739, multi-drug-resistant Klebsiella pneumoniae, Salmonella enterica and extended-spectrum beta-lactamase-producing Escherichia coli) with Gram negativity. The sensitivity tests of the plant extracts toward these microorganisms allowed us to determine the inhibition zone diameters (IZD) of the crude extract and the fractions. The results of the zones of inhibition are recorded in Table 3. Standard antibiotics (Ticarcillin, Tetracycline and Gentamicin) showed IZD between 10 ± 0.00 and 30 ± 0.00 mm in all bacterial strains studied. No inhibition zones were found around the disks impregnated with 1% DMSO. These results indicate that zones of inhibition are variable from one bacterial strain to another and from one extract or fraction to another. Considering the ethanolic crude extract of B. toxisperma (Bt EtOH Ce), we observed inhibition diameters that ranged from 14 ± 0.47 to 22 ± 0.82 mm. The best diameters were obtained for the Klebsiella pneumoniae MDR (IZD 20 ± 0.00 mm) and Escherichia coli ESBL (IZD 22 ± 0.82 mm) strains. On the other hand, the lowest inhibition diameters were registered for Salmonella enterica (IZD 14 ± 0.47 mm) and Escherichia coli ATCC 25992 (IZD 15 ± 0.00 mm). The tests performed on the fractions showed inhibition diameters varying between 9 ± 0.00 mm and 21.33 ± 0.47 mm for the Escherichia coli ATCC 25992 and Klebsiella pneumoniae MDR strains, respectively. Only the Escherichia coli ATCC 25992 strain was sensitive to all fractions and to the crude extract. Fraction F7 showed no inhibition diameter for the Escherichia coli ATCC 8739 or Escherichia coli ESBL strain. Fractions F2 and F4 showed better inhibition diameters in the majority of strains. This means that the antibacterial compounds of the ethanolic extract of Baillonella toxisperma were mainly eluted in the Bt EtOH F2 and Bt EtOH F4 fractions. The minimum inhibitory concentrations (MICs) and minimum bactericidal concentrations (MBCs), reported in Table 4, ranged from 0.31 to 5 mg/mL and 2.5 to 5 mg/mL, respectively. In order to determine the bactericidal or bacteriostatic modes of action of the tested samples, the MBC/MIC ratios were calculated. The results obtained were interpreted as follows: when the MBC/MIC ratio was between 1 and 2, the extract or fraction was bactericidal, and when this ratio was between 4 and 16, the extract was considered bacteriostatic [27]. On this basis, the results in Table 4 show that fraction Bt EtOH F2 is bactericidal against Escherichia coli ATCC 8739, Escherichia coli ATCC 25992 and Salmonella enterica (MBC/MIC ratio = 2) and bacteriostatic against Klebsiella pneumoniae MDR and Escherichia coli ESBL (MBC/MIC ratio = 4). Fraction Bt EtOH F4 shows bactericidal action in all strains (MBC/MIC ratio = 2). The remarkable antibacterial activity of the ethanolic extract of B. toxisperma trunk bark can be attributed to the presence of several phenolic compounds. Indeed, previous studies have demonstrated the antimicrobial and antioxidant activities of some compounds present in the bark of B. toxisperma, such as isorhapontin [34,35], biochanin A [36], resveratrol and its derivatives [37,38], epicatechin and epigallocatechin gallate [39,40,41]. An annotation based on mass spectrometry data was also performed on the most active fractions in order to see more clearly the different compounds present and potentially resting antimicrobial activities on the active fractions of B. toxisperma (Figure 4).
Table 3.
Inhibition-zone diameters (IZD) of crude extract and ethanolic fractions of B. toxisperma.
Table 4.
Minimum inhibitory concentrations (MICs) and bactericidal concentrations (MBCs) of crude extract and ethanolic fractions of B. toxisperma.
