

Assessing the Biological Mechanisms Linking Smoking Behavior and Cognitive Function: A Mediation Analysis of Untargeted Metabolomics

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Data and Study Population
2.2. Smoking Status
2.3. Cognitive Function
2.4. Covariates
2.5. Metabolomic Data Collection
2.6. Metabolomic Data Quality Control
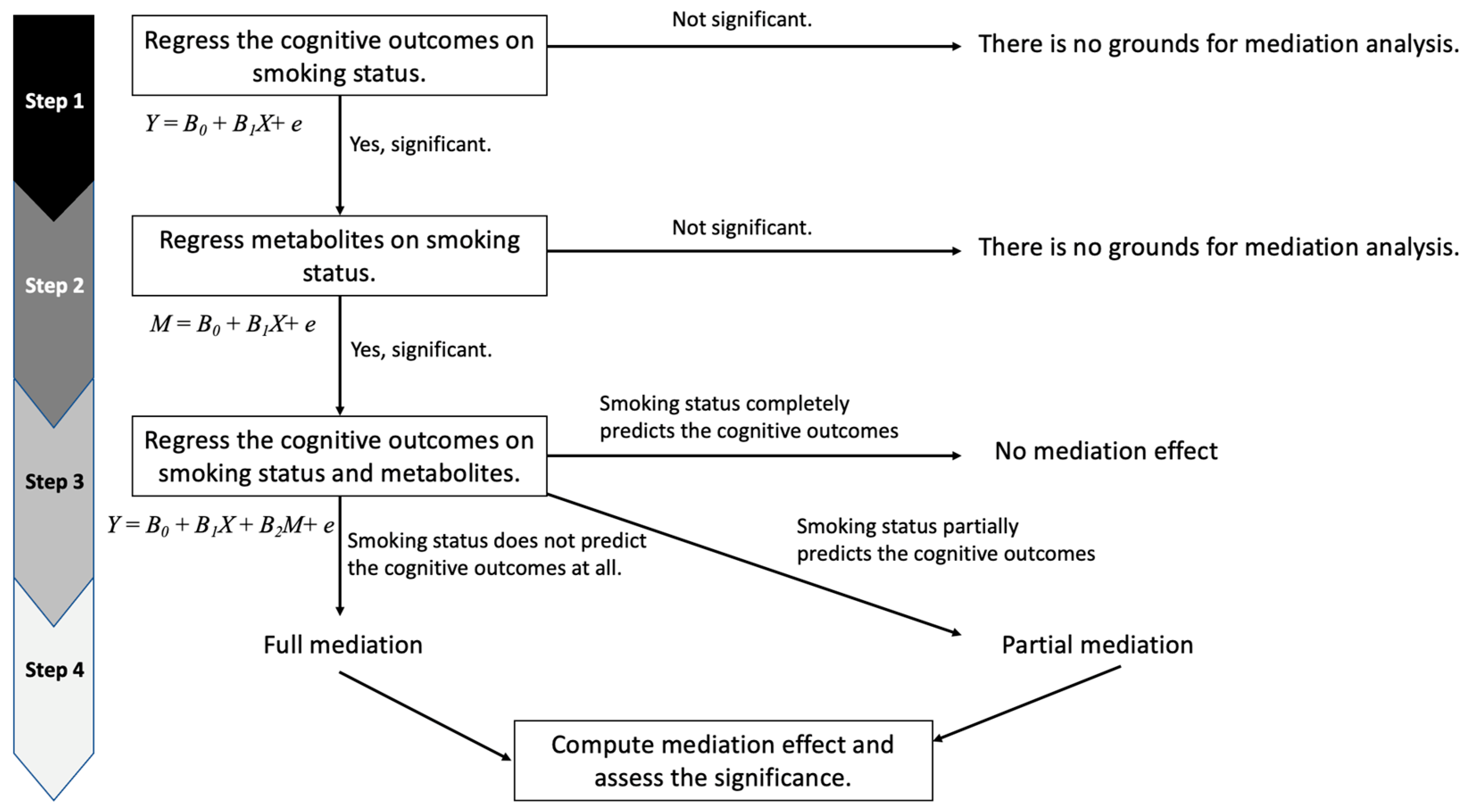
2.7. Statistical Mediation Analyses
3. Results
3.1. Sample Characteristics
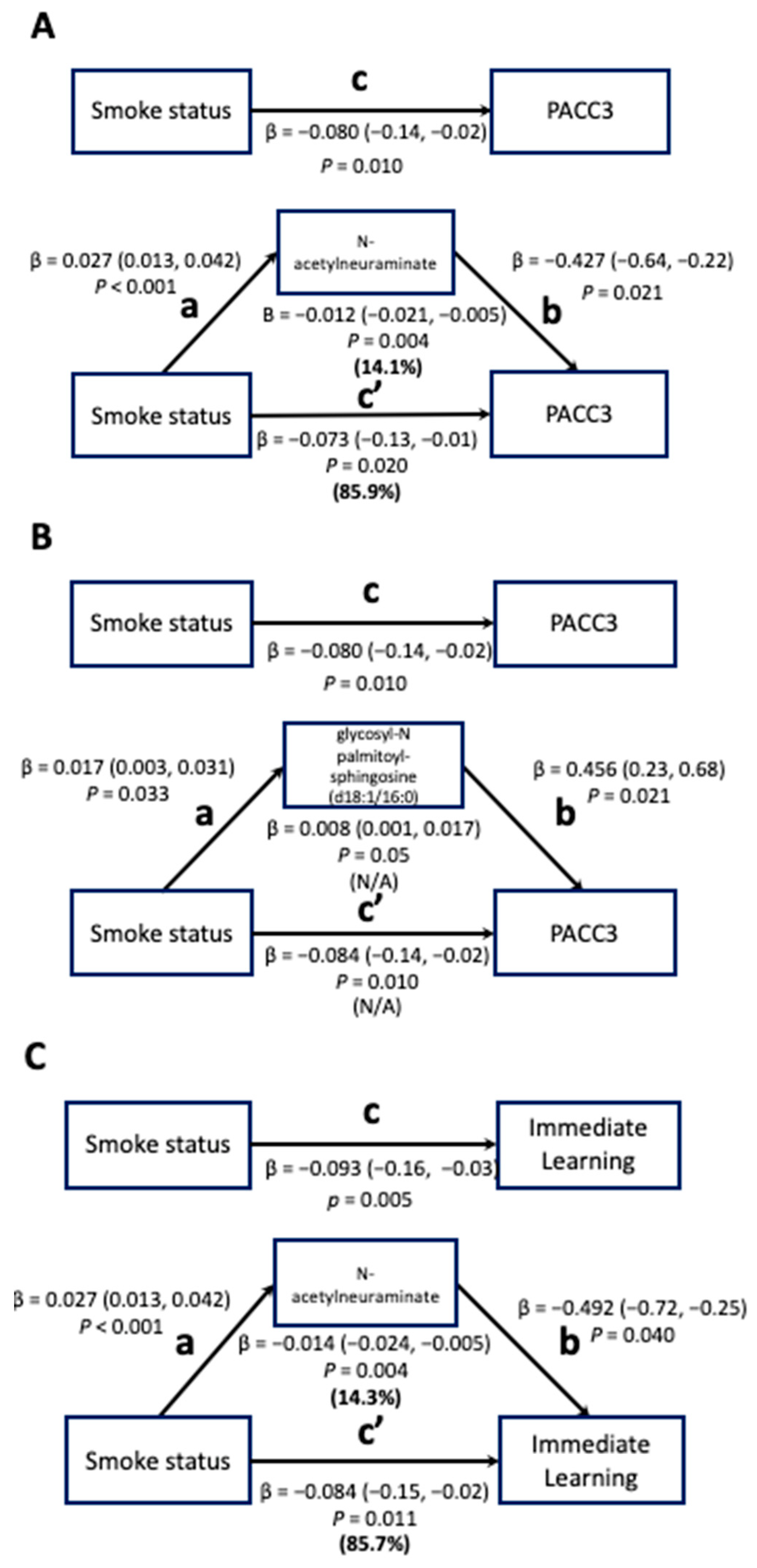
3.2. Mediation Analysis
3.2.1. Step 1: Associations between Smoking Status and Cognitive Function
3.2.2. Step 2: Associations between Smoking Status and Metabolomics
3.2.3. Step 3: Associations between Metabolites and Cognitive Function
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Centers for Disease Control and Prevention. Smoking & Tobacco Use. Fast Facts. 2021. Available online: https://www.cdc.gov/tobacco/data_statistics/fact_sheets/fast_facts/index.htm#beginning (accessed on 14 October 2023).
