
Deciphering Gut Microbiome Responses upon Microplastic Exposure via Integrating Metagenomics and Activity-Based Metabolomics

 ,
,  , , ,
, , , 
Abstract
1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Animals and Experimental Design
2.3. 16S rRNA Gene Sequencing
2.4. Quantification of Fecal Short-Chain Fatty Acids (SCFAs)
2.5. Untargeted Metabolomic Analysis
2.6. Statistical Analysis
3. Results
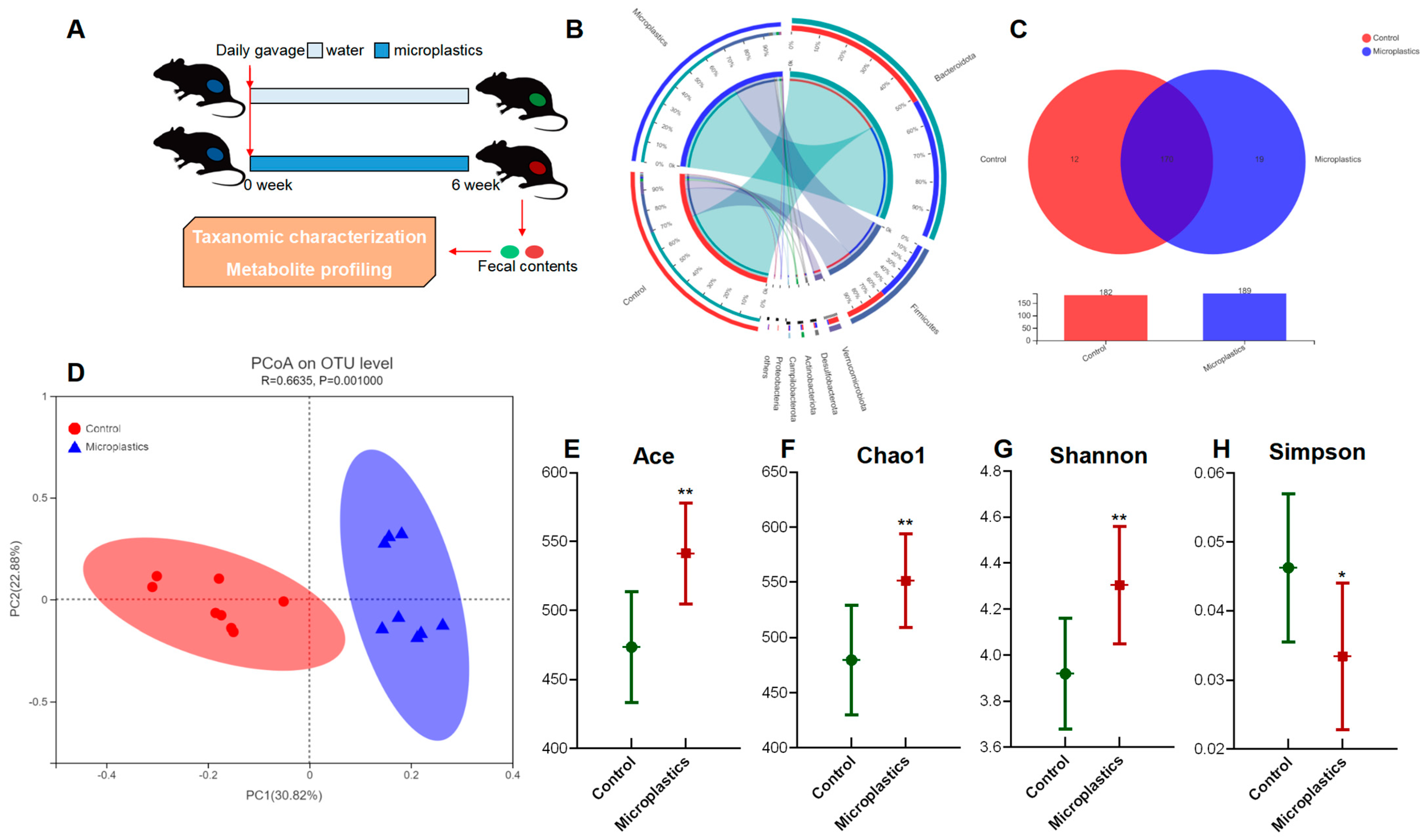
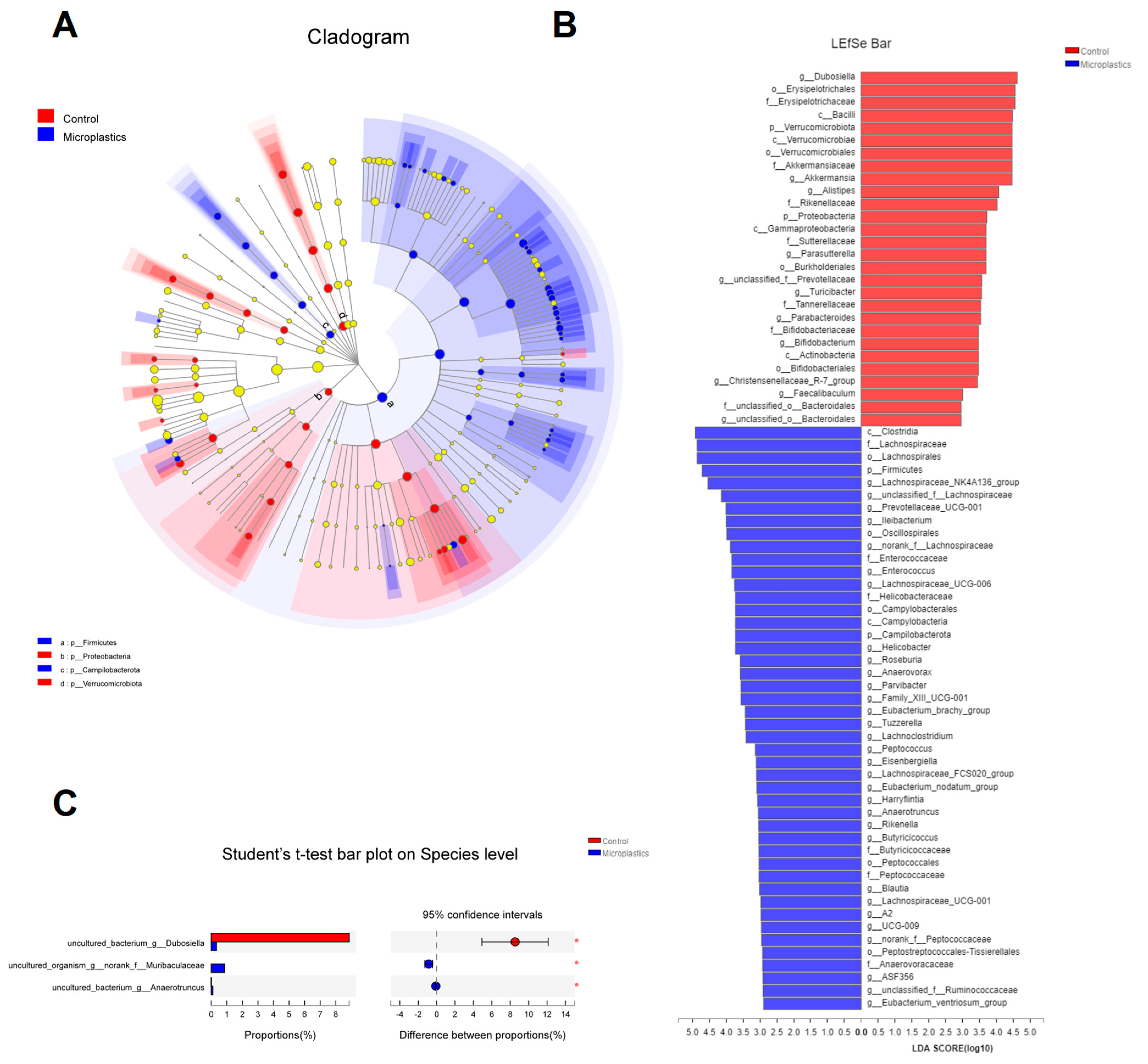
3.1. MP-Induced Alterations in Diversity and Composition of the Gut Microbial Community
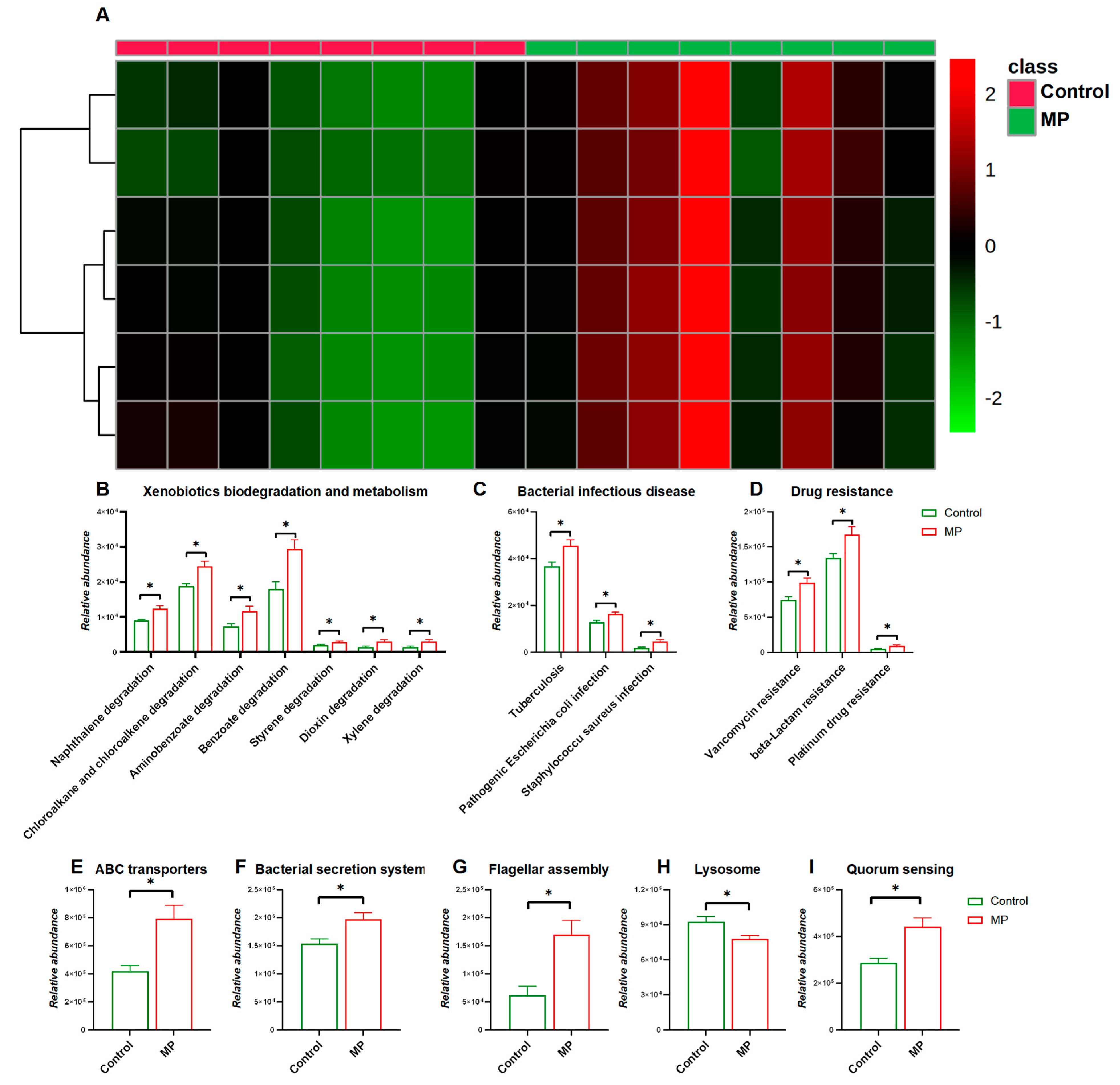
3.2. MP-Induced Changes in Functional Pathways of the Gut Microbiome
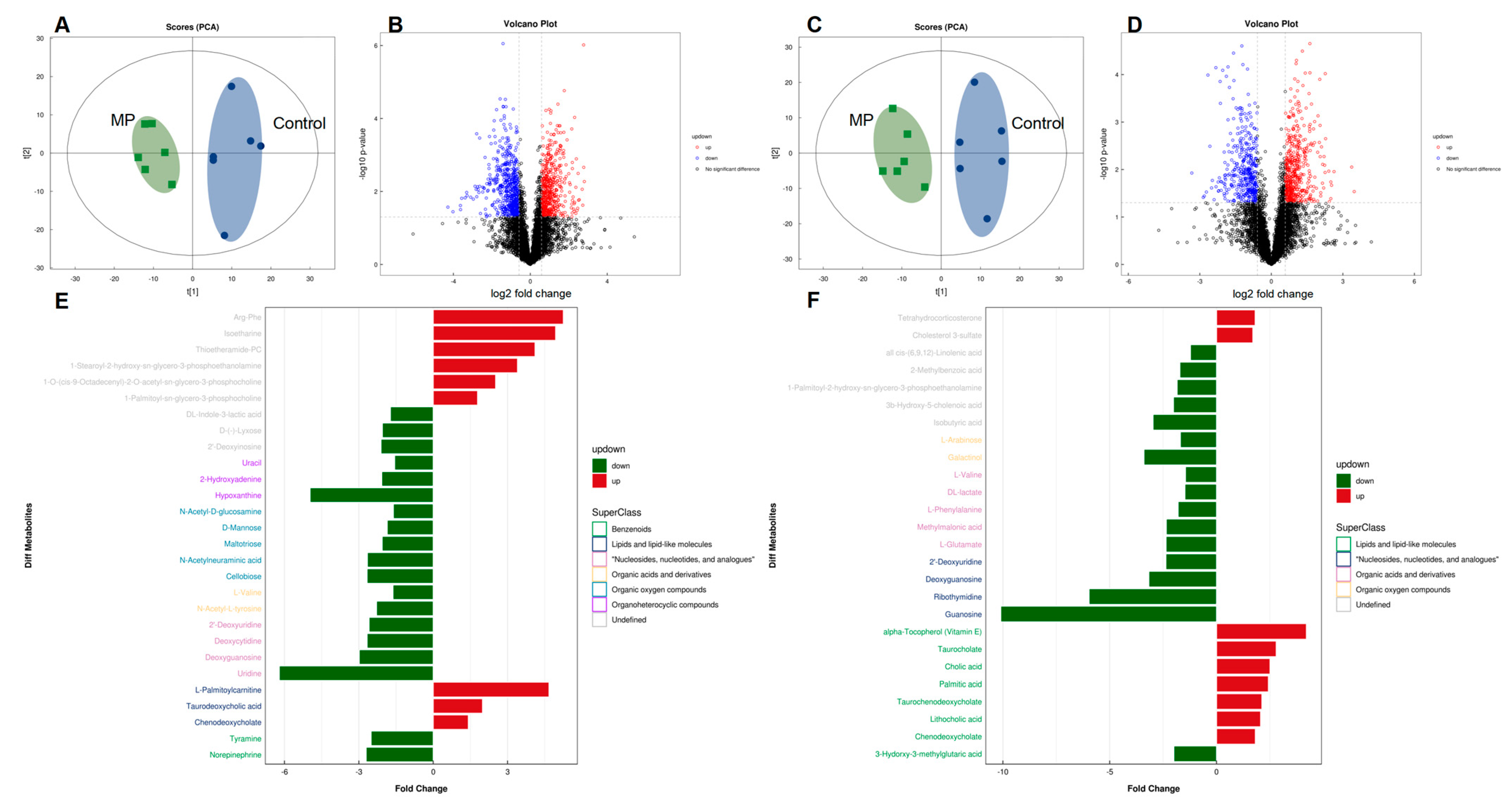
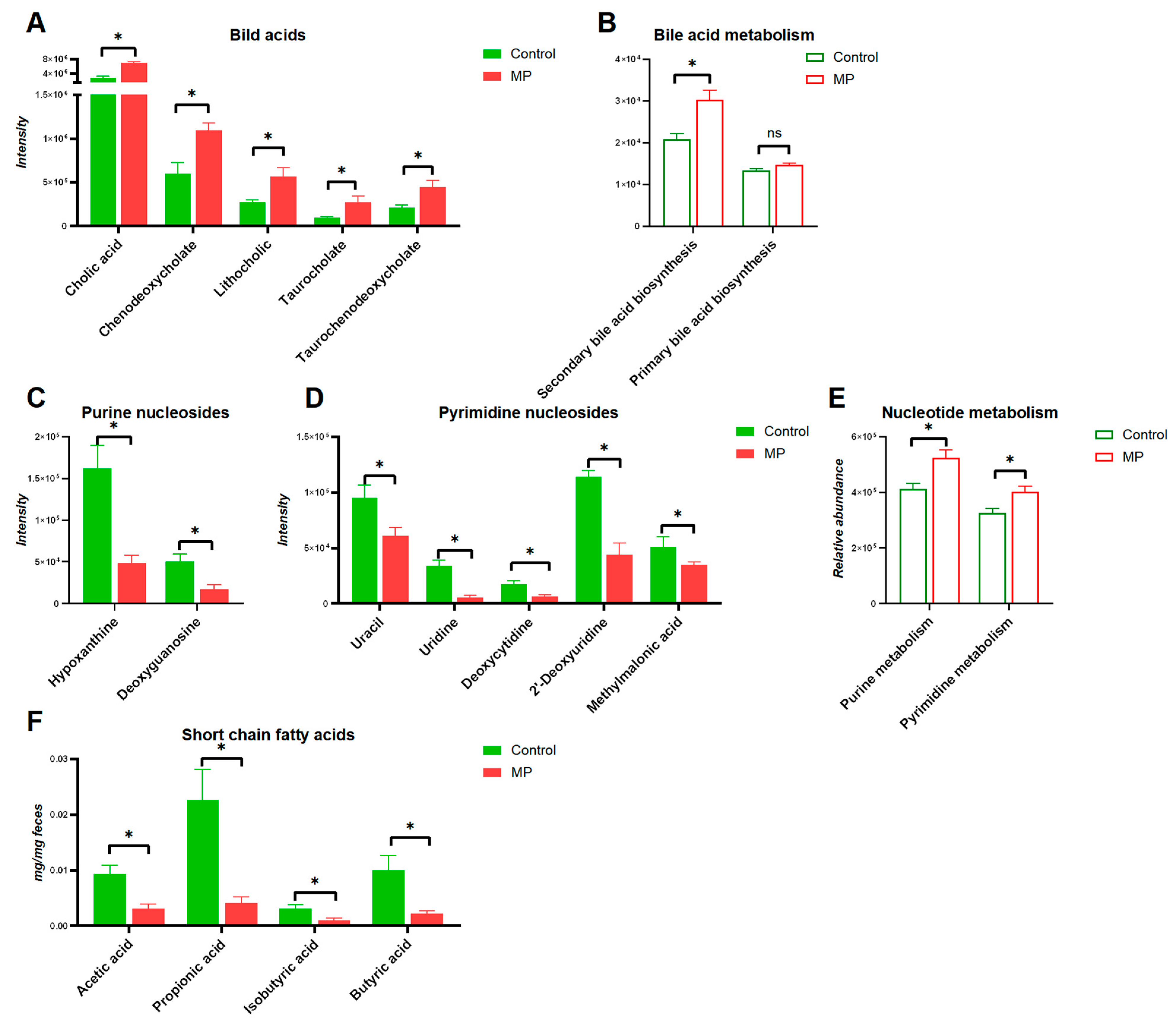
3.3. MP Alters Metabolite Profiles of the Gut Microbiome
3.4. Key Metabolites That Are Associated with MP Exposure
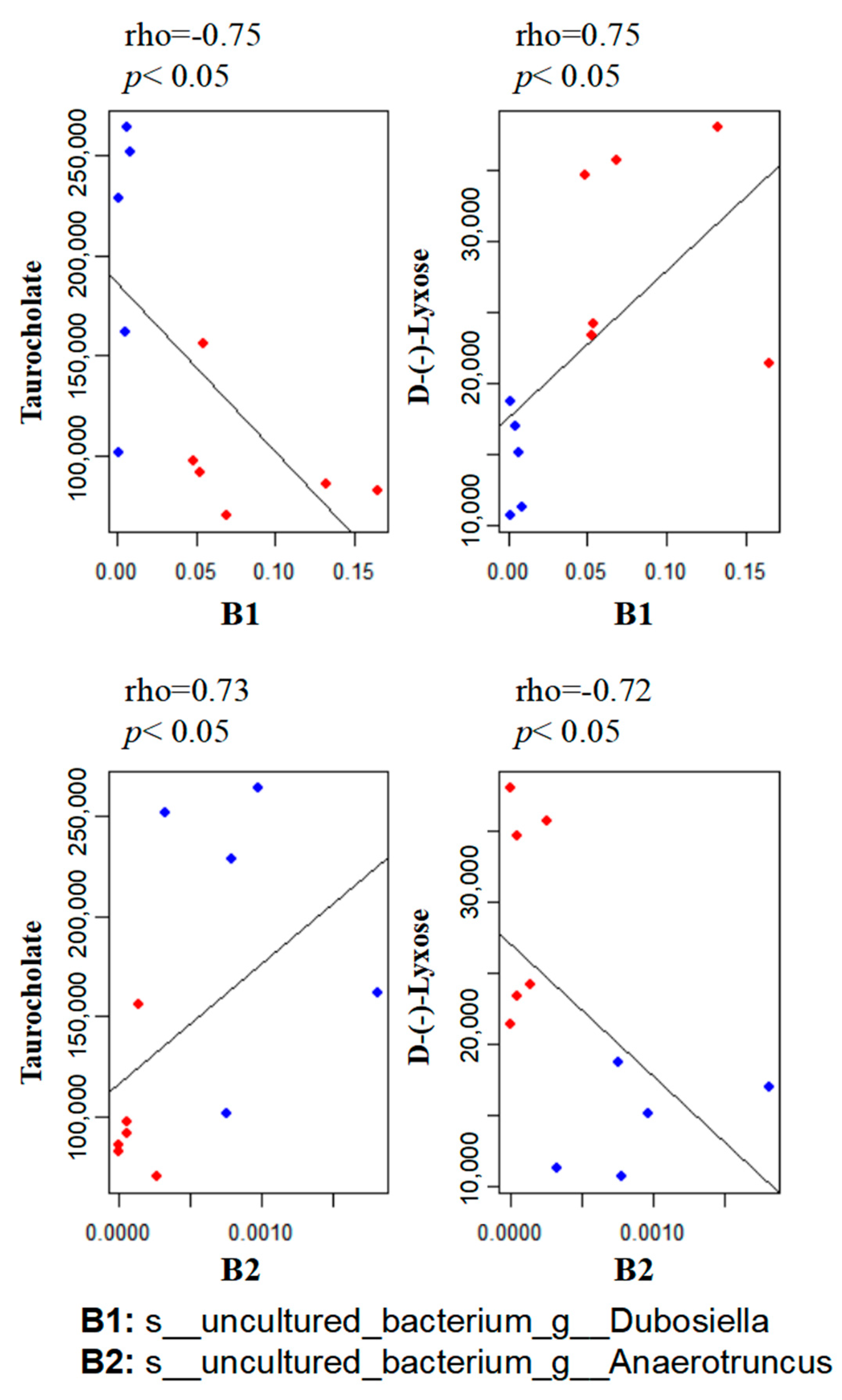
3.5. Correlation between the Gut Microbiome and Metabolites
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Pickard, J.M.; Zeng, M.Y.; Caruso, R.; Núñez, G. Gut microbiota: Role in pathogen colonization, immune responses, and inflammatory disease. Immunol. Rev. 2017, 279, 70–89. [Google Scholar] [CrossRef] [PubMed]
- Round, J.L.; Mazmanian, S.K. The gut microbiota shapes intestinal immune responses during health and disease. Nat. Rev. Immunol. 2009, 9, 313–323. [Google Scholar] [CrossRef] [PubMed]
- Tremaroli, V.; Bäckhed, F. Functional interactions between the gut microbiota and host metabolism. Nature 2012, 489, 242–249. [Google Scholar] [CrossRef] [PubMed]
- Shi, N.; Li, N.; Duan, X.; Niu, H. Interaction between the gut microbiome and mucosal immune system. Mil. Med. Res. 2017, 4, 14. [Google Scholar] [CrossRef]
- Clemente, J.C.; Manasson, J.; Scher, J.U. The role of the gut microbiome in systemic inflammatory disease. BMJ 2018, 360, j5145. [Google Scholar] [CrossRef]
- Turnbaugh, P.J.; Ley, R.E.; Mahowald, M.A.; Magrini, V.; Mardis, E.R.; Gordon, J.I. An obesity-associated gut microbiome with increased capacity for energy harvest. Nature 2006, 444, 1027–1031. [Google Scholar] [CrossRef]
- Gurung, M.; Li, Z.; You, H.; Rodrigues, R.; Jump, D.B.; Morgun, A.; Shulzhenko, N. Role of gut microbiota in type 2 diabetes pathophysiology. EBioMedicine 2020, 51, 102590. [Google Scholar] [CrossRef]
