
Rubi idaei fructus as a Source of Bioactive Chemical Compounds with an Important Role in Human Health and Comparison of the Antioxidant Potential of Fruits and Juice of Three Repeat-Fruiting Rubus idaeus L. Cultivars

Abstract
:1. Introduction
1.1. Pro-Health Antioxidant Activity of Rubus Fruits
1.1.1. Antidiabetic Properties
1.1.2. Anti-Obesity Properties
1.1.3. Anticancer Properties
1.2. Rationale behind the Study
2. Materials and Methods
2.1. Study Material
Origin and Lineage of the Analyzed Cultivars
2.2. Determination of Antioxidant Activity
2.2.1. FRAP Method (Ferric Reducing Antioxidant Power Assay)
2.2.2. DPPH Method
2.3. Chemical Analyses
2.3.1. Available Carbohydrates
2.3.2. Total Fiber
2.3.3. Total Protein
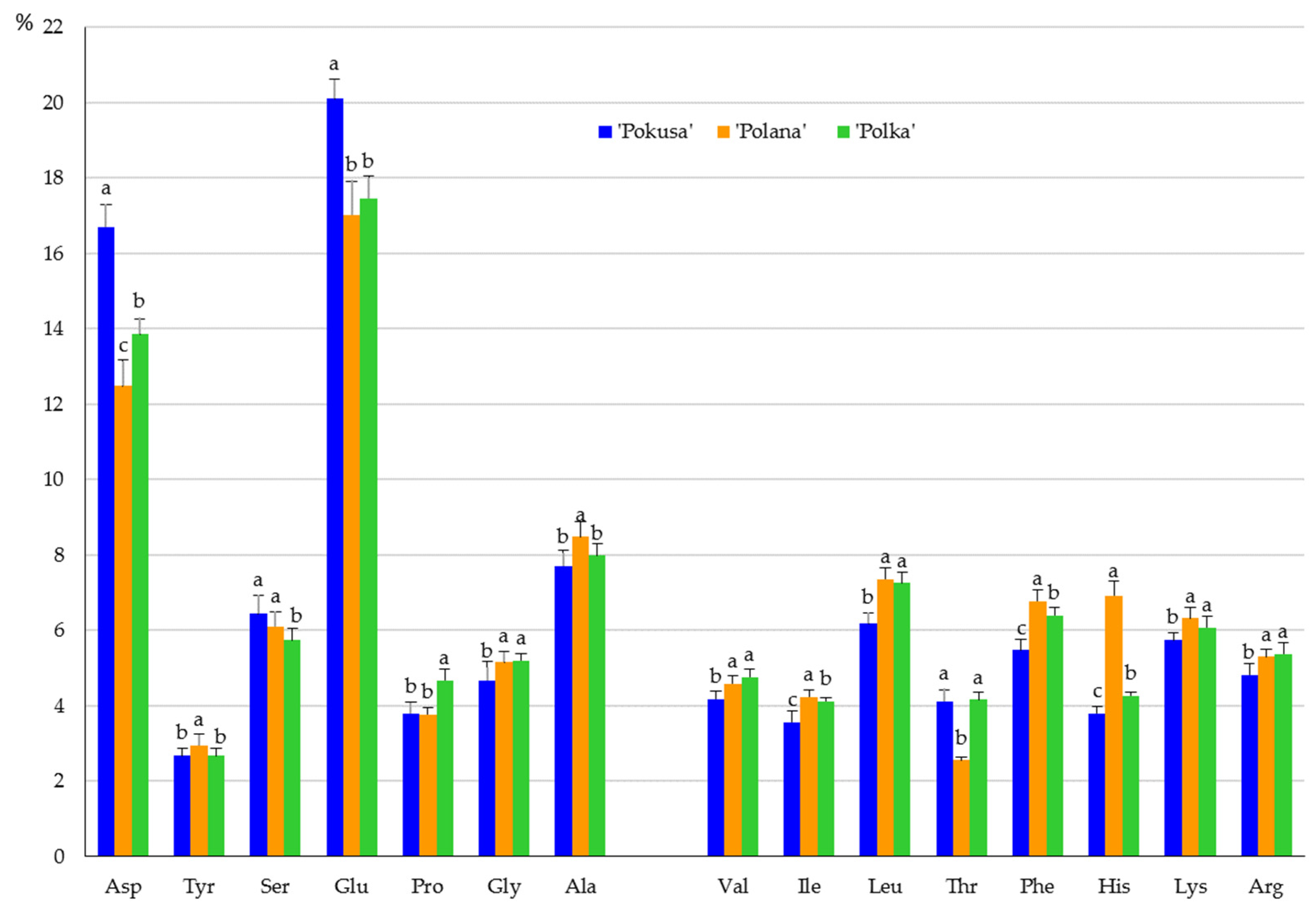
2.3.4. Amino Acids
2.3.5. Total Fat Content
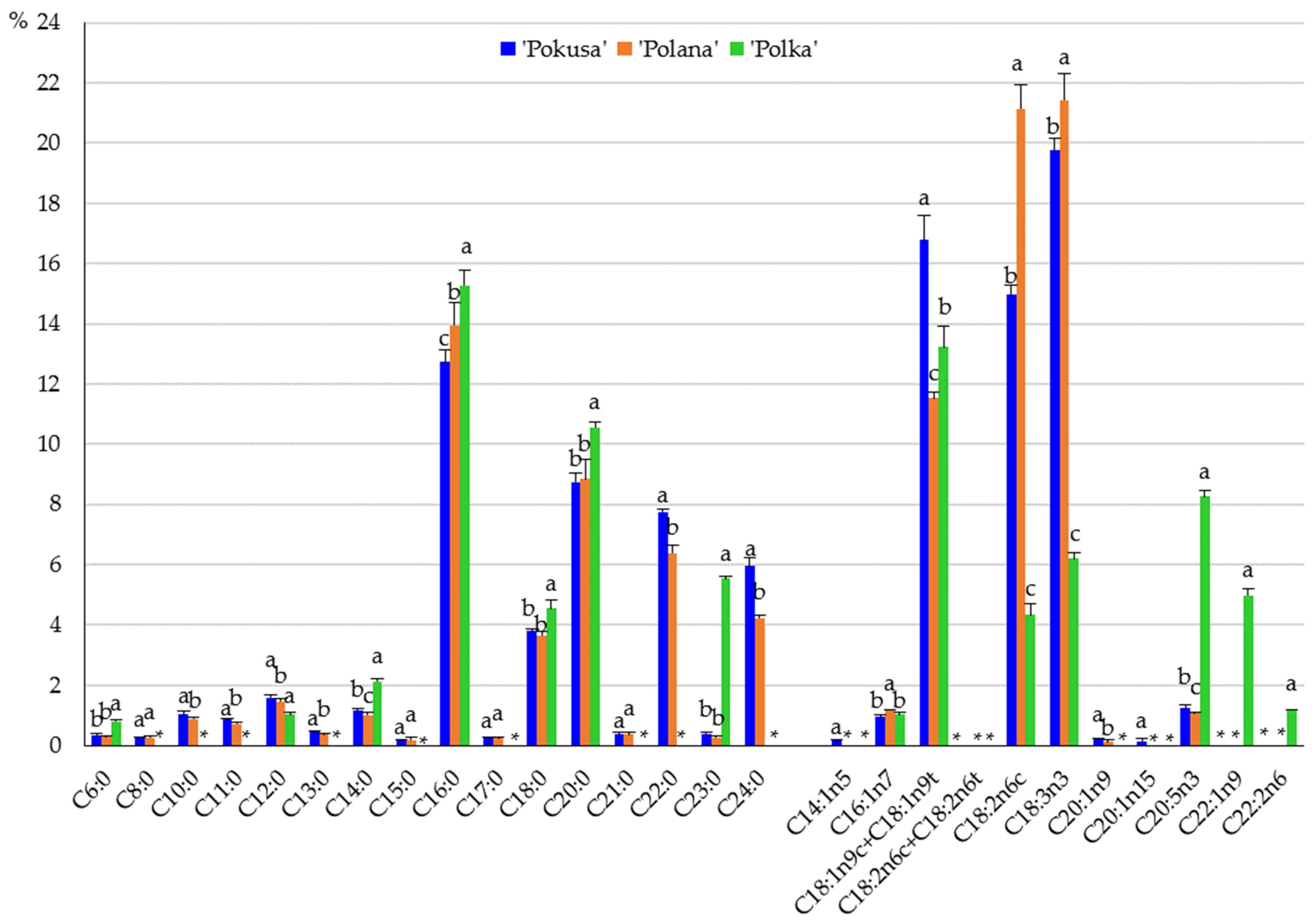
2.3.6. Qualitative and Quantitative Composition of Fatty Acids
2.3.7. Folin–Ciocalteu Method
2.3.8. Anthocyanins
2.3.9. Vitamin C
2.3.10. Flavonoids
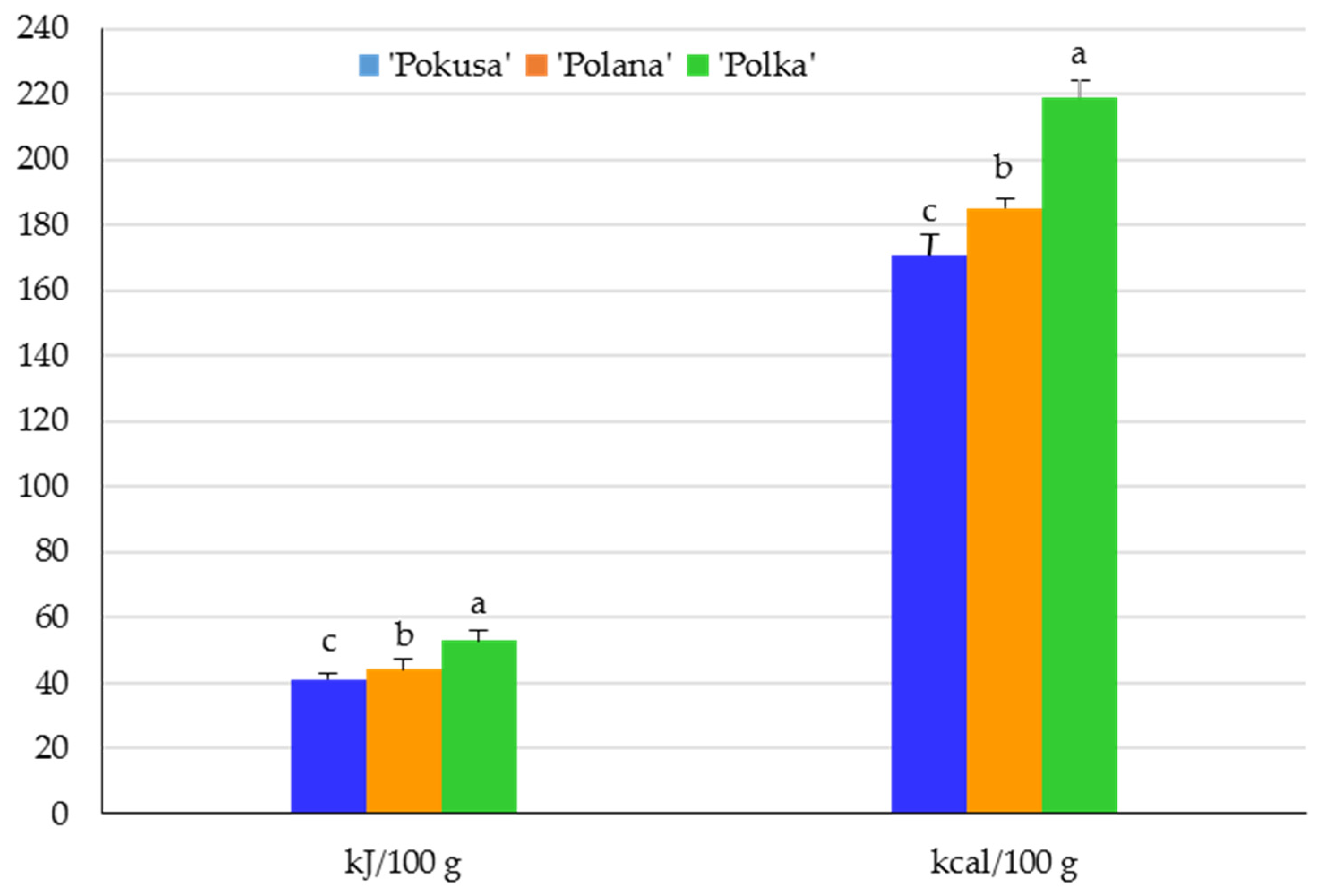
2.3.11. Energy Value
2.4. Statistical Analysis
3. Results
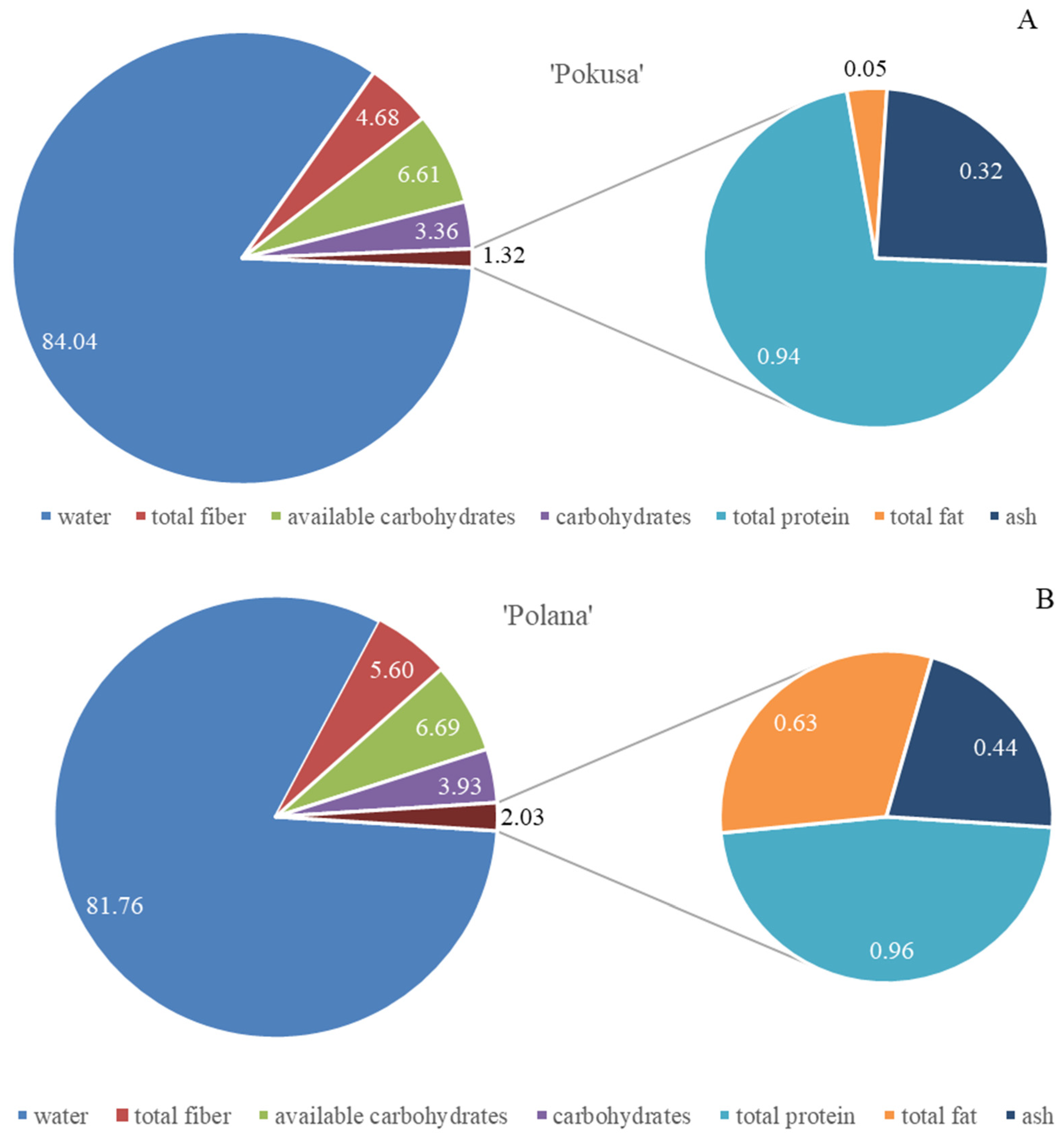
3.1. Selected Nutrients and Energy Value
3.2. Antioxidants: Vitamin C, Anthocyanins, and Flavonoids
3.3. Protein Amino Acids
3.4. Fatty Acids
3.5. Antioxidant Activity
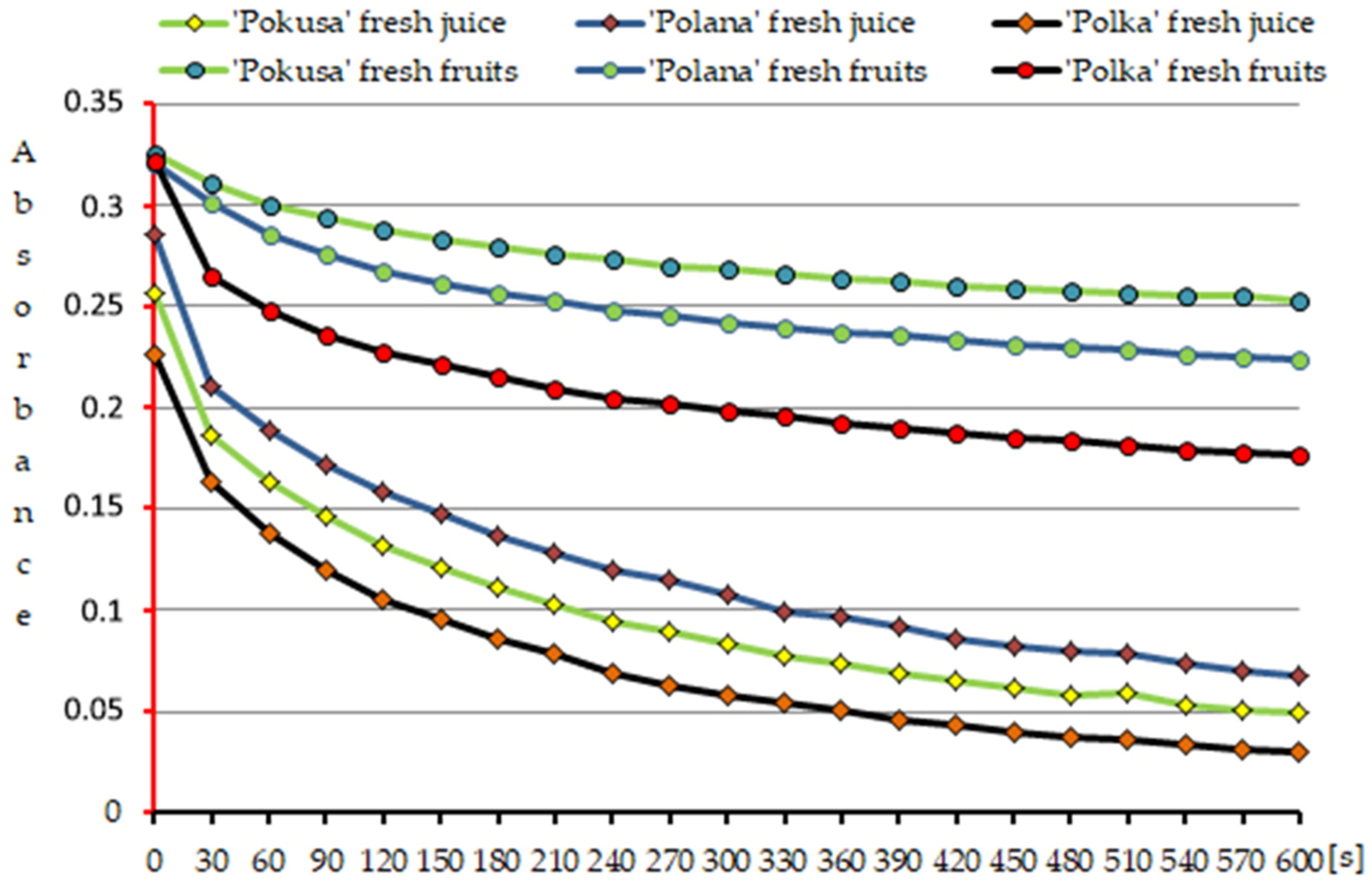
3.5.1. Fruits
3.5.2. Fresh Juice
4. Discussion
4.1. Phenolic Compounds
4.2. Fiber
4.3. Anthocyanins
4.4. Flavonoids
4.5. Vitamin C
4.6. Amino Acids
4.7. Fatty Acids
4.8. Antioxidant Activity
4.9. Applications
4.10. Future Research
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Stagos, D. Antioxidant activity of polyphenolic plant extracts. Antioxidants 2020, 9, 19. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Corona, A.V.; Valencia-Espinosa, I.; González-Sánchez, F.A.; Sánchez-López, A.L.; Garcia-Amezquita, L.E.; Garcia-Varela, R. Antioxidant, anti-inflammatory and cytotoxic activity of phenolic compound family extracted from raspberries (Rubus idaeus): A general review. Antioxidants 2022, 11, 1192. [Google Scholar] [CrossRef]
- Mannino, G.; Serio, G.; Gaglio, R.; Busetta, G.; La Rosa, L.; Lauria, A.; Settanni, L.; Gentile, C. Phytochemical profile and antioxidant, antiproliferative, and antimicrobial properties of Rubus idaeus seed powder. Foods 2022, 11, 2605. [Google Scholar] [CrossRef]
- Petruskevicius, A.; Viskelis, J.; Urbonaviciene, D.; Viskelis, P. Anthocyanin accumulation in berry fruits and their antimicrobial and antiviral properties: An overview. Horticulturae 2023, 9, 288. [Google Scholar] [CrossRef]