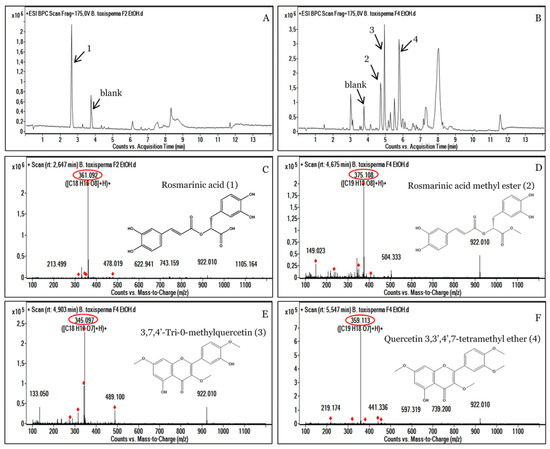
Figure 4.
HPLC-UV-MS analyses of the active ethanolic fractions F2 (A) and F4 (B) of Baillonella toxisperma bark. Mass spectrum of [M+H]+ ions of rosmarinic acid (C); rosmarinic acid methyl ester (D); 3,7,4’,-trimethylquercetin (E) and quercetin 3,3’,4’,7-tetramethyl ether (F). The red circle represents the exact masses of the identified compounds.
3.4. Analysis of the Most Active Fractions, F2 and F4
The fractions that showed the highest antibacterial activity were analyzed with mass spectrometry. The results of this analysis showed that fractions F2 and F4 had several ion pairs. Fraction F2 showed a majority compound of m/z 361.092 [C18H16O8+] at a retention time (RT) of 2.647 min, which corresponds to rosmarinic acid (1) (Figure 4A,C). Furthermore, the fraction F4 showed three major peaks, m/z 375.108 [C19H18O8+]; m/z 345.097 [C18H16O7+] and m/z 359.113 [C19H18O7+], corresponding to rosmarinic acid methyl ester (2); 3,7,4’,-trimethylquercetin (3) and quercetin 3,3’,4’,7-tetramethyl ether (4), respectively (Figure 4B,D,E,F). The presence of all these phenolic compounds in the F2 and F4 fractions could indeed justify the strong antibacterial activities of the plant B. toxisperma. Indeed, previous studies on the plant have shown that polyphenols, tannin and flavonoids are responsible for its antibacterial activities [9]. Several other studies have shown strong antimicrobial activities of rosmarinic acid mainly present in the F2 fraction of B. toxisperma [42,43,44,45]. Similarly, several studies have shown antibacterial and antioxidant activities of quercetin and its derivatives [46,47,48]. In view of our results, we can attribute the antimicrobial activity of the F2 and F4 fractions to these different compounds.
3.5. Cytotoxicity of the ethanolic crude extract of B. toxisperma
The objective of this activity was to evaluate the effect of Baillonella toxisperma bark on the overall cell proliferation of two cell lines: HaCaT and Caco-2. The cytotoxic assay was performed at different extract concentrations ranging from 10 to 500 µg/mL (Figure 5). The IC50 value was determined from the percentage of cell viability compared to the control. The results show that at up to 50 µg/mL, the B. toxisperma extract was well-tolerated by both cell lines. On the other hand, at 100 µg/mL onward, some toxicity was observed in both cell lines. Based on the different IC50 values, HaCaT cells seem to be more sensitive than Caco-2 cells, with respective IC50 values of 21.78 µg/mL and 40.54 µg/mL. These results seem very consistent not only with the traditional use of B. toxisperma but also with the different chemical groups found in the crude extract [49]. Indeed, phenolic compounds are known for their multiple properties: antimicrobial, antioxidant, antiapoptotic, anti-inflammatory and anticarcinogenic [50,51,52,53]. More specifically, certain compounds present in the bark of B. toxisperma, such as resveratrol, kelampayoside A, biochanin A and rosmarinic acid, are known to have various beneficial activities on the proliferations of several cell lines [54,55,56,57,58,59]. These compounds may therefore be partly responsible for the observed activity.
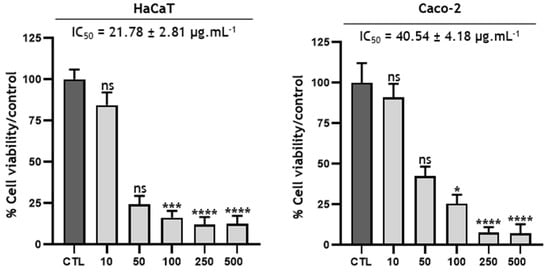
Figure 5.