- Committee on the Public Health Implications of Raising the Minimum Age for Purchasing Tobacco Products. Board on Population Health and Public Health Practice; Institute of Medicine. In Public Health Implications of Raising the Minimum Age of Legal Access to Tobacco Products; Bonnie, R.J., Stratton, K., Kwan, L.Y., Eds.; National Academies Press (US): Washington, DC, USA, 2015. Available online: https://www.ncbi.nlm.nih.gov/books/NBK310413/ (accessed on 14 October 2023).
- McKenzie, J.; Bhatti, L.; Tursan d’Espaignet, E. WHO Tobacco Knowledge Summaries: Tobacco and dementia. Available online: https://apps.who.int/iris/bitstream/handle/10665/128041/WHO_NMH_PND_CIC_TKS_14.1_eng.pdf (accessed on 14 October 2023).
- Amidfar, M.; Askari, G.; Kim, Y.-K. Association of metabolic dysfunction with cognitive decline and Alzheimer’s disease: A review of metabolomic evidence. Prog. Neuropsychopharmacol. Biol. Psychiatry 2023, 128, 110848. [Google Scholar] [CrossRef]
- Dong, R.; Denier-Fields, D.N.; Van Hulle, C.A.; Kollmorgen, G.; Suridjan, I.; Wild, N.; Lu, Q.; Anderson, R.M.; Zetterberg, H.; Blennow, K.; et al. Identification of plasma metabolites associated with modifiable risk factors and endophenotypes reflecting Alzheimer’s disease pathology. Eur. J. Epidemiol. 2023, 38, 559–571. [Google Scholar] [CrossRef] [PubMed]
- Schrimpe-Rutledge, A.C.; Codreanu, S.G.; Sherrod, S.D.; McLean, J.A. Untargeted Metabolomics Strategies—Challenges and Emerging Directions. J. Am. Soc. Mass Spectrom. 2016, 27, 1897–1905. [Google Scholar] [CrossRef] [PubMed]
- Robinette, S.L.; Holmes, E.; Nicholson, J.K.; Dumas, M.E. Genetic determinants of metabolism in health and disease: From biochemical genetics to genome-wide associations. Genome Med. 2012, 4, 30. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, S. Emerging Insights into the Metabolic Alterations in Aging Using Metabolomics. Metabolites 2019, 9, 301. [Google Scholar] [CrossRef] [PubMed]
- Green, R.; Lord, J.; Xu, J.; Maddock, J.; Kim, M.; Dobson, R.; Legido-Quigley, C.; Wong, A.; Richards, M.; Proitsi, P. Metabolic Correlates of Late Midlife Cognitive Function: Findings from the 1946 British Birth Cohort. Brain Commun. 2022, 4, fcab291. [Google Scholar] [CrossRef] [PubMed]
- Proitsi, P.; Kuh, D.; Wong, A.; Maddock, J.; Bendayan, R.; Wulaningsih, W.; Hardy, R.; Richards, M. Lifetime cognition and late midlife blood metabolites: Findings from a British birth cohort. Transl. Psychiatry 2018, 8, 203. [Google Scholar] [CrossRef] [PubMed]
- Toledo, J.B.; Arnold, M.; Kastenmüller, G.; Chang, R.; Baillie, R.A.; Han, X.; Thambisetty, M.; Tenenbaum, J.D.; Suhre, K.; Thompson, J.W.; et al. Metabolic network failures in Alzheimer’s disease: A biochemical road map. Alzheimers Dement. J. Alzheimers Assoc. 2017, 13, 965–984. [Google Scholar] [CrossRef]
- Jiang, Y.; Zhu, Z.; Shi, J.; An, Y.; Zhang, K.; Wang, Y.; Li, S.; Jin, L.; Ye, W.; Cui, M.; et al. Metabolomics in the Development and Progression of Dementia: A Systematic Review. Front. Neurosci. 2019, 13, 343. [Google Scholar] [CrossRef]