- Marić, B.; Abramović, B.; Ilić, N.; Bodroža-Solarov, M.; Pavlić, B.; Oczkowski, M.; Wilczak, J.; Četojević-Simin, D.; Šarić, L.; Teslić, N. UHPLC-Triple-TOF-MS Characterization, antioxidant, antimicrobial and antiproliferative activity of raspberry (Rubus idaeus L.) seed extracts. Foods 2023, 12, 161. [Google Scholar] [CrossRef]
- Gil-Martínez, L.; Mut-Salud, N.; Ruiz-García, J.A.; Falcón-Piñeiro, A.; Maijó-Ferré, M.; Baños, A.; De la Torre-Ramírez, J.M.; Guillamón, E.; Verardo, V.; Gómez-Caravaca, A.M. Phytochemicals determination and antioxidant, antimicrobial, anti-inflammatory and anticancer activities of blackberry fruits. Foods 2023, 12, 1505. [Google Scholar] [CrossRef]
- Sławińska, N.; Prochoń, K.; Olas, B. A review on berry seeds—A special Emphasis on their chemical content and health-promoting properties. Nutrients 2023, 15, 1422. [Google Scholar] [CrossRef]
- Kim, K.J.; Jeong, E.S.; Lee, K.H.; Na, J.R.; Park, S.; Kim, J.S.; Na, C.S.; Kim, Y.R.; Kim, S. Unripe Rubus coreanus Miquel extract containing ellagic acid promotes lipolysis and thermogenesis in vitro and in vivo. Molecules 2020, 25, 5954. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q.; Naeem, A.; Zou, J.; Yu, C.; Wang, Y.; Chen, J.; Ping, Y. Isolation of phenolic compounds from raspberry based on molecular imprinting techniques and investigation of their anti-Alzheimer’s disease properties. Molecules 2022, 27, 6893. [Google Scholar] [CrossRef] [PubMed]
- Gu, I.; Brownmiller, C.; Howard, L.; Lee, S.O. Chemical composition of volatile extracts from black raspberries, blueberries, and blackberries and their antiproliferative effect on A549 non-small-cell lung cancer cells. Life 2022, 12, 2056. [Google Scholar] [CrossRef] [PubMed]
- Salim, G.M.; El-Dakdouki, M.H.; Abdallah, H.; Nasser, H.M.; Arnold-Apostolides, N. Antioxidative and hepatoprotective effects of Rubus canescens DC. growing wild in Lebanon. J. Nat. Prod. 2021, 11, 44–56. [Google Scholar] [CrossRef]
- Rambaran, T.F.; Nembhard, N.; Bowen-Forbes, C.S.; Alexander-Lindo, R.L. Hypoglycemic effect of the fruit extracts of two varieties of Rubus rosifolius. J. Food Biochem. 2020, 44, 13365. [Google Scholar] [CrossRef]