- Zheng, P.; Li, Z.; Zhou, Z. Gut microbiome in type 1 diabetes: A comprehensive review. Diabetes Metab. Res. Rev. 2018, 34, e3043. [Google Scholar] [CrossRef]
- Louis, P.; Hold, G.L.; Flint, H.J. The gut microbiota, bacterial metabolites and colorectal cancer. Nat. Rev. Microbiol. 2014, 12, 661–672. [Google Scholar] [CrossRef]
- Chi, L.; Bian, X.; Gao, B.; Ru, H.; Tu, P.; Lu, K. Sex-Specific Effects of Arsenic Exposure on the Trajectory and Function of the Gut Microbiome. Chem. Res. Toxicol. 2016, 29, 949–951. [Google Scholar] [CrossRef]
- Chi, L.; Gao, B.; Bian, X.; Tu, P.; Ru, H.; Lu, K. Manganese-induced sex-specific gut microbiome perturbations in C57BL/6 mice. Toxicol. Appl. Pharmacol. 2017, 331, 142–153. [Google Scholar] [CrossRef] [PubMed]
- Gao, B.; Chi, L.; Mahbub, R.; Bian, X.; Tu, P.; Ru, H.; Lu, K. Multi-Omics Reveals that Lead Exposure Disturbs Gut Microbiome Development, Key Metabolites, and Metabolic Pathways. Chem. Res. Toxicol. 2017, 30, 996–1005. [Google Scholar] [CrossRef] [PubMed]
- Lu, K.; Abo, R.P.; Schlieper, K.A.; Graffam, M.E.; Levine, S.; Wishnok, J.S.; Swenberg, J.A.; Tannenbaum, S.R.; Fox, J.G. Arsenic exposure perturbs the gut microbiome and its metabolic profile in mice: An integrated metagenomics and metabolomics analysis. Environ. Health Perspect 2014, 122, 284–291. [Google Scholar] [CrossRef] [PubMed]
- Gao, B.; Bian, X.; Chi, L.; Tu, P.; Ru, H.; Lu, K. Editor’s Highlight: OrganophosphateDiazinon Altered Quorum Sensing, Cell Motility, Stress Response, and Carbohydrate Metabolism of Gut Microbiome. Toxicol. Sci. 2017, 157, 354–364. [Google Scholar] [CrossRef] [PubMed]
- Bian, X.; Tu, P.; Chi, L.; Gao, B.; Ru, H.; Lu, K. Saccharin induced liver inflammation in mice by altering the gut microbiota and its metabolic functions. Food Chem. Toxicol. 2017, 107, 530–539. [Google Scholar] [CrossRef]
- Bian, X.; Chi, L.; Gao, B.; Tu, P.; Ru, H.; Lu, K. The artificial sweetener acesulfame potassium affects the gut microbiome and body weight gain in CD-1 mice. PLoS ONE 2017, 12, e0178426. [Google Scholar] [CrossRef]
- Bian, X.; Chi, L.; Gao, B.; Tu, P.; Ru, H.; Lu, K. Gut Microbiome Response to Sucralose and Its Potential Role in Inducing Liver Inflammation in Mice. Front. Physiol. 2017, 8, 487. [Google Scholar] [CrossRef]