Cytotoxicity of Baillonella toxisperma Pierre bark on HaCaT and Caco-2 cells. Statistical tests were performed via reading the Kruskal-Wallis test and Dunn’s post hoc test (ns p > 0.9999; * p = 0.0304; *** p = 0.0007; **** p < 0.0001). Differences were observed in comparison with the control (CTL).
4. Conclusions
In this study, the antibacterial properties and in vitro cytotoxicity of Baillonella toxisperma were investigated using a combined mass spectrometry and molecular array approach. Thus, the combination of these two identification techniques allowed determination, for the first time, of not only the different chemical groups present in this plant but also the molecules potentially responsible for the antibacterial activities of B. toxisperma. Among the compounds putatively identified, phenolic compounds such as (i) rosmarinic acid, (ii) quercetin 3,3’,4’,7 tetramethyl ether and (iii) 3,7,4’-tris-O-methyl quercetin were identified in the most active fractions. This identification is not exhaustive but brings new information on the chemical composition of B. toxisperma with the presence of several polyphenols, such as resveratrol and its derivatives. B. toxisperma bark, at very low concentrations, reduces not only the metabolic activity of colon-cancer-cell lines (CaCo-2), but also that of human keratinocyte (HaCaT) cells, which encourages further research on the molecules responsible for this inhibitory activity. All of these results showed therapeutic potential of B. toxisperma and justified its use in traditional medicine.
Author Contributions
Conceptualization, R.R. and J.-P.O.; methodology, E.C. and E.D.; investigation, M.E.M., D.H. and C.S.O.; resources, R.R., J.-P.O., B.S.K. and F.M.; data curation, E.C. and E.D.; writing—original draft preparation, M.E.M.; writing—review and editing, F.M.; project administration, F.M. and B.S.K.; funding acquisition, F.M. and B.S.K. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
The authors wish to thank Elvis Otogo N’nang for helpful discussions and Mehdi Beniddir for data acquisition.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Segers, A. Détermination des Visiteurs Floraux de Trois Essences Ligneuses à Haute Valeur Commerciale: Baillonella toxisperma Pierre, Afzelia Bipindensis Harms et Erythrophleum Suaveolens (Guill. & Perr.) Brenan. 72. Available online: https://www.semanticscholar.org/paper/D%C3%A9termination-des-visiteurs-floraux-de-trois-%C3%A0-%3A-et-Segers/66554ccb59f9560950888148134b2600bef7b542 (accessed on 19 February 2023).
- Ngueguim, J.R.; Dondjang, J.P.; Onana, J.; Ijang, P.T.; Zapfack, L.; Noumi, V.N.; Kengne, O.C.; Solefack, C.M. Moabi (Baillonella toxisperma Pierre): Arbre à Usage Multiple de Forêt Dense Humide Du Cameroun. Int. J. Biol. Chem. Sci. 2012, 5, 2395–2406. [Google Scholar] [CrossRef]
- Simo, R.; Ateba, S.B.; Zingue, S.; Pieme, A.; Njamen, D. Baillonella toxisperma Improves Sexual Performance and Protects against Stress-Induced Reproductive Dysfunction in Male Wistar Rats. J. Phytopharm. 2019, 8, 117–123. [Google Scholar] [CrossRef]