- van der Lee, S.J.; Teunissen, C.E.; Pool, R.; Shipley, M.J.; Teumer, A.; Chouraki, V.; Melo van Lent, D.; Tynkkynen, J.; Fischer, K.; Hernesniemi, J.; et al. Circulating metabolites and general cognitive ability and dementia: Evidence from 11 cohort studies. Alzheimers Dement. 2018, 14, 707–722. [Google Scholar] [CrossRef] [PubMed]
- Shi, M.; Bazzano, L.A.; He, J.; Gu, X.; Li, C.; Li, S.; Yaffe, K.; Kinchen, J.M.; Stuchlik, P.; Mi, X.; et al. Novel serum metabolites associate with cognition phenotypes among Bogalusa Heart Study participants. Aging 2019, 11, 5124–5139. [Google Scholar] [CrossRef]
- Gu, F.; Derkach, A.; Freedman, N.D.; Landi, M.T.; Albanes, D.; Weinstein, S.J.; Mondul, A.M.; Matthews, C.E.; Guertin, K.A.; Xiao, Q.; et al. Cigarette smoking behaviour and blood metabolomics. Int. J. Epidemiol. 2016, 45, 1421–1432. [Google Scholar] [CrossRef] [PubMed]
- Sager, M.A.; Hermann, B.; La Rue, A. Middle-aged children of persons with Alzheimer’s disease: APOE genotypes and cognitive function in the Wisconsin Registry for Alzheimer’s Prevention. J. Geriatr. Psychiatry Neurol. 2005, 18, 245–249. [Google Scholar] [CrossRef] [PubMed]
- Johnson, S.C.; Koscik, R.L.; Jonaitis, E.M.; Clark, L.R.; Mueller, K.D.; Berman, S.E.; Bendlin, B.B.; Engelman, C.D.; Okonkwo, O.C.; Hogan, K.J.; et al. The Wisconsin Registry for Alzheimer’s Prevention: A review of findings and current directions. Alzheimers Dement. Amst. Neth. 2018, 10, 130–142. [Google Scholar] [CrossRef]
- Donohue, M.C.; Sperling, R.A.; Salmon, D.P.; Rentz, D.M.; Raman, R.; Thomas, R.G.; Weiner, M.; Aisen, P.S.; Australian Imaging, Biomarkers, and Lifestyle Flagship Study of Ageing; Alzheimer’s Disease Neuroimaging Initiative; et al. The preclinical Alzheimer cognitive composite: Measuring amyloid-related decline. JAMA Neurol. 2014, 71, 961–970. [Google Scholar] [CrossRef]
- Clark, L.R.; Racine, A.M.; Koscik, R.L.; Okonkwo, O.C.; Engelman, C.D.; Carlsson, C.M.; Asthana, S.; Bendlin, B.B.; Chappell, R.; Nicholas, C.R.; et al. Beta-amyloid and cognitive decline in late middle age: Findings from the Wisconsin Registry for Alzheimer’s Prevention study. Alzheimers Dement. J. Alzheimers Assoc. 2016, 12, 805–814. [Google Scholar] [CrossRef] [PubMed]
- Jonaitis, E.M.; Koscik, R.L.; Clark, L.R.; Ma, Y.; Betthauser, T.J.; Berman, S.E.; Allison, S.L.; Mueller, K.D.; Hermann, B.P.; Van Hulle, C.A.; et al. Measuring longitudinal cognition: Individual tests versus composites. Alzheimers Dement. Amst. Neth. 2019, 11, 74–84. [Google Scholar] [CrossRef]
- Bartels, C.; Wagner, M.; Wolfsgruber, S.; Ehrenreich, H.; Schneider, A. Alzheimer’s Disease Neuroimaging Initiative Impact of SSRI Therapy on Risk of Conversion From Mild Cognitive Impairment to Alzheimer’s Dementia in Individuals With Previous Depression. Am. J. Psychiatry 2018, 175, 232–241. [Google Scholar] [CrossRef] [PubMed]