- Tu, P.; Chi, L.; Bodnar, W.; Zhang, Z.; Gao, B.; Bian, X.; Stewart, J.; Fry, R.; Lu, K. Gut Microbiome Toxicity: Connecting the Environment and Gut Microbiome-Associated Diseases. Toxics 2020, 8, 19. [Google Scholar] [CrossRef]
- Chi, L.; Bian, X.; Gao, B.; Tu, P.; Ru, H.; Lu, K. The Effects of an Environmentally Relevant Level of Arsenic on the Gut Microbiome and Its Functional Metagenome. Toxicol. Sci. 2017, 160, 193–204. [Google Scholar] [CrossRef]
- Chi, L.; Gao, B.; Tu, P.; Liu, C.-W.; Xue, J.; Lai, Y.; Ru, H.; Lu, K. Individual susceptibility to arsenic-induced diseases: The role of host genetics, nutritional status, and the gut microbiome. Mamm. Genome 2018, 29, 63–79. [Google Scholar] [CrossRef]
- Chi, L.; Mahbub, R.; Gao, B.; Bian, X.; Tu, P.; Ru, H.; Lu, K. Nicotine Alters the Gut Microbiome and Metabolites of Gut-Brain Interactions in a Sex-Specific Manner. Chem. Res. Toxicol. 2017, 30, 2110–2119. [Google Scholar] [CrossRef] [PubMed]
- Toussaint, B.; Raffael, B.; Angers-Loustau, A.; Gilliland, D.; Kestens, V.; Petrillo, M.; Rio-Echevarria, I.M.; Van den Eede, G. Review of micro- and nanoplastic contamination in the food chain. Food Addit. Contam. Part A 2019, 36, 639–673. [Google Scholar] [CrossRef] [PubMed]
- Prata, J.C.; da Costa, J.P.; Lopes, I.; Duarte, A.C.; Rocha-Santos, T. Environmental exposure to microplastics: An overview on possible human health effects. Sci. Total Environ. 2020, 702, 134455. [Google Scholar] [CrossRef] [PubMed]
- Vandenberg, L.N.; Hauser, R.; Marcus, M.; Olea, N.; Welshons, W.V. Human exposure to bisphenol A (BPA). Reprod. Toxicol. 2007, 24, 139–177. [Google Scholar] [CrossRef]
- Li, B.; Ding, Y.; Cheng, X.; Sheng, D.; Xu, Z.; Rong, Q.; Wu, Y.; Zhao, H.; Ji, X.; Zhang, Y. Polyethylene microplastics affect the distribution of gut microbiota and inflammation development in mice. Chemosphere 2020, 244, 125492. [Google Scholar] [CrossRef]
- Lu, L.; Wan, Z.; Luo, T.; Fu, Z.; Jin, Y. Polystyrene microplastics induce gut microbiota dysbiosis and hepatic lipid metabolism disorder in mice. Sci. Total Environ. 2018, 631–632, 449–458. [Google Scholar] [CrossRef]
- Jin, Y.; Lu, L.; Tu, W.; Luo, T.; Fu, Z. Impacts of polystyrene microplastic on the gut barrier, microbiota and metabolism of mice. Sci. Total Environ. 2019, 649, 308–317. [Google Scholar] [CrossRef]