- Loizzo, M.R.; Tundis, R.; Leporini, M.; D’Urso, G.; Candela, R.G.; Falco, T.; Piacente, S.; Bruno, M.; Sottile, F. Almond (Prunus dulcis cv. Casteltermini) skin confectionery by-products: New opportunity for the development of a functional blackberry (Rubus ulmifolius Schott) Jam. Antioxidants 2021, 10, 1218. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Zhang, X.; Yu, J.; Du, J.; Wu, X.; Chen, L.; Wang, R.; Wu, Y.; Li, Y. Constituents of the fruits of Rubus chingii Hu and their neuroprotective effects on human neuroblastoma SH-SY5Y cells. Food Res. Int. 2023, 173, 113255. [Google Scholar] [CrossRef] [PubMed]
- Land Lail, H.; Feresin, R.G.; Hicks, D.; Stone, B.; Price, E.; Wanders, D. Berries as a treatment for obesity-induced inflammation: Evidence from preclinical models. Nutrients 2021, 13, 334. [Google Scholar] [CrossRef] [PubMed]
- Papaioanou, M.; Chronopoulou, E.G.; Ciobotari, G.; Efrose, R.C.; Sfichi-Duke, L.; Chatzikonstantinou, M.; Pappa, E.; Ganopoulos, I.; Madesis, P.; Nianiou-Obeidat, I. Cosmeceutical properties of two cultivars of red raspberry grown under different conditions. Cosmetics 2018, 5, 20. [Google Scholar] [CrossRef]
- Huo, Y.; Zhao, X.; Zhao, J.; Kong, X.; Li, L.; Yuan, T.; Xu, J. Hypoglycemic effects of Fu-Pen-Zi (Rubus chingii Hu) fruit extracts in streptozotocin-induced type 1 diabetic mice. J. Funct. Foods 2021, 87, 104837. [Google Scholar] [CrossRef]
- De Ancos, M.; González, B.E.; Cano, M.P. Ellagic acid, vitamin C, and total phenolic contents and radical scavenging capacity affected by freezing and frozen storage in raspberry fruit. J. Agric. Food Chem. 2000, 48, 4565–4570. [Google Scholar] [CrossRef] [PubMed]
- Bobinaitė, R.; Viškelis, P.; Venskutonis, P.R. Variation of total phenolics, anthocyanins, ellagic acid and radical scavenging capacity in various raspberry (Rubus spp.) cultivars. Food Chem. 2012, 132, 1495–1501. [Google Scholar] [CrossRef] [PubMed]
- He, B.; Dai, L.; Jin, L.; Liu, Y.; Li, X.; Luo, M.; Wang, Z.; Kai, G. Bioactive components, pharmacological effects, and drug development of traditional herbal medicine Rubus chingii Hu (Fu-Pen-Zi). Front. Nutr. 2023, 9, 1052504. [Google Scholar] [CrossRef] [PubMed]
- An, J.H.; Kim, D.L.; Lee, T.B.; Kim, K.J.; Kim, S.H.; Kim, N.H.; Kim, H.Y.; Choi, D.S.; Kim, S.G. Effect of Rubus occidentalis extract on metabolic parameters in subjects with prediabetes: A proof-of-concept, randomized, double-blind, placebo-controlled clinical trial. Phytother. Res. 2016, 30, 1634–1640. [Google Scholar] [CrossRef] [PubMed]
- Ion, R.M.; Sibianu, M.; Hutanu, A.; Beresescu, F.G.; Sala, D.T.; Flavius, M.; Rosca, A.; Constantin, C.; Scurtu, A.; Moriczi, R.; et al. A comprehensive summary of the current understanding of the relationship between severe obesity, metabolic syndrome, and inflammatory status. J. Clin. Med. 2023, 12, 3818. [Google Scholar] [CrossRef] [PubMed]
