
Evaluating the Effects of Omega-3 Polyunsaturated Fatty Acids on Inflammatory Bowel Disease via Circulating Metabolites: A Mediation Mendelian Randomization Study


Abstract
:1. Introduction
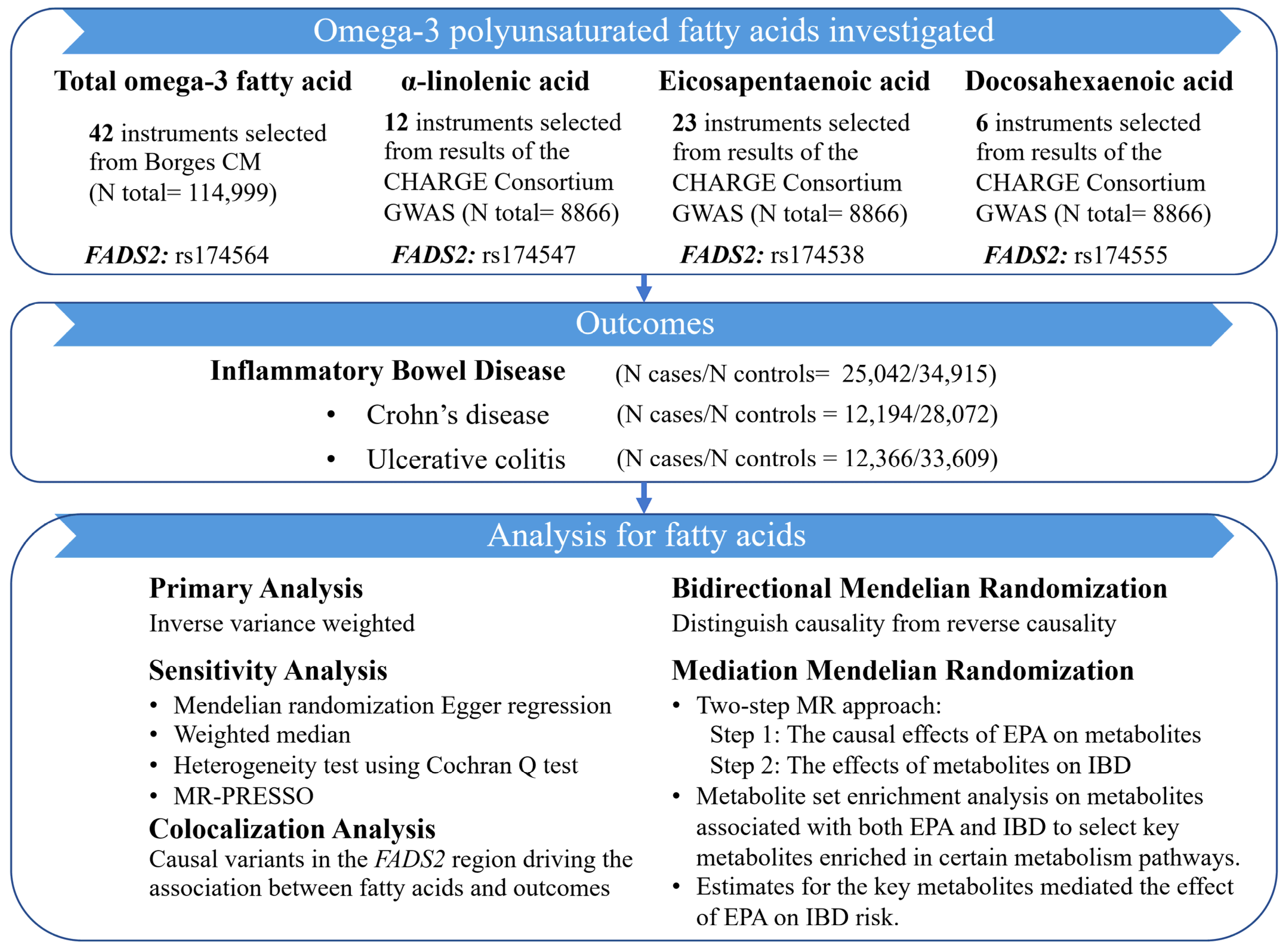
2. Materials and Methods