- Tyas, S.L. Alcohol use and the risk of developing Alzheimer’s disease. Alcohol Res. Health 2001, 25, 299–306. [Google Scholar]
- Nordestgaard, L.T.; Tybjærg-Hansen, A.; Nordestgaard, B.G.; Frikke-Schmidt, R. Body Mass Index and Risk of Alzheimer’s Disease: A Mendelian Randomization Study of 399,536 Individuals. J. Clin. Endocrinol. Metab. 2017, 102, 2310–2320. [Google Scholar] [CrossRef]
- Darst, B.F.; Lu, Q.; Johnson, S.C.; Engelman, C.D. Integrated analysis of genomics, longitudinal metabolomics, and Alzheimer’s risk factors among 1111 cohort participants. Genet. Epidemiol. 2019, 43, 657–674. [Google Scholar] [CrossRef] [PubMed]
- Evans, A.M.; Bridgewater, B.R.; Liu, Q.; Mitchell, M.W.; Robinson, R.J.; Dai, H.; Stewart, S.J.; DeHaven, C.D.; Miller, L.A.D. High resolution mass spectrometry improves data quantity and quality as compared to unit mass resolution mass spectrometry in high-throughput profiling metabolomics. Metabolomics 2014, 4, 1. [Google Scholar] [CrossRef]
- Ford, L.; Kennedy, A.D.; Goodman, K.D.; Pappan, K.L.; Evans, A.M.; Miller LA, D.; Wulff, J.E.; Wiggs, B.R.; Lennon, J.J.; Elsea, S.; et al. Precision of a Clinical Metabolomics Profiling Platform for Use in the Identification of Inborn Errors of Metabolism. J. Appl. Lab. Med. 2020, 5, 342–356. [Google Scholar] [CrossRef] [PubMed]
- Baron, R.M.; Kenny, D.A. The moderator-mediator variable distinction in social psychological research: Conceptual, strategic, and statistical considerations. J. Pers. Soc. Psychol. 1986, 51, 1173–1182. [Google Scholar] [CrossRef] [PubMed]
- Judd, C.M.; Kenny, D.A. Process Analysis: Estimating Mediation in Treatment Evaluations. Eval. Rev. 1981, 5, 602–619. [Google Scholar] [CrossRef]
- James, L.R.; Brett, J.M. Mediators, moderators, and tests for mediation. J. Appl. Psychol. 1984, 69, 307–321. [Google Scholar] [CrossRef]
- Perini, G.; Cotta Ramusino, M.; Sinforiani, E.; Bernini, S.; Petrachi, R.; Costa, A. Cognitive impairment in depression: Recent advances and novel treatments. Neuropsychiatr. Dis. Treat. 2019, 15, 1249–1258. [Google Scholar] [CrossRef]
- Caspani, G.; Turecki, G.; Lam, R.W.; Milev, R.V.; Frey, B.N.; MacQueen, G.M.; Müller, D.J.; Rotzinger, S.; Kennedy, S.H.; Foster, J.A.; et al. Metabolomic signatures associated with depression and predictors of antidepressant response in humans: A CAN-BIND-1 report. Commun. Biol. 2021, 4, 903. [Google Scholar] [CrossRef]
- Michaud, T.L.; Siahpush, M.; Farazi, P.A.; Kim, J.; Yu, F.; Su, D.; Murman, D.L. The Association Between Body Mass Index, and Cognitive, Functional, and Behavioral Declines for Incident Dementia. J. Alzheimers Dis. 2018, 66, 1507–1517. [Google Scholar] [CrossRef]
- Ottosson, F.; Brunkwall, L.; Ericson, U.; Nilsson, P.M.; Almgren, P.; Fernandez, C.; Melander, O.; Orho-Melander, M. Connection Between BMI-Related Plasma Metabolite Profile and Gut Microbiota. J. Clin. Endocrinol. Metab. 2018, 103, 1491–1501. [Google Scholar] [CrossRef]