- Jiang, P.; Yuan, G.-H.; Jiang, B.-R.; Zhang, J.-Y.; Wang, Y.-Q.; Lv, H.-J.; Zhang, Z.; Wu, J.-L.; Wu, Q.; Li, L. Effects of microplastics (MPs) and tributyltin (TBT) alone and in combination on bile acids and gut microbiota crosstalk in mice. Ecotoxicol. Environ. Saf. 2021, 220, 112345. [Google Scholar] [CrossRef]
- Claus, S.P.; Ellero, S.L.; Berger, B.; Krause, L.; Bruttin, A.; Molina, J.; Paris, A.; Want, E.J.; de Waziers, I.; Cloarec, O.; et al. Colonization-induced host-gut microbial metabolic interaction. MBio 2011, 2, e00271-10. [Google Scholar] [CrossRef]
- Diaz Heijtz, R.; Wang, S.; Anuar, F.; Qian, Y.; Björkholm, B.; Samuelsson, A.; Hibberd, M.L.; Forssberg, H.; Pettersson, S. Normal gut microbiota modulates brain development and behavior. Proc. Natl. Acad. Sci. USA 2011, 108, 3047–3052. [Google Scholar] [CrossRef]
- Chen, W.; Zhu, R.; Ye, X.; Sun, Y.; Tang, Q.; Liu, Y.; Yan, F.; Yu, T.; Zheng, X.; Tu, P. Food-derived cyanidin-3-O-glucoside reverses microplastic toxicity promoting discharge and modulating the gut microbiota in mice. Food Funct. 2022, 13, 1447–1458. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Ye, X.; Tang, Q.; Yu, T.; Tu, P.; Zheng, X. Cyanidin-3-O-glucoside reduces nanopolystyrene-induced toxicity and accumulation: Roles of mitochondrial energy metabolism and cellular efflux. Environ. Sci. Nano 2022, 7, 2572–2586. [Google Scholar] [CrossRef]
- Kik, K.; Bukowska, B.; Sicińska, P. Polystyrene nanoparticles: Sources, occurrence in the environment, distribution in tissues, accumulation and toxicity to various organisms. Environ. Pollut. 2020, 262, 114297. [Google Scholar] [CrossRef] [PubMed]
- Deng, Y.; Zhang, Y.; Lemos, B.; Ren, H. Tissue accumulation of microplastics in mice and biomarker responses suggest widespread health risks of exposure. Sci. Rep. 2017, 7, 46687. [Google Scholar] [CrossRef]
- Watts, A.J.R.; Lewis, C.; Goodhead, R.M.; Beckett, S.J.; Moger, J.; Tyler, C.R.; Galloway, T.S. Uptake and retention of microplastics by the shore crab Carcinus maenas. Environ. Sci. Technol. 2014, 48, 8823–8830. [Google Scholar] [CrossRef]
- Hämer, J.; Gutow, L.; Köhler, A.; Saborowski, R. Fate of microplastics in the marine isopod Idotea emarginata. Environ. Sci. Technol. 2014, 48, 13451–13458. [Google Scholar] [CrossRef]