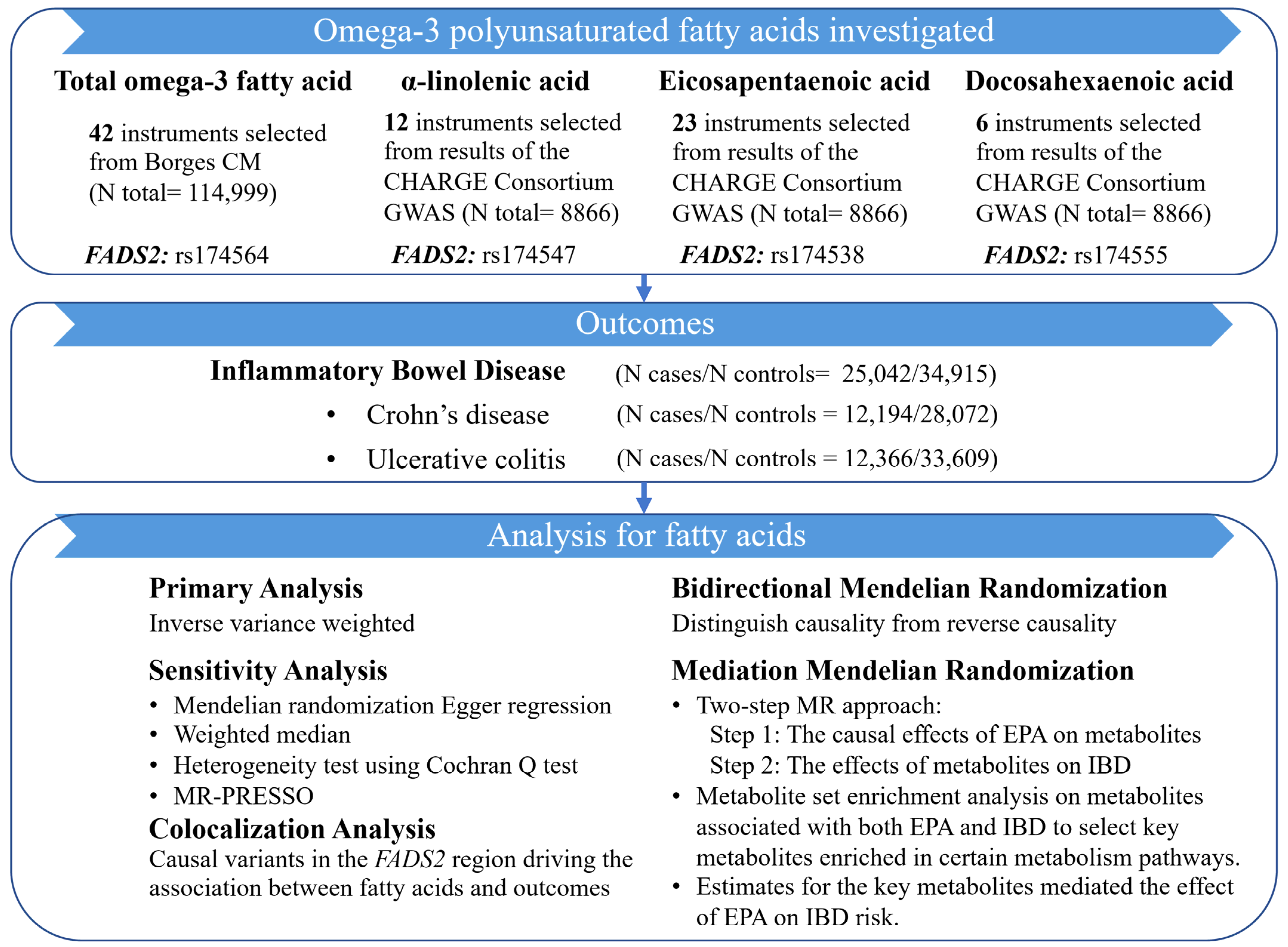
2.1. Study Design
2.2. Data Sources and Genetic Instruments for Omega-3 PUFAs