- Brennan, S.E.; McDonald, S.; Page, M.J.; Reid, J.; Ward, S.; Forbes, A.B.; McKenzie, J.E. Long-term effects of alcohol consumption on cognitive function: A systematic review and dose-response analysis of evidence published between 2007 and 2018. Syst. Rev. 2020, 9, 33. [Google Scholar] [CrossRef] [PubMed]
- Cederbaum, A.I. Alcohol metabolism. Clin. Liver Dis. 2012, 16, 667–685. [Google Scholar] [CrossRef]
- Tofighi, D.; MacKinnon, D.P. Monte Carlo Confidence Intervals for Complex Functions of Indirect Effects. Struct. Equ. Model. Multidiscip. J. 2016, 23, 194–205. [Google Scholar] [CrossRef]
- MacKinnon, D.P.; Fairchild, A.J.; Fritz, M.S. Mediation analysis. Annu. Rev. Psychol. 2007, 58, 593–614. [Google Scholar] [CrossRef] [PubMed]
- Varki, A. Sialic acids in human health and disease. Trends Mol. Med. 2008, 14, 351–360. [Google Scholar] [CrossRef]
- Rawal, P.; Zhao, L. Sialometabolism in Brain Health and Alzheimer’s Disease. Front. Neurosci. 2021, 15, 648617. [Google Scholar] [CrossRef]
- Miles, L.A.; Hermans, S.J.; Crespi, G.A.N.; Gooi, J.H.; Doughty, L.; Nero, T.L.; Markulić, J.; Ebneth, A.; Wroblowski, B.; Oehlrich, D.; et al. Small Molecule Binding to Alzheimer Risk Factor CD33 Promotes Aβ Phagocytosis. iScience 2019, 19, 110–118. [Google Scholar] [CrossRef]
- Zhao, L. CD33 in Alzheimer’s Disease—Biology, Pathogenesis, and Therapeutics: A Mini-Review. Gerontology 2019, 65, 323–331. [Google Scholar] [CrossRef]
- Mehdi, M.M.; Singh, P.; Rizvi, S.I. Erythrocyte sialic acid content during aging in humans: Correlation with markers of oxidative stress. Dis. Markers 2012, 32, 179–186. [Google Scholar] [CrossRef]
- Rajappa, M.; Ikkruthi, S.; Nandeesha, H.; Satheesh, S.; Sundar, I.; Ananthanarayanan, P.H.; Harichandrakumar, K.T. Relationship of raised serum total and protein bound sialic acid levels with hyperinsulinemia and indices of insulin sensitivity and insulin resistance in non-diabetic normotensive obese subjects. Diabetes Metab. Syndr. Clin. Res. Rev. 2013, 7, 17–19. [Google Scholar] [CrossRef]
- Prajna, K.; Kumar, A.; Rai, S.; Shetty, S.K.; Rai, T.; Begum, M.; Shashikala, M.D. Predictive Value of Serum Sialic Acid in Type-2 Diabetes Mellitus and Its Complication (Nephropathy). J. Clin. Diagn. Res. 2013, 7, 2435–2437. [Google Scholar] [CrossRef]
- Sillanaukee, P.; Pönniö, M.; Jääskeläinen, I.P. Occurrence of sialic acids in healthy humans and different disorders: Sialic acid in humans. Eur. J. Clin. Investig. 1999, 29, 413–425. [Google Scholar] [CrossRef] [PubMed]
- Suzzi, S.; Croese, T.; Ravid, A.; Gold, O.; Clark, A.R.; Medina, S.; Kitsberg, D.; Adam, M.; Vernon, K.A.; Kohnert, E.; et al. N-acetylneuraminic acid links immune exhaustion and accelerated memory deficit in diet-induced obese Alzheimer’s disease mouse model. Nat. Commun. 2023, 14, 1293. [Google Scholar] [CrossRef] [PubMed]
- Showing Metabocard for GlcCer(d18:1/16:0) (HMDB0004971). The Metabolomics Innovation Centre. Available online: https://hmdb.ca/metabolites/HMDB0004971#references (accessed on 14 October 2023).