- Saha, J.-B.T.; Abia, D.; Dumarçay, S.; Ndikontar, M.K.; Gérardin, P.; Ngamveng Noah, J.; Perrin, D. Antioxidant Activities, Total Phenolic Contents and Chemical Compositions of Extracts from Four Cameroonian Woods: Padouk (Pterocarpus Soyauxii Taubb), Tali (Erythrophleum Suaveolens), Moabi (Baillonella toxisperma), and Movingui (Distemonanthus Benthamianus). Ind. Crops Prod. 2013, 41, 71–77. [Google Scholar] [CrossRef]
- Ndiade-Bourobou, D.; Vaillant, A.; Favreau, B.; Gayrin, E.; Bouvet, J.M. Isolation and Characterization of 15 Nuclear Microsatellite Markers for Baillonella toxisperma Pierre (Sapotaceae), a Low-Density Tree Species of Central Africa. Mol. Ecol. Resour. 2009, 9, 1135–1138. [Google Scholar] [CrossRef] [PubMed]
- Pennington, T.D. Sapotaceae. In Flowering Plants Dicotyledons: Celastrales, Oxalidales, Rosales, Cornales, Ericales; Kubitzki, K., Ed.; la série de Livres The Families and Genera of Vascular Plants; Springer: Berlin/Heidelberg, Germany, 2004; pp. 390–421. ISBN 978-3-662-07257-8. [Google Scholar]
- Khayi, S.; Gaboun, F.; Pirro, S.; Tatusova, T.; El Mousadik, A.; Ghazal, H.; Mentag, R. Complete Chloroplast Genome of Argania Spinosa: Structural Organization and Phylogenetic Relationships in Sapotaceae. Plants 2020, 9, 1354. [Google Scholar] [CrossRef] [PubMed]
- Swenson, U.; Richardson, J.E.; Bartish, I.V. Multi-Gene Phylogeny of the Pantropical Subfamily Chrysophylloideae (Sapotaceae): Evidence of Generic Polyphyly and Extensive Morphological Homoplasy. Cladistics 2008, 24, 1006–1031. [Google Scholar] [CrossRef]
- Essama, S.H.R.; Nyegue, M.A.; Foe, C.N.; Tamo, S.P.B.; Etoa, F.X. In Vitro Evaluation of the Antifungal Activity of Extracts of Baillonella toxisperma (Pierre), a Sapotaceae, on the Growth of Some Human Pathogenic Yeasts. AJPP 2015, 9, 299–306. [Google Scholar] [CrossRef]
- Fodouop, M.; Tamo, S.P.B.; Pegnyemb, D.E.; Etoa, F.X.; de Paula, R.A.; Barbosa, A.A.T.; Machado, S.R.S.; Saraiva, M.A.F.; de Moraes, C.A.; Mantovani, H.C.; et al. In Vitro Antibacterial Activity of Baillonella toxisperma (Pierre) Extracts against Staphylococcus Aureus, Salmonella Typhi, Proteus Mirabilis and Bacillus Cereus F3748. AJMR 2015, 9, 2088–2094. [Google Scholar] [CrossRef]
- Guy Roussel, T.N.; Martin, F.; Janvier Aimé, Y.F.; Ferdinand Lanvin, E.E.; Ruth Edwige, D.K.; Boris, A.K.; Laure, N.J.; Enyong, O.J. Antihyperglycemic and Antihyperlipidemic Activities of Hydroethanolic Extract of the Fruit of Baillonella toxisperma in Streptozotocin-Induced Diabetic Rats. Metab. Open 2022, 15, 100199. [Google Scholar] [CrossRef]
- Benoit, M.Z.; Florence, N.T.; Raceline, G.K.; Yannick, F.B.; Alide, W.N.M.; Theophile, D. Antidiabetic Effects of Aqueous Extract of Baillonella toxisperma Pierre (Sapotacae) in Streptozotocin-Induced Diabetic Rats. J. Med. Plants Stud. 2021, 9, 28–37. [Google Scholar] [CrossRef]
- Ohigashi, H.; Kaji, M.; Sakaki, M.; Koshimizu, K. 3-Hydroxyuridine, an Allelopathic Factor of an African Tree, Baillonella toxisperma. Phytochemistry 1989, 28, 1365–1368. [Google Scholar] [CrossRef]