- Stepan, M.D.; Vintilescu, Ș.B.; Streață, I.; Podeanu, M.A.; Florescu, D.N. The role of vitamin D in obese children with non-alcoholic fatty liver disease and associated metabolic syndrome. Nutrients 2023, 15, 2113. [Google Scholar] [CrossRef]
- Wang, L.; Wei, Y.; Ning, C.; Zhang, M.; Fan, P.; Lei, D.; Du, J.; Gale, M.; Ma, Y.; Yang, Y. Ellagic acid promotes browning of white adipose tissues in high-fat diet-induced obesity in rats through suppressing white adipocyte maintaining genes. Endocr. J. 2019, 66, 923–936. [Google Scholar] [CrossRef] [PubMed]
- Jeong, M.Y.; Kim, H.L.; Park, J.; An, H.J.; Kim, S.H.; Kim, S.J.; So, H.S.; Park, R.; Um, J.Y.; Hong, S.H. Rubi fructus (Rubus coreanus) inhibits differentiation to adipocytesc in 3T3-L1 cells. Evid. Based Complement. Altern. Med. 2013, 2013, 475386. [Google Scholar] [CrossRef]
- Tu, L.; Sun, H.; Tang, M.; Zhao, J.; Zhang, Z.; Sun, X.; He, S. Red raspberry extract (Rubus idaeus L shrub) intake ameliorates hyperlipidemia in HFD-induced mice through PPAR signaling pathway. Food Chem. Toxicol. 2019, 133, 110796. [Google Scholar] [CrossRef]
- Ke, H.; Bao, T.; Chen, W. Polysaccharide from Rubus chingii Hu affords protection against palmitic acid-induced lipotoxicity in human hepatocytes. Int. J. Biol. Macromol. 2019, 133, 1063–1071. [Google Scholar] [CrossRef]
- Harper, G.M. Geriatrics experts explore relationship between heart disease and cancer, the top two leading causes of death. J. Gerontol. Nurs. 2022, 48, 55–56. [Google Scholar] [CrossRef]
- Siegel, R.L.; Miller, K.D.; Wagle, N.S.; Jemal, A. Cancer statistics. CA Cancer J. Clin. 2023, 73, 17–48. [Google Scholar] [CrossRef] [PubMed]
- Ding, Y.; Ye, B.; Sun, Z.; Mao, Z.; Wang, W. Reactive oxygen species-mediated pyroptosis with the help of nanotechnology: Prospects for cancer therapy. Adv. NanoBiomed Res. 2023, 3, 2200077. [Google Scholar] [CrossRef]
- Li, Z.; Zhao, T.; Ding, J.; Gu, H.; Wang, Q.; Wang, Y.; Zhang, D.; Gao, C. Reactive oxygen species-responsive hydrogel encapsulated with bone marrow derived stem cells promotes repair and regeneration of spinal cord injury. Bioact. Mater. 2023, 19, 550–568. [Google Scholar] [CrossRef] [PubMed]
- Nowak, A.; Sójka, M.; Klewicka, E.; Lipinska, L.; Klewicki, R.; Kołodziejczyk, K. Ellagitannins from Rubus idaeus L. exert geno- and cytotoxic effects against human colon adenocarcinoma cell line Caco-2. J. Agric. Food Chem. 2017, 65, 2947–2955. [Google Scholar] [CrossRef]