- Eerkes-Medrano, D.; Thompson, R.C.; Aldridge, D.C. Microplastics in freshwater systems: A review of the emerging threats, identification of knowledge gaps and prioritisation of research needs. Water Res. 2015, 75, 63–82. [Google Scholar] [CrossRef]
- Aßhauer, K.P.; Wemheuer, B.; Daniel, R.; Meinicke, P. Tax4Fun: Predicting functional profiles from metagenomic 16S rRNA data. Bioinformatics 2015, 31, 2882–2884. [Google Scholar] [CrossRef]
- Zheng, X.; Qiu, Y.; Zhong, W.; Baxter, S.; Su, M.; Li, Q.; Xie, G.; Ore, B.M.; Qiao, S.; Spencer, M.D.; et al. A targeted metabolomic protocol for short-chain fatty acids and branched-chain amino acids. Metabolomics 2013, 9, 818–827. [Google Scholar] [CrossRef]
- Gomaa, E.Z. Human gut microbiota/microbiome in health and diseases: A review. Antonie Van Leeuwenhoek 2020, 113, 2019–2040. [Google Scholar] [CrossRef]
- Patterson, E.; Ryan, P.M.; Cryan, J.F.; Dinan, T.G.; Ross, R.P.; Fitzgerald, G.F.; Stanton, C. Gut microbiota, obesity and diabetes. Postgrad. Med. J. 2016, 92, 286–300. [Google Scholar] [CrossRef]
- Scheithauer, T.P.M.; Rampanelli, E.; Nieuwdorp, M.; Vallance, B.A.; Verchere, C.B.; van Raalte, D.H.; Herrema, H. Gut Microbiota as a Trigger for Metabolic Inflammation in Obesity and Type 2 Diabetes. Front. Immunol. 2020, 11, 571731. [Google Scholar] [CrossRef] [PubMed]
- Wu, B.; Wu, X.; Liu, S.; Wang, Z.; Chen, L. Size-dependent effects of polystyrene microplastics on cytotoxicity and efflux pump inhibition in human Caco-2 cells. Chemosphere 2019, 221, 333–341. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, A.; Bouscarel, B. Bile acids and signal transduction: Role in glucose homeostasis. Cell. Signal. 2008, 20, 2180–2197. [Google Scholar] [CrossRef] [PubMed]
- Swann, J.R.; Want, E.J.; Geier, F.M.; Spagou, K.; Wilson, I.D.; Sidaway, J.E.; Nicholson, J.K.; Holmes, E. Systemic gut microbial modulation of bile acid metabolism in host tissue compartments. Proc. Natl. Acad. Sci. USA 2011, 108 (Suppl. 1), 4523–4530. [Google Scholar] [CrossRef] [PubMed]
- Brestoff, J.R.; Artis, D. Commensal bacteria at the interface of host metabolism and the immune system. Nat. Immunol. 2013, 14, 676–684. [Google Scholar] [CrossRef]
- Jing, J.; Zhang, L.; Han, L.; Wang, J.; Zhang, W.; Liu, Z.; Gao, A. Polystyrene micro-/nanoplastics induced hematopoietic damages via the crosstalk of gut microbiota, metabolites, and cytokines. Environ. Int. 2022, 161, 107131. [Google Scholar] [CrossRef]