- Kim, J.G.; Le, T.P.L.; Han, J.S.; Cho, Y.B.; Lee, D.; Lee, M.K.; Hwang, B.Y. Molecular Networking-Guided Isolation of Cycloartane-Type Triterpenoids from Curculigo Orchioides and Their Inhibitory Effect on Nitric Oxide Production. ACS Omega. 2022, 7, 26853–26862. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, C.; Miyata, R.; Wakayama, S.; Kumazawa, S. New Acylated Flavonoid Isolated from Thai Bee Pollen Using Molecular Networking Analysis and Determination of Its Catechol-O-Methyltransferase Inhibitory Activity. Phytochem. Lett. 2023, 53, 239–244. [Google Scholar] [CrossRef]
- Allard, P.-M.; Péresse, T.; Bisson, J.; Gindro, K.; Marcourt, L.; Pham, V.C.; Roussi, F.; Litaudon, M.; Wolfender, J.-L. Integration of Molecular Networking and In-Silico MS/MS Fragmentation for Natural Products Dereplication. Anal. Chem. 2016, 88, 3317–3323. [Google Scholar] [CrossRef]
- Essono Mintsa, M.; Otogo N’nang, E.; Choque, É.; Siah, A.; Jacquin, J.; Muchembled, J.; Molinié, R.; Roulard, R.; Cailleu, D.; Beniddir, M.A.; et al. Combined LC-MS/MS and Molecular Networking Approach Reveals Antioxidant and Antimicrobial Compounds from Erismadelphus Exsul Bark. Plants 2022, 11, 1505. [Google Scholar] [CrossRef]
- Lang, G.; Mayhudin, N.A.; Mitova, M.I.; Sun, L.; van der Sar, S.; Blunt, J.W.; Cole, A.L.J.; Ellis, G.; Laatsch, H.; Munro, M.H.G. Evolving Trends in the Dereplication of Natural Product Extracts: New Methodology for Rapid, Small-Scale Investigation of Natural Product Extracts. J. Nat. Prod. 2008, 71, 1595–1599. [Google Scholar] [CrossRef]
- Lučić, D.; Pavlović, I.; Brkljačić, L.; Bogdanović, S.; Farkaš, V.; Cedilak, A.; Nanić, L.; Rubelj, I.; Salopek-Sondi, B. Antioxidant and Antiproliferative Activities of Kale (Brassica Oleracea L. Var. Acephala DC.) and Wild Cabbage (Brassica Incana Ten.) Polyphenolic Extracts. Molecules 2023, 28, 1840. [Google Scholar] [CrossRef]
- Jilani, H.; Cilla, A.; Barberá, R.; Hamdi, M. Antiproliferative Activity of Green, Black Tea and Olive Leaves Polyphenols Subjected to Biosorption and in Vitro Gastrointestinal Digestion in Caco-2 Cells. Food Res. Int. 2020, 136, 109317. [Google Scholar] [CrossRef]
- Huang, D.; Jiang, Y.; Chen, W.; Yao, F.; Sun, L. Polyphenols with Anti-Proliferative Activities from Penthorum Chinense Pursh. Molecules 2014, 19, 11045–11055. [Google Scholar] [CrossRef]
- Yanez, J.; Vicente, V.; Alcaraz, M.; Castillo, J.; Benavente-Garcia, O.; Canteras, M.; Teruel, J.A.L. Cytotoxicity and Antiproliferative Activities of Several Phenolic Compounds Against Three Melanocytes Cell Lines: Relationship Between Structure and Activity. Nutr. Cancer 2004, 49, 191–199. [Google Scholar] [CrossRef]
- Otogo N’Nang, E.; Bernadat, G.; Mouray, E.; Kumulungui, B.; Grellier, P.; Poupon, E.; Champy, P.; Beniddir, M.A. Theionbrunonines A and B: Dimeric Vobasine Alkaloids Tethered by a Thioether Bridge from Mostuea brunonis. Org. Lett. 2018, 20, 6596–6600. [Google Scholar] [CrossRef] [PubMed]