- Assad, N.K.; Dheeba, B.I.; Mohammad, F.I.; Hamad, A.I. Anti-cancer activity of the Rubus idaeus extracts against HepG2 and L20B cell lines using tissue culture technique. Egypt. Acad. J. Biol. Sci. 2015, 7, 19–23. [Google Scholar] [CrossRef]
- Chen, T.; Shi, N.; Afzali, A. Chemopreventive effects of strawberry and black raspberry on colorectal cancer in inflammatory bowel disease. Nutrients 2019, 11, 1261. [Google Scholar] [CrossRef]
- Danek, J. ‘Polka’ and ‘Pokusa’—New primocane fruiting raspberry cultivars from Poland. Acta Hortic. 2002, 585, 197–198. [Google Scholar] [CrossRef]
- Benzie, I.F.F.; Strain, J.J. The ferric reducing ability of plasma (FRAP) as a measure of “antioxidant power”: The FRAP assay. Anal. Biochem. 1996, 239, 70–76. [Google Scholar] [CrossRef]
- Bondet, V.; Brand-Williams, W.; Ber, C. Kinetics and mechanism of antioxidant activity using the DPPH• free radical method. LWT Food Sci. Technol. 1997, 30, 609–615. [Google Scholar] [CrossRef]
- Dekker, K.D. The Luff-Schoorl method for determination of reducing sugar in juices, molasses and sugar. S. Afr. Sugar J. 1950, 34, 157–171. [Google Scholar]
- Asp, N.G.; Johansson, C.G.; Hallmer, H.; Siljeströem, M. Rapid enzymic assay of insoluble and soluble dietary fiber. J. Agric. Food Chem. 1983, 31, 476–482. [Google Scholar] [CrossRef]
- Sáez-Plaza, P.; Michałowski, T.; Navas, M.J.; Asuero, A.G.; Wybraniec, S. An overview of the Kjeldahl method of nitrogen determination. Part I. Early history, chemistry of the procedure, and titrimetric finish. Crit. Rev. Anal. Chem. 2013, 43, 178–223. [Google Scholar] [CrossRef]
- PN75/A-04018:1975/Az3:2002; Agricultural Food Products: Nitrogen Contents Determination with Kjeldahl’s Method and Recalculation into Protein. Polish Committee for Standardization: Warsaw, Poland, 2002. (In Polish)
- Rabie, A.L.; Wells, J.D.; Dent, L.K. The nitrogen content of pollen protein. J. Apic. Res. 1983, 22, 119–123. [Google Scholar] [CrossRef]
- Davies, M.G.; Thomas, A.J. An investigation of hydrolytic techniques for the amino acid analysis of foodstuffs. J. Sci. Food Agric. 1973, 24, 1525–1540. [Google Scholar] [CrossRef]
- PN-EN ISO 5508:1996; Animal and Vegetable Fats and Oils: Analysis by Gas Chromatography of Methyl esters of Fatty Acids. Polish Committee for Standardization: Warsaw, Poland, 1996. (In Polish)
- PN-EN ISO 12966-1:2014; Animal and Vegetable Fats and Oils: Gas Chromatography of Fatty Acids Methyl Esters: Part 1: Guidelines on Modern Gas Chromatography of Fatty Acid Methyl Esters. ISO: Geneva, Switzerland, 2014.