2.3. Outcome Data Sources
2.4. Metabolite Data Sources
2.5. Statistical Analysis
2.5.1. Two-Sample MR Analysis
2.5.2. Mediation MR Analysis Linking EPA with IBD via Metabolites
3. Results
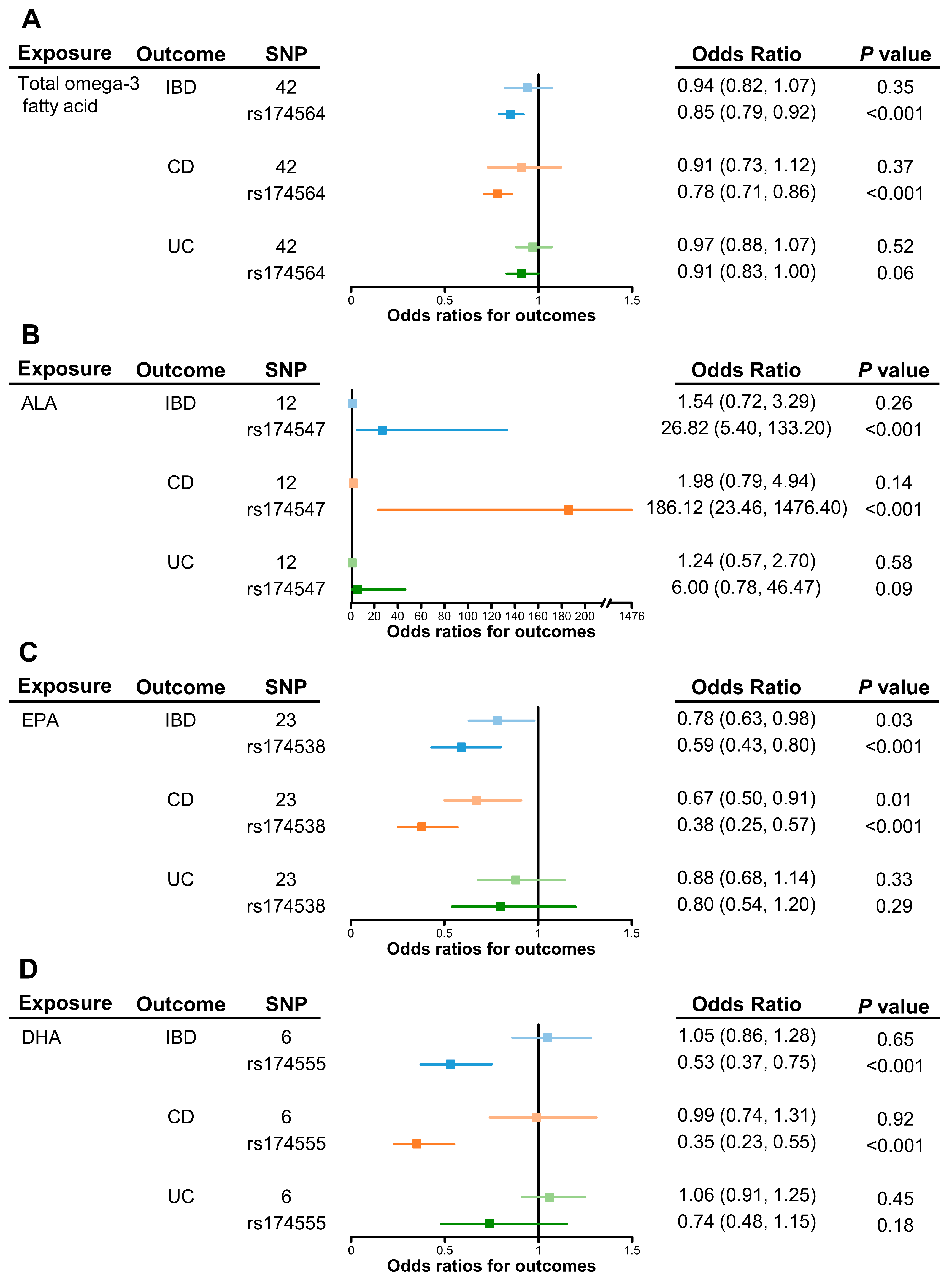
3.1. Genetically Predicted Omega-3 