- Lührs, H.; Gerke, T.; Müller, J.G.; Melcher, R.; Schauber, J.; Boxberge, F.; Scheppach, W.; Menzel, T. Butyrate inhibits NF-kappaB activation in lamina propria macrophages of patients with ulcerative colitis. Scand. J. Gastroenterol. 2002, 37, 458–466. [Google Scholar] [CrossRef]
- Maeda, T.; Towatari, M.; Kosugi, H.; Saito, H. Up-regulation of costimulatory/adhesion molecules by histone deacetylase inhibitors in acute myeloid leukemia cells. Blood 2000, 96, 3847–3856. [Google Scholar] [CrossRef]
- Arpaia, N.; Campbell, C.; Fan, X.; Dikiy, S.; van der Veeken, J.; deRoos, P.; Liu, H.; Cross, J.R.; Pfeffer, K.; Coffer, P.J.; et al. Metabolites produced by commensal bacteria promote peripheral regulatory T-cell generation. Nature 2013, 504, 451–455. [Google Scholar] [CrossRef]
- Furusawa, Y.; Obata, Y.; Fukuda, S.; Endo, T.A.; Nakato, G.; Takahashi, D.; Nakanishi, Y.; Uetake, C.; Kato, K.; Kato, T.; et al. Commensal microbe-derived butyrate induces the differentiation of colonic regulatory T cells. Nature 2013, 504, 446–450. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Annotated Feature | Adduct | m/z | rt(s) a | VIP b | Fold Change c | p-Value | HMDB ID |
|---|---|---|---|---|---|---|---|
| Nucleosides, nucleotides, and analogues | |||||||
| 2’-Deoxyuridine | (M − H)− | 227.0671 | 112.8 | 10.7 | 0.4 | 6.80 × 10−4 | 12 |
| Deoxycytidine | (M + H)+ | 228.0964 | 206.6 | 1 | 0.4 | 7.24 × 10−3 | 14 |
| Deoxyguanosine | (M − H)− | 266.0890 | 230.3 | 2.1 | 0.3 | 6.30 × 10−3 | 85 |
| Guanosine | (M − H)− | 282.0840 | 262.8 | 1.3 | 0.1 | 1.20 × 10−2 | 133 |
| Uridine | (M + H)+ | 245.0757 | 159.2 | 1.7 | 0.2 | 3.44 × 10−4 | 296 |
| Ribothymidine | (M − H)− | 257.0780 | 142 | 4 | 0.2 | 3.20 × 10−3 | 884 |
| Lipids and lipid-like molecules | |||||||
| 3-Hydorxy−3-methylglutaric acid | (M − H)− | 161.0452 | 373.8 | 1.1 | 0.5 | 9.00 × 10−4 | 355 |
| alpha-Tocopherol (Vitamin E) | (M − H)− | 429.3724 | 31.5 | 1.9 | 4.2 | 3.10 × 10−2 | 1893 |
| Cholic acid | (M − H)− | 407.2802 | 227.2 | 14 | 2.5 | 1.70 × 10−4 | 619 |
| Palmitic acid | (M − H)− | 255.2327 | 46.7 | 9.4 | 2.4 | 4.60 × 10−2 | 220 |
| Taurochenodeoxycholate | (M − H)− | 498.2886 | 140.7 | 3 | 2.1 | 1.60 × 10−2 | 951 |
| Taurocholate | (M − H)− | 514.2840 | 200.3 | 2.4 | 2.8 | 4.40 × 10−2 | 36 |