- Wang, Q.; Ji, X.; Rahman, I. Dysregulated Metabolites Serve as Novel Biomarkers for Metabolic Diseases Caused by E-Cigarette Vaping and Cigarette Smoking. Metabolites 2021, 11, 345. [Google Scholar] [CrossRef]
- Mill, J.; Patel, V.; Okonkwo, O.; Li, L.; Raife, T. Erythrocyte sphingolipid species as biomarkers of Alzheimer’s disease. J. Pharm. Anal. 2022, 12, 178–185. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
| Variable | Value |
|---|---|
| Age (years) | |
| Mean (SD) | 58.5 (6.47) |
| Median (Min, Max) | 59.0 (40.7, 75.0) |
| Female sex | 891 (70.4%) |
| White race | 1200 (94.8%) |
| College or graduate degree | 777 (61.4%) |
| Smoking status | |
| Never smoker | 711 (56.2%) |
| Former smoker | 469 (37.0%) |
| Current smoker | 86 (6.8%) |
| CES-D score (60 points) | |
| Mean (SD) | 7.13 (7.16) |
| Median (Min, Max) | 5.00 (0, 44.0) |
| Body Mass Index | |
| Underweight | 8 (0.6%) |
| Normal | 352 (27.8%) |
| Overweight | 449 (35.5%) |
| Obese | 457 (36.1%) |
| Weekly alcohol consumption | |
| Mean (SD) | 4.25 (6.74) |
| Median (Min, Max) | 2.00 (0, 52.5) |
| PACC3 | |
| Mean (SD) | 0.0160 (0.760) |
| Median (Min, Max) | 0.0717 (−3.17, 2.55) |
| Immediately Learning (IMM) | |
| Mean (SD) | 0.0145 (0.783) |
| Median (Min, Max) | 0.0365 (−3.41, 2.36) |
| Delayed Recall (DEL) | |
| Mean (SD) | 0.0155 (0.780) |
| Median (Min, Max) | 0.109 (−3.94, 1.96) |
| Executive Function (EXE) | |
| Mean (SD) | 0.00546 (0.822) |
| Median (Min, Max) | 0.0970 (−6.42, 2.43) |
| Predictors | PACC3 | Immediate Learning | Delayed Recall | Executive Function | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Beta | SE | 95% CI | p | Beta | SE | 95% CI | p | Beta | SE | 95% CI | p | Beta | SE | 95% CI | p | |
| Intercept | −1.098 | 0.095 | −1.285–−0.910 | <0.001 | −0.955 | 0.101 | −1.154–−0.756 | <0.001 | −0.881 | 0.102 | −1.082–−0.681 | <0.001 | −1.059 | 0.104 | −1.262–−0.855 | <0.001 |
| Female | 0.529 | 0.039 | 0.452–0.605 | <0.001 | 0.443 | 0.041 | 0.362–0.523 | <0.001 | 0.388 | 0.041 | 0.307–0.469 | <0.001 | 0.354 | 0.042 | 0.271–0.437 | <0.001 |
| White race | 0.400 | 0.083 | 0.236–0.563 | <0.001 | 0.353 | 0.088 | 0.181–0.525 | <0.001 | 0.327 | 0.089 | 0.153–0.501 | <0.001 | 0.552 | 0.092 | 0.372–0.732 | <0.001 |
| College or graduate degree | 0.375 | 0.037 | 0.302–0.448 | <0.001 | 0.358 | 0.039 | 0.281–0.434 | <0.001 | 0.358 | 0.040 | 0.280–0.435 | <0.001 | 0.266 | 0.041 | 0.186–0.346 | <0.001 |
| CES-D | −0.005 | 0.001 | −0.007–−0.002 | <0.001 | −0.005 | 0.001 | −0.008–−0.002 | 0.001 | −0.004 | 0.001 | −0.007–−0.001 | 0.005 | −0.007 | 0.001 | −0.010–−0.005 | <0.001 |
| BMI (underweight) | −0.238 | 0.093 | −0.421–−0.055 | 0.011 | −0.157 | 0.109 | −0.372–0.057 | 0.151 | −0.090 | 0.109 | −0.303–0.124 | 0.41 | −0.170 | 0.090 | −0.346–0.007 | 0.06 |
| BMI (overweight) | 0.004 | 0.024 | −0.042–0.051 | 0.855 | 0.011 | 0.028 | −0.043–0.065 | 0.691 | −0.001 | 0.027 | −0.054–0.053 | 0.984 | 0.004 | 0.023 | −0.041–0.050 | 0.858 |
| BMI (obese) | 0.032 | 0.029 | −0.025–0.089 | 0.27 | 0.039 | 0.033 | −0.026–0.104 | 0.244 | 0.026 | 0.033 | −0.039–0.090 | 0.441 | −0.027 | 0.029 | −0.083–0.030 | 0.358 |
| Alcohol Weekly Consumption | 0.005 | 0.002 | 0.002–0.008 | 0.002 | 0.005 | 0.002 | 0.002–0.009 | 0.004 | 0.004 | 0.002 | 0.000–0.007 | 0.034 | 0.006 | 0.001 | 0.003–0.009 | <0.001 |