PUFAs on Risk of IBD (Including CD and UC)
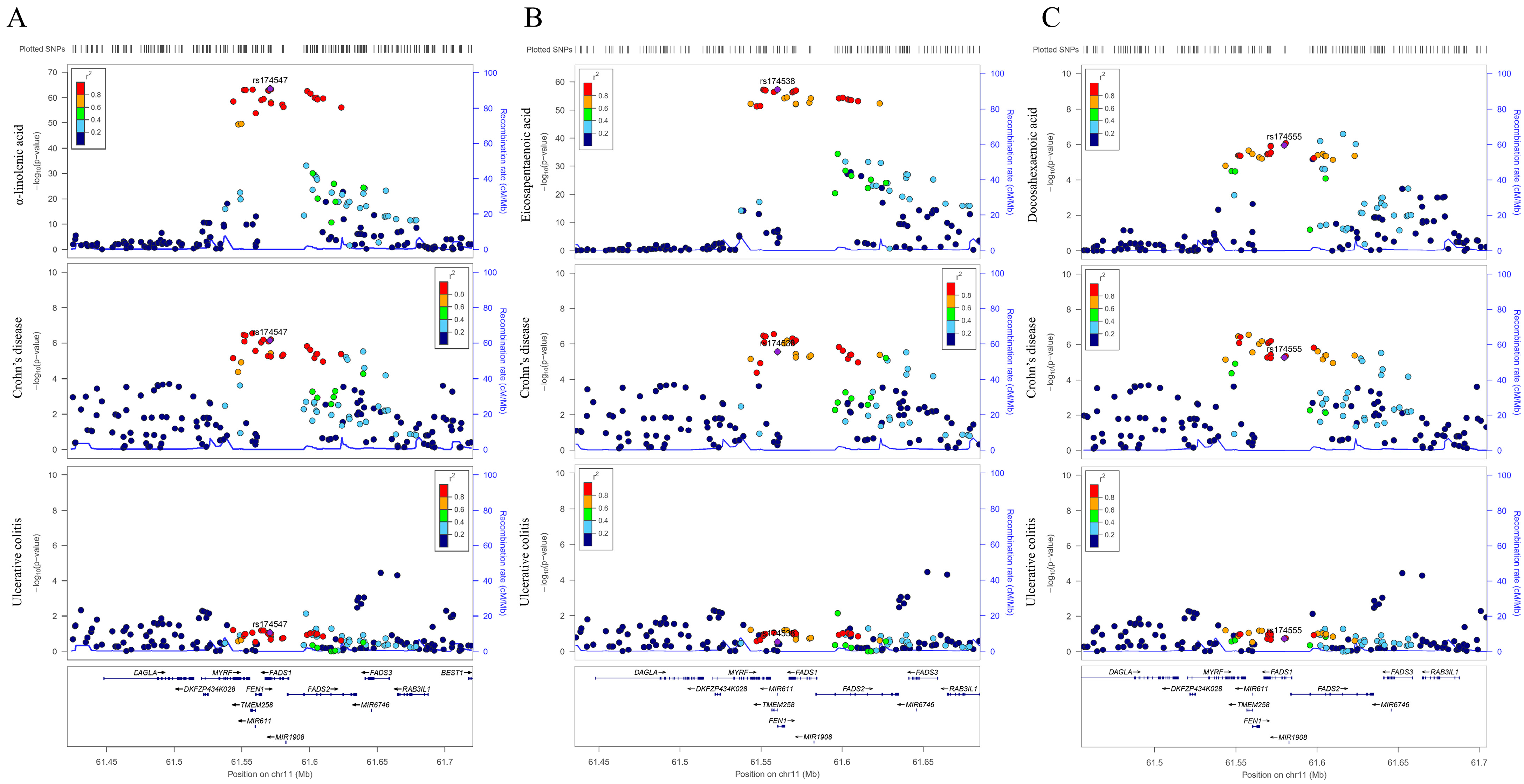
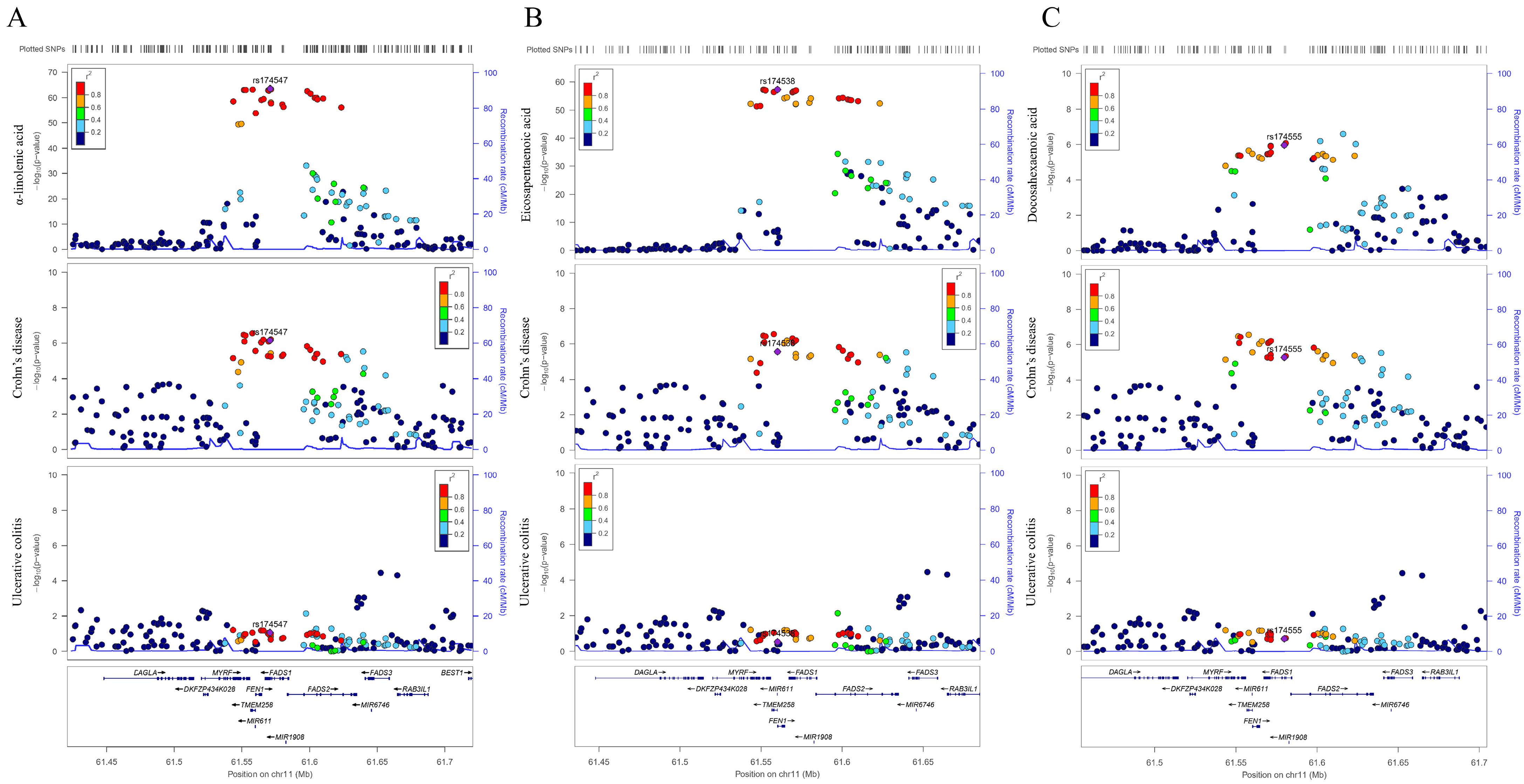