- Chambers, M.C.; Maclean, B.; Burke, R.; Amodei, D.; Ruderman, D.L.; Neumann, S.; Gatto, L.; Fischer, B.; Pratt, B.; Egertson, J.; et al. A Cross-Platform Toolkit for Mass Spectrometry and Proteomics. Nat. Biotechnol. 2012, 30, 918–920. [Google Scholar] [CrossRef]
- Pluskal, T.; Castillo, S.; Villar-Briones, A.; Orešič, M. MZmine 2: Modular Framework for Processing, Visualizing, and Analyzing Mass Spectrometry-Based Molecular Profile Data. BMC Bioinform. 2010, 11, 395. [Google Scholar] [CrossRef] [PubMed]
- Shannon, P. Cytoscape: A Software Environment for Integrated Models of Biomolecular Interaction Networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef]
- Obiang, C.S.; Misso, R.L.N.M.; Atome, G.R.N.; Obame, R.B.M.; Ondo, J.P.; Engonga, L.C.O.; Emvo, E.N. Antimicrobial, Antioxidant, Anti-Inflammatory and Cytotoxic Study of Extracts of Guibourtia Tessmanii (Harms) J. Léonard from Gabon. Clin. Phytosci. 2021, 7, 45. [Google Scholar] [CrossRef]
- Moldovan, C.V.; Savu, M.; Dussert, E.; Aboubacar, H.; Sarbu, L.G.; Matiut, S.; Cudennec, B.; Krier, F.; Ravallec, R.; Birsa, L.M.; et al. Synthetic Flavonoid BrCl-Flav—An Alternative Solution to Combat ESKAPE Pathogens. Antibiotics 2022, 11, 1389. [Google Scholar] [CrossRef]
- Buckingham, J. Dictionary of Natural Products, 1st ed.; Chapman & Hall: London, UK; New York, NY, USA, 1994; ISBN 978-0-412-46620-5. [Google Scholar]
- Baky, M.H.; Kamal, A.M.; Elgindi, M.R.; Haggag, E.G. A Review on Phenolic Compounds from Family Sapotaceae. 8. Available online: https://www.phytojournal.com/archives/2016/vol5issue2/PartD/5-2-15-216.pdf (accessed on 19 February 2023).
- Blaženović, I.; Kind, T.; Ji, J.; Fiehn, O. Software Tools and Approaches for Compound Identification of LC-MS/MS Data in Metabolomics. Metabolites 2018, 8, 31. [Google Scholar] [CrossRef]
- Ernst, M.; Kang, K.B.; Caraballo-Rodríguez, A.M.; Nothias, L.-F.; Wandy, J.; Chen, C.; Wang, M.; Rogers, S.; Medema, M.H.; Dorrestein, P.C.; et al. MolNetEnhancer: Enhanced Molecular Networks by Integrating Metabolome Mining and Annotation Tools. Metabolites 2019, 9, 144. [Google Scholar] [CrossRef]
- Ma, X.; Zheng, C.; Hu, C.; Rahman, K.; Qin, L. The Genus Desmodium (Fabaceae)-Traditional Uses in Chinese Medicine, Phytochemistry and Pharmacology. J. Ethnopharmacol. 2011, 138, 314–332. [Google Scholar] [CrossRef]
- Metsämuuronen, S.; Sirén, H. Bioactive Phenolic Compounds, Metabolism and Properties: A Review on Valuable Chemical Compounds in Scots Pine and Norway Spruce. Phytochem. Rev. 2019, 18, 623–664. [Google Scholar] [CrossRef]
- Ma, J.; Yang, H.; Basile, M.J.; Kennelly, E.J. Analysis of Polyphenolic Antioxidants from the Fruits of Three Pouteria Species by Selected Ion Monitoring Liquid Chromatography−Mass Spectrometry. J. Agric. Food Chem. 2004, 52, 5873–5878. [Google Scholar] [CrossRef] [PubMed]
- Sarfraz, A.; Javeed, M.; Shah, M.A.; Hussain, G.; Shafiq, N.; Sarfraz, I.; Riaz, A.; Sadiqa, A.; Zara, R.; Zafar, S.; et al. Biochanin A: A Novel Bioactive Multifunctional Compound from Nature. Sci. Total Environ. 2020, 722, 137907. [Google Scholar] [CrossRef] [PubMed]