| Former smoker | 0.053 | 0.038 | −0.021–0.127 | 0.159 | 0.025 | 0.039 | −0.052–0.102 | 0.524 | 0.057 | 0.040 | −0.021–0.135 | 0.151 | 0.024 | 0.041 | −0.056–0.105 | 0.557 |
| Current smoker | −0.383 | 0.073 | −0.526–−0.240 | <0.001 | −0.360 | 0.077 | −0.510–−0.210 | <0.001 | −0.360 | 0.077 | −0.512–−0.209 | <0.001 | −0.270 | 0.078 | −0.424–−0.117 | 0.001 |
| Age | −0.047 | 0.003 | −0.052–−0.041 | <0.001 | −0.039 | 0.003 | −0.045–−0.033 | <0.001 | −0.035 | 0.003 | −0.041–−0.029 | <0.001 | −0.060 | 0.003 | −0.066–−0.054 | <0.001 |
| Age2 | −0.001 | 0.000 | −0.001–−0.001 | <0.001 | −0.001 | 0.000 | −0.001–−0.000 | <0.001 | −0.001 | 0.000 | −0.001–−0.000 | <0.001 | −0.001 | 0.000 | −0.001–−0.001 | <0.001 |
| Practice Effect | 0.082 | 0.008 | 0.067–0.098 | <0.001 | 0.124 | 0.009 | 0.107–0.141 | <0.001 | 0.117 | 0.009 | 0.100–0.134 | <0.001 | 0.074 | 0.008 | 0.058–0.091 | <0.001 |
| Random Effects | ||||||||||||||||
| ICC | 0.78 | 0.71 | 0.73 | 0.83 | ||||||||||||
| N (family) | 1007 | 1007 | 1007 | 1007 | ||||||||||||
| N (Individual) | 1266 | 1266 | 1266 | 1266 | ||||||||||||
| Observations | 4680 | 4680 | 4680 | 4680 | ||||||||||||
| Marginal R2/Conditional R2 | 0.298/0.844 | 0.216/0.773 | 0.194/0.779 | 0.283/0.881 | ||||||||||||
| Cognitive Function | Plasma Metabolite | Estimates | 95% CI | p | FDR |
|---|---|---|---|---|---|
| PACC3 | glycosyl-N palmitoyl-sphingosine (d18:1/16:0) | 0.456 | 0.680–0.233 | <0.001 | 0.021 |
| N-acetylneuraminate | −0.427 | −0.636–−0.219 | <0.001 | 0.021 | |
| IMM | N-acetylneuraminate | −0.429 | −0.733–−0.251 | <0.001 | 0.040 |
| EXE | metabolonic lactone sulfate | 0.177 | 0.091–0.263 | <0.001 | 0.035 |
| androstenediol (3alpha, 17alpha) monosulfate (2) | 0.135 | 0.066–0.204 | <0.001 | 0.037 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Choi, J.J.; Koscik, R.L.; Jonaitis, E.M.; Panyard, D.J.; Morrow, A.R.; Johnson, S.C.; Engelman, C.D.; Schmitz, L.L. Assessing the Biological Mechanisms Linking Smoking Behavior and Cognitive Function: A Mediation Analysis of Untargeted Metabolomics. Metabolites 2023, 13, 1154. https://doi.org/10.3390/metabo13111154
Choi JJ, Koscik RL, Jonaitis EM, Panyard DJ, Morrow AR, Johnson SC, Engelman CD, Schmitz LL. Assessing the Biological Mechanisms Linking Smoking Behavior and Cognitive Function: A Mediation Analysis of Untargeted Metabolomics. Metabolites. 2023; 13(11):1154. https://doi.org/10.3390/metabo13111154
Chicago/Turabian StyleChoi, Jerome J., Rebecca L. Koscik, Erin M. Jonaitis, Daniel J. Panyard, Autumn R. Morrow, Sterling C. Johnson, Corinne D. Engelman, and Lauren L. Schmitz. 2023. "Assessing the Biological Mechanisms Linking Smoking Behavior and Cognitive Function: A Mediation Analysis of Untargeted Metabolomics" Metabolites 13, no. 11: 1154. https://doi.org/10.3390/metabo13111154
APA StyleChoi, J. J., Koscik, R. L., Jonaitis, E. M., Panyard, D. J., Morrow, A. R., Johnson, S. C., Engelman, C. D., & Schmitz, L. L. (2023). Assessing the Biological Mechanisms Linking Smoking Behavior and Cognitive Function: A Mediation Analysis of Untargeted Metabolomics. Metabolites, 13(11), 1154. https://doi.org/10.3390/metabo13111154