- Prior, R.L.; Wu, X.; Schaich, K. Standardized methods for the determination of antioxidant capacity and phenolics in foods and dietary supplements. J. Agric. Food Chem. 2005, 53, 4290–4302. [Google Scholar] [CrossRef] [PubMed]
- Martínez, A.E.; Favret, E.A. Anthocyanin synthesis and lengthening in the first leaf of barley isogenic lines. Plant Sci. 1990, 71, 35–43. [Google Scholar] [CrossRef]
- PN-EN 14130:2004; Foodstuffs—Determination of Vitamin C by HPLC. Polish Committee for Standardization: Warsaw, Poland, 2004.
- Lamaison, J.L.C.; Carnet, A. Teneurs en principaux flavonoids des fleurs de Crataegeus monogyna Jacq et de Crataegeus laevigata (Poiret D.C) en function de la vegetation. Pharm. Acta Helv. 1990, 65, 315–320. [Google Scholar]
- European Commission. UE Commission Delegated Regulation No 78/2014. Regulation of the European Parliament and the European Council (EU) no. 1169/2011 from 25 October 2011 with amendments. Regulation of the European Council (EU) no. 1155/2013 from 21 August 2013, Regulation of the European Council (EU) no. 78/2014 from 22 November 2013 (applied from 19 February 2014); European Commission: Brussels, Belgium, 2014. [Google Scholar]
- Cervantes, L.; Martínez-Ferri, E.; Soria, C.; Ariza, M.T. Bioavailability of phenolic compounds in strawberry, raspberry and blueberry: Insights for breeding programs. Food Biosci. 2020, 37, 100680. [Google Scholar] [CrossRef]
- Dobani, S.; Latimer, C.; McDougall, G.J.; Allwood, J.W.; Pereira-Caro, G.; Moreno-Rojas, J.M.; Ternan, N.G.; Pourshahidi, L.K.; Lawther, R.; Tuohy, K.M.; et al. Ex vivo fecal fermentation of human ileal fluid collected after raspberry consumption modifies (poly) phenolics and modulates genoprotective effects in colonic epithelial cells. Redox Biol. 2021, 40, 101862. [Google Scholar] [CrossRef] [PubMed]
- Viuda-Martos, M.; Lucas-Gonzalez, R.; Ballester-Costa, C.; Pérez-Álvarez, J.A.; Muñoz, L.A.; Fernández-López, J. Evaluation of protective effect of different dietary fibers on polyphenolic profile stability of maqui berry (Aristotelia chilensis (Molina) Stuntz) during in vitro gastrointestinal digestion. Food Funct. 2018, 9, 573–584. [Google Scholar] [CrossRef] [PubMed]
- Baenas, N.; Nuñez-Gómez, V.; Navarro-González, I.; Sánchez-Martínez, L.; García-Alonso, J.; Periago, M.J.; González-Barrio, R. Raspberry dietary dibre: Chemical properties, functional evaluation and prebiotic in vitro effect. LWT Food Sci. Technol. 2020, 134, 110140. [Google Scholar] [CrossRef]
- Noratto, G.D.; Chew, B.P.; Antieza, L.M. Red raspberry (Rubus idaeus L.) intake decreases oxidative stress in obese diabetic (db/db) mice. Food Chem. 2017, 227, 305–314. [Google Scholar] [CrossRef]
- Li, M.; Liu, Y.; Yang, G.; Sun, L.; Song, X.; Chen, Q.; Bao, Y.; Luo, T.; Wang, J. Microstructure, physicochemical properties, and adsorption capacity of deoiled red raspberry pomace and its total dietary fiber. LWT Food Sci. Technol. 2022, 153, 112478. [Google Scholar] [CrossRef]
- Tomas, M. Effect of dietary fiber addition on the content and in vitro bioaccessibility of antioxidants in red raspberry puree. Food Chem. 2022, 375, 131897. [Google Scholar] [CrossRef]
- Ivanović, M.; Pavlović, A.; Mitić, M.; Pecev-Marinković, E.; Krstić, J.; Mrmošanin, J. Determination of total and individual anthocyanins in raspberries grown in South Serbia. In Proceedings of the XXI 21: Savetovanje o Biotehnologiji sa Međunarodnim Učešćem, Čačak, Serbia, 11–12 March 2016; Volume 21, pp. 263–267. [Google Scholar]
- Sánchez-Velaázquez, O.A.; Cortes-Rodriguez, M.; Milán-Carrillo, J.; Montes-Ávila, J.; Robles-Bañuelos, B.; del Ángel, A.S.; Cuevas-Rodríguez, E.O.; Rangel-López, E. Anti-oxidant and anti-proliferative effect of anthocyanin enriched fractions from two Mexican wild blackberries (Rubus spp.) on HepG2 and glioma cell lines. J. Berry Res. 2020, 10, 513–529. [Google Scholar] [CrossRef]
- Im, S.E.; Nam, T.G.; Lee, H.; Han, M.W.; Heo, H.J.; Koo, S.I.; Lee, C.Y.; Kim, D.O. Anthocyanins in the ripe fruits of Rubus coreanus Miquel and their protective effect on neuronal PC-12 cells. Food Chem. 2013, 139, 604–610. [Google Scholar] [CrossRef] [PubMed]
- Murapa, P.; Dai, J.; Chung, M.; Mumper, R.J.; D’Orazio, J. Anthocyanin-rich fractions of blackberry extracts reduce UV-induced free radicals and oxidative damage in keratinocytes. Phytother. Res. 2012, 26, 106–112. [Google Scholar] [CrossRef] [PubMed]
- Grabek-Lejko, D.; Miłek, M.; Sidor, E.; Puchalski, C.; Dżugan, M. Antiviral and antibacterial effect of honey enriched with Rubus spp. as a functional food with enhanced antioxidant properties. Molecules 2022, 27, 4859. [Google Scholar] [CrossRef]
- Wu, Y.; Zhang, C.; Huang, Z.; Lyu, L.; Li, J.; Li, W.; Wu, W. The color difference of Rubus fruits is closely related to the composition of flavonoids including anthocyanins. LWT Food Sci. Technol. 2021, 149, 111825. [Google Scholar] [CrossRef]
- Zejak, D.; Glisic, I.; Spalevic, V.; Maskovic, P.; Dudic, B. Sustainable management of fruit growing in rural areas of Montenegro: The impact of location on the phonological and nutritional properties on raspberry (Rubus idaeus L.). Agronomy 2021, 11, 1663. [Google Scholar] [CrossRef]