- Paulo, L.; Ferreira, S.; Gallardo, E.; Queiroz, J.A.; Domingues, F. Antimicrobial Activity and Effects of Resveratrol on Human Pathogenic Bacteria. World J. Microbiol. Biotechnol. 2010, 26, 1533–1538. [Google Scholar] [CrossRef]
- Mattio, L.M.; Dallavalle, S.; Musso, L.; Filardi, R.; Franzetti, L.; Pellegrino, L.; D’Incecco, P.; Mora, D.; Pinto, A.; Arioli, S. Antimicrobial Activity of Resveratrol-Derived Monomers and Dimers against Foodborne Pathogens. Sci. Rep. 2019, 9, 19525. [Google Scholar] [CrossRef] [PubMed]
- Demir, T. Effects of Green Tea Powder, Pomegranate Peel Powder, Epicatechin and Punicalagin Additives on Antimicrobial, Antioxidant Potential and Quality Properties of Raw Meatballs. Molecules 2021, 26, 4052. [Google Scholar] [CrossRef]
- Daglia, M. Polyphenols as Antimicrobial Agents. Curr. Opin. Biotechnol. 2012, 23, 174–181. [Google Scholar] [CrossRef]
- Pei, J.; Yu, H.; Qiu, W.; Mei, J.; Xie, J. Antimicrobial Effect of Epigallocatechin Gallate Against Shewanella Putrefaciens ATCC 8071: A Study Based on Cell Membrane and Biofilm. Curr. Microbiol. 2022, 79, 297. [Google Scholar] [CrossRef]
- Wang, J.; Pan, X.; Han, Y.; Guo, D.; Guo, Q.; Li, R. Rosmarinic Acid from Eelgrass Shows Nematicidal and Antibacterial Activities against Pine Wood Nematode and Its Carrying Bacteria. Mar. Drugs 2012, 10, 2729–2740. [Google Scholar] [CrossRef]
- Iqbal, H.; Wright, C.L.; Jones, S.; da Silva, G.R.; McKillen, J.; Gilmore, B.F.; Kavanagh, O.; Green, B.D. Extracts of Sida Cordifolia Contain Polysaccharides Possessing Immunomodulatory Activity and Rosmarinic Acid Compounds with Antibacterial Activity. BMC Complement. Med. 2022, 22, 27. [Google Scholar] [CrossRef] [PubMed]
- Yeo, H.J.; Kwon, M.J.; Han, S.Y.; Jeong, J.C.; Kim, C.Y.; Park, S.U.; Park, C.H. Effects of Carbohydrates on Rosmarinic Acid Production and In Vitro Antimicrobial Activities in Hairy Root Cultures of Agastache Rugosa. Plants 2023, 12, 797. [Google Scholar] [CrossRef]
- Zhang, J.; Cui, X.; Zhang, M.; Bai, B.; Yang, Y.; Fan, S. The Antibacterial Mechanism of Perilla Rosmarinic Acid. Biotechnol. Appl. Biochem. 2022, 69, 1757–1764. [Google Scholar] [CrossRef] [PubMed]
- Jaisinghani, R.N. Antibacterial Properties of Quercetin. Microbiol. Res. 2017, 8, 6877. [Google Scholar] [CrossRef]
- Chittasupho, C.; Manthaisong, A.; Okonogi, S.; Tadtong, S.; Samee, W. Effects of Quercetin and Curcumin Combination on Antibacterial, Antioxidant, In Vitro Wound Healing and Migration of Human Dermal Fibroblast Cells. Int. J. Mol. Sci. 2022, 23, 142. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, T.L.A.; Bhattacharya, D. Antimicrobial Activity of Quercetin: An Approach to Its Mechanistic Principle. Molecules 2022, 27, 2494. [Google Scholar] [CrossRef]
- Podolak, I.; Galanty, A.; Sobolewska, D. Saponins as Cytotoxic Agents: A Review. Phytochem. Rev. 2010, 9, 425–474. [Google Scholar] [CrossRef] [PubMed]