| Chenodeoxycholate | (M + CH3COO)− | 451.3053 | 160.3 | 4.4 | 1.8 | 8.00 × 10−3 | 518 |
| Lithocholic acid | (M + CH3COO)− | 435.3105 | 82.6 | 3.3 | 2.1 | 2.10 × 10−2 | 761 |
| L-Palmitoylcarnitine | (M + H)+ | 400.3401 | 172.9 | 2.4 | 4.7 | 1.41 × 10−2 | 222 |
| Taurodeoxycholic acid | (M + NH4)+ | 517.3270 | 140.8 | 1.7 | 2 | 1.01 × 10−2 | 896 |
| Organic acids and derivatives | |||||||
| DL-lactate | (M − H)− | 89.0243 | 304.1 | 1.8 | 0.7 | 4.00 × 10−2 | 1311 |
| L-Glutamate | (M − H)− | 146.0458 | 398.3 | 2 | 0.4 | 4.70 × 10−2 | 148 |
| L-Phenylalanine | (M − H)− | 164.0719 | 261.5 | 3.1 | 0.6 | 5.00 × 10−3 | 159 |
| L-Valine | (M − H)− | 116.0714 | 304.9 | 2.3 | 0.7 | 2.50 × 10−2 | 883 |
| Methylmalonic acid | (M − H)− | 117.0188 | 104.9 | 1.2 | 0.4 | 1.10 × 10−2 | 202 |
| L-Arabinose | (M − H)− | 149.0449 | 133 | 2 | 0.6 | 1.50 × 10−2 | 646 |
| Galactinol | (M + CH3COO)− | 401.1292 | 391.2 | 1.1 | 0.3 | 1.20 × 10−2 | 5826 |
| N-Acetyl-L-tyrosine | (M − H + 2Na)+ | 268.0606 | 242.4 | 1.3 | 0.4 | 3.45 × 10−2 | 866 |
| N-Acetyl-D-glucosamine | (M + H)+ | 222.0966 | 256.4 | 3 | 0.6 | 2.76 × 10−2 | 215 |
| N-Acetylneuraminic acid | (M + H)+ | 310.1121 | 373.6 | 2.2 | 0.4 | 4.82 × 10−5 | 230 |
| Cellobiose | (M + NH4)+ | 360.1487 | 389.6 | 4.4 | 0.4 | 3.05 × 10−2 | 55 |
| D-Mannose | (M + NH4)+ | 198.0958 | 302.8 | 2 | 0.5 | 4.46 × 10−3 | 169 |
| Maltotriose | (M + NH4)+ | 522.2001 | 449.6 | 1.6 | 0.5 | 2.10 × 10−2 | 1262 |
| Benzenoids | |||||||
| Tyramine | (M + H)+ | 138.0900 | 218.2 | 1.2 | 0.4 | 2.46 × 10−3 | 306 |
| Norepinephrine | (M + H − H2O)+ | 152.0691 | 105.2 | 2.7 | 0.4 | 3.43 × 10−4 | 216 |
| Organoheterocyclic compounds | |||||||
| 2-Hydroxyadenine | (M + H)+ | 152.0559 | 262.4 | 1.5 | 0.5 | 3.03 × 10−2 | 403 |
| Hypoxanthine | (M + H)+ | 137.0448 | 217.3 | 4.2 | 0.2 | 4.17 × 10−2 | 157 |
| Uracil | (M + H)+ | 113.0334 | 84.7 | 1.9 | 0.6 | 3.31 × 10−2 | 300 |
| Others | |||||||
| 1-Palmitoyl−2-hydroxy-sn-glycero−3-phosphoethanolamine | (M − H)− | 452.2773 | 200.6 | 4.6 | 0.5 | 2.00 × 10−2 | n.a.d |
| 2-Methylbenzoic acid | (M − H)− | 135.0445 | 133.3 | 1.2 | 0.6 | 1.20 × 10−2 | 2340 |
| 3b-Hydroxy−5-cholenoic acid | (M − H)− | 373.2733 | 61.2 | 2.5 | 0.5 | 3.00 × 10−2 | 308 |
| gamma-Linolenic acid | (M − H)− | 277.2174 | 46.1 | 6.4 | 0.8 | 4.20 × 10−2 | 3073 |