- Veljković, B.; Djordjević, N.; Dolićanin, Z.; Ličina, B.; Topuzović, M.; Stanković, M.; Zlatić, N.; Dajić-Stevanović, Z. Antioxidant and anticancer properties of leaf and fruit extracts of the wild raspberry (Rubus idaeus L.). Not. Bot. Horti Agrobot. Cluj-Napoca 2019, 47, 359–367. [Google Scholar] [CrossRef]
- Veljković, B.; Jakovljević, V.; Stanković, M.; Dajić-Stevanović, Z. Phytochemical and antioxidant properties of fresh fruits and some traditional products of wild grown raspberry (Rubus idaeus L.). Not. Bot. Horti Agrobot. Cluj-Napoca 2019, 47, 565–573. [Google Scholar] [CrossRef]
- George, B.P.; Parimelazhagan, T.; Chandran, R. Evaluation of total phenolic content, antioxidant and analgesic potential of Rubus fairholmianus gard. Int. J. Pharm. Pharm. Sci. 2013, 5 (Suppl. 3), 484–488. [Google Scholar]
- Choi, G.J.; Kang, H.; Lee, O.H.; Kwon, J.W. Effect of immature Rubus occidentalis on postoperative pain in a rat model. Medicina 2023, 59, 264. [Google Scholar] [CrossRef] [PubMed]
- Henrotin, Y.; Cozannet, R.L.; Fança-Berthon, P.; Truillet, R.; Cohen-Solhal, M.; DunnGalvin, G.; Grouin, J.M.; Doolan, A. Rubus idaeus extract improves symptoms in knee osteoarthritis patients: Results from a phase II double-blind randomized controlled trial. BMC Musculoskelet. Disord. 2022, 23, 650. [Google Scholar] [CrossRef] [PubMed]
- Kopustinskiene, D.M.; Jakstas, V.; Savickas, A.; Bernatoniene, J. Flavonoids as anticancer agents. Nutrients 2020, 12, 457. [Google Scholar] [CrossRef]
- Yang, Y.; Zhang, K.; Xiao, Y.; Zhang, L.; Huang, Y.; Li, X.; Chen, S.; Peng, Y.; Yang, S.; Liu, Y.; et al. Genome assembly and population resequencing reveal the geographical divergence of shanmei (Rubus corchorifolius). Genom. Proteom. Bioinform. 2022, 20, 1106–1118. [Google Scholar] [CrossRef]
- Chen, Q.; Liu, X.; Hu, Y.; Sun, B.; Hu, Y.; Wang, X.; Tang, H.; Wang, Y. Transcriptomic profiling of fruit development in black raspberry Rubus coreanus. Int. J. Genom. 2018, 2018, 8084032. [Google Scholar] [CrossRef]
- Tosun, M.; Ercisli, S.; Karlidag, H.; Sengul, M. Characterization of red raspberry (Rubus idaeus L.) genotype for their physiochemical properties. J. Food Sci. 2009, 74, 575–579. [Google Scholar] [CrossRef] [PubMed]
- Kazimierczak, R.; Hallmann, E.; Kowalska, K.; Rembiałkowska, E. Biocompounds content in organic and conventional raspberry fruits. Acta Fytotech. Zootech. 2016, 18, 40–42. [Google Scholar] [CrossRef]
- Ponder, A.; Hallmann, E. The nutritional value and vitamin C content of different raspberry cultivars from organic and conventional production. J. Food Compos. Anal. 2020, 87, 103429. [Google Scholar] [CrossRef]
- Pantelidis, G.E.; Vasilakakis, M.; Manganaris, G.A.; Diamantidis, G.R. Antioxidant capacity, phenol, anthocyanin and ascorbic acid contents in raspberries, blackberries, red currants, gooseberries and cornelian cherries. Food Chem. 2007, 102, 777–783. [Google Scholar] [CrossRef]
- Akimov, M.Y.; Koltsov, V.A.; Zhbanova, E.V.; Akimova, O.M. Nutritional value of promising raspberry varieties. IOP Conf. Ser. Earth Environ. Sci. 2021, 640, 022078. [Google Scholar] [CrossRef]
- Surya, M.I.; Suhartati, S.; Ismaini, L.; Lusini, Y.; Anggraeni, D.; Normasiwi, S.; Asni, N.; Bakar Sidiq, M.A. Fruit nutrients of five species of wild raspberries (Rubus spp.) from Indonesian mountain’s forests. J. Trop. Life Sci. 2018, 8, 75–80. [Google Scholar] [CrossRef]
- Shad, Z.; Arsalan, A.; Bano, R.; Khan, M.F.; Ahmed, I. Physicochemical, biochemical and antioxidant properties of ascorbic acid. J. Baqai Med. Univ. 2011, 14, 33–40. [Google Scholar]
- Pehlivan, F.E. Vitamin C: An antioxidant agent. In Vitamin C; Hamza, A.H., Ed.; IntechOpen: London, UK, 2017; Chapter 2; pp. 23–35. [Google Scholar]
- Hertog, M.G.L.; Hollman, P.C.H.; Katan, M.B. Content of potentially anticancerogenic flavonoids of 28 vegetables and 9 fruits commonly consumed in the Netherlands. J. Agric. Food Chem. 1992, 40, 2379–2383. [Google Scholar] [CrossRef]
- Zhang, M.; Jativa, D.F. Vitamin C supplementation in the critically ill: A systematic review and meta-analysis. SAGE Open Med. 2018, 6, 2050312118807615. [Google Scholar] [CrossRef] [PubMed]
- Kwon, J.; Nishisaka, M.M.; McGrath, A.F.; Kristo, A.S.; Sikalidis, A.K.; Reaves, S.K. Protein intake in NCAA Division 1 soccer players: Assessment of daily amounts, distribution patterns, and leucine levels as a quality indicator. Sports 2023, 11, 45. [Google Scholar] [CrossRef]
- Holowaty, M.N.; Lees, M.J.; Abou Sawan, S.; Paulussen, K.J.M.; Jäger, R.; Purpura, M.; Paluska, S.A.; Burd, N.A.; Hodson, N.; Moore, D.R. Leucine ingestion promotes mTOR translocation to the periphery and enhances total and peripheral RPS6 phosphorylation in human skeletal muscle. Amino Acids 2023, 55, 253–261. [Google Scholar] [CrossRef]
- Wang, B.; Du, M. Increasing adipocyte number and reducing adipocyte size: The role of retinoids in adipose tissue development and metabolism. Crit. Rev. Food Sci. Nutr. 2023, 2023, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Zorzi, M.; Gai, F.; Medana, C.; Aigotti, R.; Morello, S.; Peiretti, P.G. Bioactive compounds and antioxidant capacity of small berries. Foods 2020, 9, 623. [Google Scholar] [CrossRef]
- Celik, F.; Ercisli, S. Lipid and fatty acid composition of wild and cultivated red raspberry fruits (Rubus idaeus L.). J. Med. Plant Res. 2009, 3, 583–585. Available online: http://www.academicjournals.org/JMPR (accessed on 11 June 2023).