- Kumla, J.; Suwannarach, N.; Tanruean, K.; Lumyong, S. Comparative Evaluation of Chemical Composition, Phenolic Compounds, and Antioxidant and Antimicrobial Activities of Tropical Black Bolete Mushroom Using Different Preservation Methods. Foods 2021, 10, 781. [Google Scholar] [CrossRef] [PubMed]
- Csepregi, R.; Temesfői, V.; Das, S.; Alberti, Á.; Tóth, C.A.; Herczeg, R.; Papp, N.; Kőszegi, T. Cytotoxic, Antimicrobial, Antioxidant Properties and Effects on Cell Migration of Phenolic Compounds of Selected Transylvanian Medicinal Plants. Antioxidants 2020, 9, 166. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Corona, A.V.; Valencia-Espinosa, I.; González-Sánchez, F.A.; Sánchez-López, A.L.; Garcia-Amezquita, L.E.; Garcia-Varela, R. Antioxidant, Anti-Inflammatory and Cytotoxic Activity of Phenolic Compound Family Extracted from Raspberries (Rubus Idaeus): A General Review. Antioxidants 2022, 11, 1192. [Google Scholar] [CrossRef]
- Xie, P.; Cecchi, L.; Bellumori, M.; Balli, D.; Giovannelli, L.; Huang, L.; Mulinacci, N. Phenolic Compounds and Triterpenes in Different Olive Tissues and Olive Oil By-Products, and Cytotoxicity on Human Colorectal Cancer Cells: The Case of Frantoio, Moraiolo and Leccino Cultivars (Olea Europaea L.). Foods 2021, 10, 2823. [Google Scholar] [CrossRef]
- Hsu, Y.-N.; Shyu, H.-W.; Hu, T.-W.; Yeh, J.-P.; Lin, Y.-W.; Lee, L.-Y.; Yeh, Y.-T.; Dai, H.-Y.; Perng, D.-S.; Su, S.-H.; et al. Anti-Proliferative Activity of Biochanin A in Human Osteosarcoma Cells via Mitochondrial-Involved Apoptosis. Food Chem. Toxicol. 2018, 112, 194–204. [Google Scholar] [CrossRef]
- Kole, L.; Giri, B.; Manna, S.K.; Pal, B.; Ghosh, S. Biochanin-A, an Isoflavon, Showed Anti-Proliferative and Anti-Inflammatory Activities through the Inhibition of INOS Expression, P38-MAPK and ATF-2 Phosphorylation and Blocking NFκB Nuclear Translocation. Eur. J. Pharmacol. 2011, 653, 8–15. [Google Scholar] [CrossRef] [PubMed]
- Storniolo, C.E.; Moreno, J.J. Resveratrol Analogs with Antioxidant Activity Inhibit Intestinal Epithelial Cancer Caco-2 Cell Growth by Modulating Arachidonic Acid Cascade. J. Agric. Food Chem. 2019, 67, 819–828. [Google Scholar] [CrossRef]
- Aboushanab, S.A.; Shevyrin, V.A.; Melekhin, V.V.; Andreeva, E.I.; Makeev, O.G.; Kovaleva, E.G. Cytotoxic Activity and Phytochemical Screening of Eco-Friendly Extracted Flavonoids from Pueraria Montana Var. Lobata (Willd.) Sanjappa & Pradeep and Trifolium Pratense L. Flowers Using HPLC-DAD-MS/HRMS. AppliedChem 2023, 3, 119–140. [Google Scholar] [CrossRef]
- Gurgul, A.; Nauman, M.C.; Wu, Z.; Shetye, G.; Ma, R.; Youn, I.; Souliya, O.; Bisson, J.; Johnson, J.J.; Che, C.-T. Chemical Constituents of the Stem of Marsypopetalum Modestum and Their Bioactivities. Nat. Prod. Res. 2022, 2022, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Yesil-Celiktas, O.; Sevimli, C.; Bedir, E.; Vardar-Sukan, F. Inhibitory Effects of Rosemary Extracts, Carnosic Acid and Rosmarinic Acid on the Growth of Various Human Cancer Cell Lines. Plant Foods Hum. Nutr. 2010, 65, 158–163. [Google Scholar] [CrossRef] [PubMed]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).