| Cholesterol 3-sulfate | (M − H)− | 465.3042 | 26.3 | 15.8 | 1.7 | 1.40 × 10−2 | 653 |
| Isobutyric acid | (M − H)− | 87.0452 | 132.6 | 6 | 0.3 | 1.30 × 10−4 | 1873 |
| Tetrahydrocorticosterone | (M − H)− | 349.2373 | 67.6 | 1.4 | 1.8 | 7.50 × 10−3 | 268 |
| Isoetharine | (M + CH3CN + Na)+ | 303.1689 | 197.6 | 1.6 | 4.9 | 2.00 ×10−2 | 14366 |
| 1-Palmitoyl-sn-glycero−3-phosphocholine | (M + H)+ | 496.3396 | 194.5 | 10.3 | 1.8 | 3.93 ×10−3 | n.a. |
| 1-Stearoyl−2-hydroxy-sn-glycero−3-phosphoethanolamine | (M + H)+ | 482.3230 | 195.7 | 3.1 | 3.4 | 1.73 ×10−5 | n.a. |
| 2’-Deoxyinosine | (M + H)+ | 253.0924 | 179.6 | 3.5 | 0.5 | 3.21 ×10−2 | 71 |
| Arg-Phe | (M + H)+ | 322.1851 | 340.8 | 1.2 | 5.3 | 1.28 ×10−3 | n.a. |
| DL-Indole−3-lactic acid | (M + H − H2O)+ | 188.0693 | 259.1 | 1.3 | 0.6 | 3.25 ×10−2 | 671 |
| Thioetheramide-PC | (M + Na)+ | 758.5646 | 129.4 | 2.1 | 4.1 | 5.77 ×10−3 | n.a. |
| D-(-)-Lyxose | (M + NH4)+ | 168.0853 | 152.6 | 1.3 | 0.5 | 9.26 ×10−4 | n.a. |
| 1-O-(cis−9-Octadecenyl)−2-O-acetyl-sn-glycero−3-phosphocholine | (M + H)+ | 550.3832 | 188.7 | 1.7 | 2.5 | 4.49 ×10−4 | n.a. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tu, P.; Xue, J.; Niu, H.; Tang, Q.; Mo, Z.; Zheng, X.; Wu, L.; Chen, Z.; Cai, Y.; Wang, X. Deciphering Gut Microbiome Responses upon Microplastic Exposure via Integrating Metagenomics and Activity-Based Metabolomics. Metabolites 2023, 13, 530. https://doi.org/10.3390/metabo13040530
Tu P, Xue J, Niu H, Tang Q, Mo Z, Zheng X, Wu L, Chen Z, Cai Y, Wang X. Deciphering Gut Microbiome Responses upon Microplastic Exposure via Integrating Metagenomics and Activity-Based Metabolomics. Metabolites. 2023; 13(4):530. https://doi.org/10.3390/metabo13040530
Chicago/Turabian StyleTu, Pengcheng, Jingchuan Xue, Huixia Niu, Qiong Tang, Zhe Mo, Xiaodong Zheng, Lizhi Wu, Zhijian Chen, Yanpeng Cai, and Xiaofeng Wang. 2023. "Deciphering Gut Microbiome Responses upon Microplastic Exposure via Integrating Metagenomics and Activity-Based Metabolomics" Metabolites 13, no. 4: 530. https://doi.org/10.3390/metabo13040530
APA StyleTu, P., Xue, J., Niu, H., Tang, Q., Mo, Z., Zheng, X., Wu, L., Chen, Z., Cai, Y., & Wang, X. (2023). Deciphering Gut Microbiome Responses upon Microplastic Exposure via Integrating Metagenomics and Activity-Based Metabolomics. Metabolites, 13(4), 530. https://doi.org/10.3390/metabo13040530