- Vara, A.L.; Pinela, J.; Dias, M.I.; Petrović, J.; Nogueira, A.; Soković, M.; Ferreira, I.C.F.R.; Barros, L. Compositional features of the “Kweli” red raspberry and its antioxidant and antimicrobial activities. Foods 2020, 9, 1522. [Google Scholar] [CrossRef]
- Kafkas, E.; Özgen, M.; Özoğul, Y.; Türemiş, N. Phytochemical and fatty scid profile of selected red raspberry cultivars: A comparative study. J. Food Qual. 2008, 31, 67–78. [Google Scholar] [CrossRef]
- Cekiç, C.; Özgen, M. Comparison of antioxidant capacity and phytochemical properties of wild and cultivated red raspberries (Rubus idaeus L.). J. Food Compos. Anal. 2010, 23, 540–544. [Google Scholar] [CrossRef]
- Gramza-Michałowska, A.; Bueschke, M.; Kulczyński, B.; Gliszczyńska-Świgło, A.; Kmiecik, D.; Bilska, A.; Purłan, M.; Wałęsa, L.; Ostrowski, M.; Filipczuk, M.; et al. Phenolic compounds and multivariate analysis of antiradical properties of red fruits. J. Food Meas. Charact. 2019, 13, 1739–1747. [Google Scholar] [CrossRef]
- Gülçin, I.; Topal, F.; Çakmakçı, R.; Bilsel, M.; Gören, A.C.; Erdogan, U. Pomological features, nutritional quality, polyphenol content analysis, and antioxidant properties of domesticated and 3 wild ecotype forms of raspberries (Rubus idaeus L.). J. Food Sci. 2011, 76, C585–C593. [Google Scholar] [CrossRef]
- Wu, J.; Zhang, D.; Zhu, B.; Wang, S.; Xu, Y.; Zhang, C.; Yang, H.; Wang, S.; Liu, P.; Qin, L.; et al. Rubus chingii Hu. unripe fruits extract ameliorates carbon tetrachloride-induced liver fibrosis and improves the associated gut microbiota imbalance. Chin. Med. 2022, 17, 56. [Google Scholar] [CrossRef] [PubMed]
- Martins, M.S.; Gonçalves, A.C.; Alves, G.; Silva, L.R. Blackberries and mulberries: Berries with significant health-promoting properties. Int. J. Mol. Sci. 2023, 24, 12024. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cultivar | Folin–Ciocalteu | |||
|---|---|---|---|---|
| Min.–Max. | Mean ± SD | Min.–Max. | Mean ± SD | |
| Fruits (mg/g f.w.) | Juice (mg/mL f.w.) | |||
| ‘Pokusa’ | 1.69–1.73 | 1.71 ± 0.02 a | 216.45–227.36 | 223.72 ± 6.30 c |
| ‘Polana’ | 1.59–1.75 | 1.68 ± 0.09 a | 232.81–260.70 | 250.98 ± 15.7 b |
| ‘Polka’ | 1.57–1.73 | 1.65 ± 0.08 a | 243.71–276.42 | 260.07 ± 16.3 a |
| Cultivar | FRAP μmol/g | Kinetics of DPPH• Radical Reduction | |||||
|---|---|---|---|---|---|---|---|
| TEC50 [s] | DPPH rem % | AE dm³/µmol × s | |||||
| Min.–Max. | Mean ± SD | Mean ± SD | Min.–Max. | Mean ± SD | Min.–Max. | Mean ± SD | |
| ‘Pokusa’ | 12.51–12.64 | 12.59 ± 0.07 b | 600 ± 0.0 | 86.17–87.23 | 86.61 ± 0.55 a | 0.00140–0.00143 | 0.00142 ± 0.00002 c |
| ‘Polana’ | 9.09–11.60 | 20.27 ± 1.26 a | 600 ± 0.0 | 82.71–86.97 | 85.20 ± 2.21 a | 0.00159–0.00163 | 0.00161 ± 0.00002 b |
| ‘Polka’ | 12.28–13.17 | 12.79 ± 0.46 b | 600 ± 0.0 | 78.72–92.29 | 83.24 ± 7.83 a | 0.00189–0.00197 | 0.00193 ± 0.00004 a |
| Cultivar | FRAP μmol/g | Kinetics of DPPH• Radical Reduction | ||||||
|---|---|---|---|---|---|---|---|---|
| TEC50 [s] | DPPH rem % | AE dm³/µmol × s | ||||||
| Min.–Max. | Mean ± SD | Mean ± SD | Min.–Max. | Mean ± SD | Min.–Max. | Min.–Max. | Mean ± SD | |
| ‘Pokusa’ | 10000.01–13780.49 | 12317.08 ± 2029.6 a | 245–298 | 276.67 ± 27.97 b | 77.66–78.99 | 78.28 ± 0.67 a | 0.00328–0.00385 | 0.00352 ± 0.0003 b |
| ‘Polana’ | 12012.20–12743.90 | 12276.42 ± 405.9 b | 234–352 | 291.67 ± 59.05 a | 72.61–77.93 | 76.06 ± 3.00 b | 0.00286–0.00422 | 0.00348 ± 0.0006 c |
| ‘Polka’ | 10000.01–12378.05 | 11707.32 ± 948.5 c | 120–182 | 141.67 ± 34.96 c | 59.84–60.37 | 60.20 ± 0.31 c | 0.00630–0.00907 | 0.00807 ± 0.0015 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chwil, M.; Matraszek-Gawron, R.; Kostryco, M. Rubi idaei fructus as a Source of Bioactive Chemical Compounds with an Important Role in Human Health and Comparison of the Antioxidant Potential of Fruits and Juice of Three Repeat-Fruiting Rubus idaeus L. Cultivars. Metabolites 2023, 13, 1124. https://doi.org/10.3390/metabo13111124
Chwil M, Matraszek-Gawron R, Kostryco M. Rubi idaei fructus as a Source of Bioactive Chemical Compounds with an Important Role in Human Health and Comparison of the Antioxidant Potential of Fruits and Juice of Three Repeat-Fruiting Rubus idaeus L. Cultivars. Metabolites. 2023; 13(11):1124. https://doi.org/10.3390/metabo13111124
Chicago/Turabian StyleChwil, Mirosława, Renata Matraszek-Gawron, and Mikołaj Kostryco. 2023. "Rubi idaei fructus as a Source of Bioactive Chemical Compounds with an Important Role in Human Health and Comparison of the Antioxidant Potential of Fruits and Juice of Three Repeat-Fruiting Rubus idaeus L. Cultivars" Metabolites 13, no. 11: 1124. https://doi.org/10.3390/metabo13111124
APA StyleChwil, M., Matraszek-Gawron, R., & Kostryco, M. (2023). Rubi idaei fructus as a Source of Bioactive Chemical Compounds with an Important Role in Human Health and Comparison of the Antioxidant Potential of Fruits and Juice of Three Repeat-Fruiting Rubus idaeus L. Cultivars. Metabolites, 13(11), 1124. https://doi.org/10.3390/metabo13111124