3.2. Sensitivity Analysis in FADS2 Gene Region
3.3. Effects of Genetic Liability to IBD, CD, and UC on the Levels of Omega-3 PUFAs
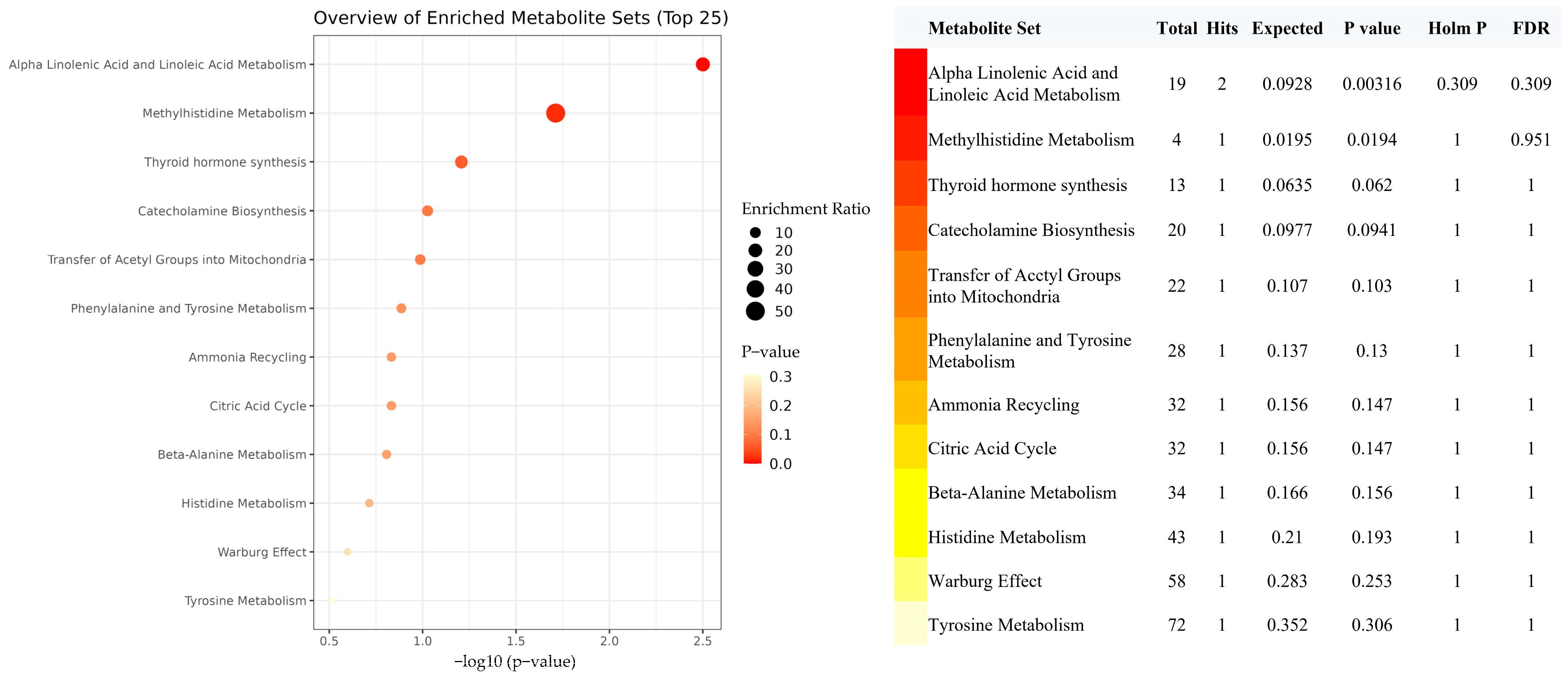
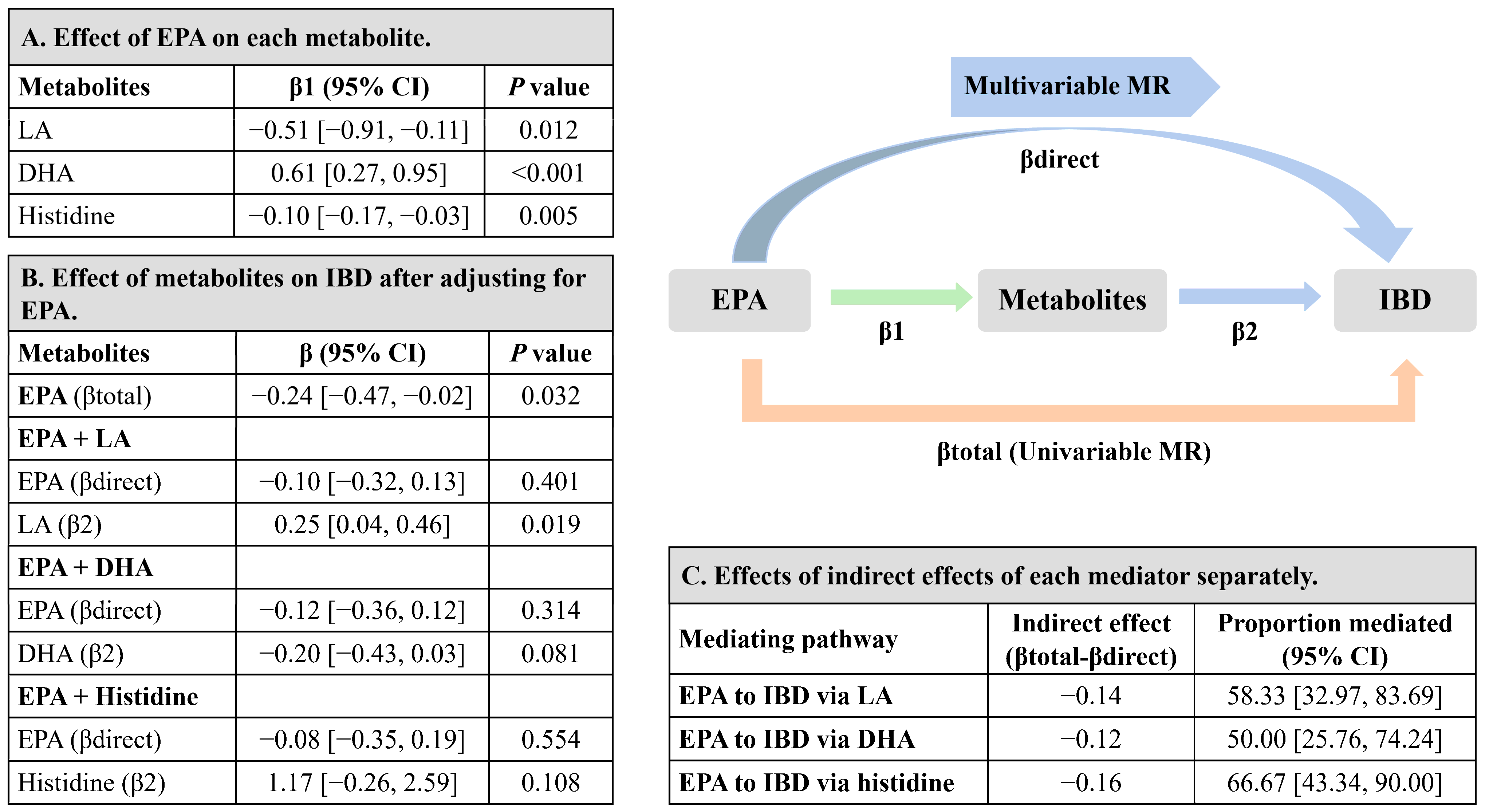
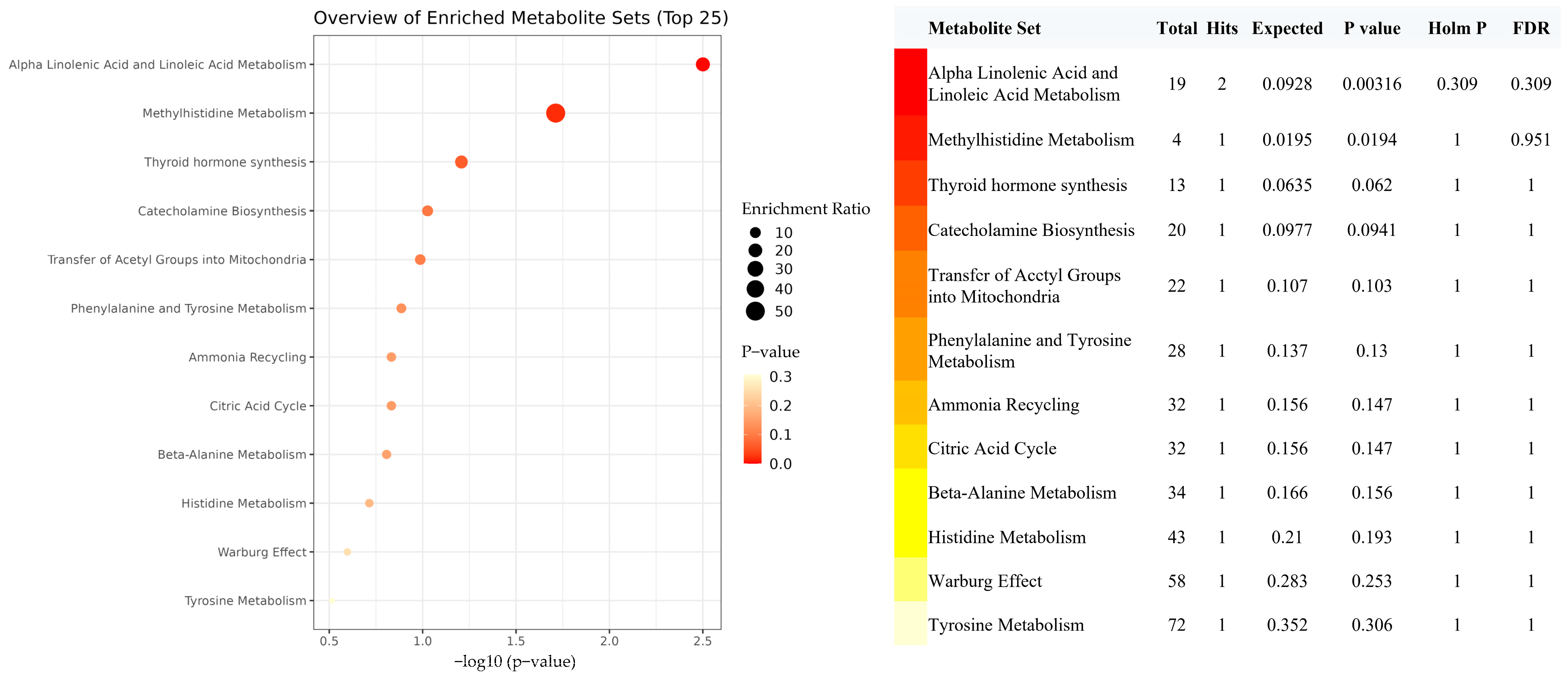
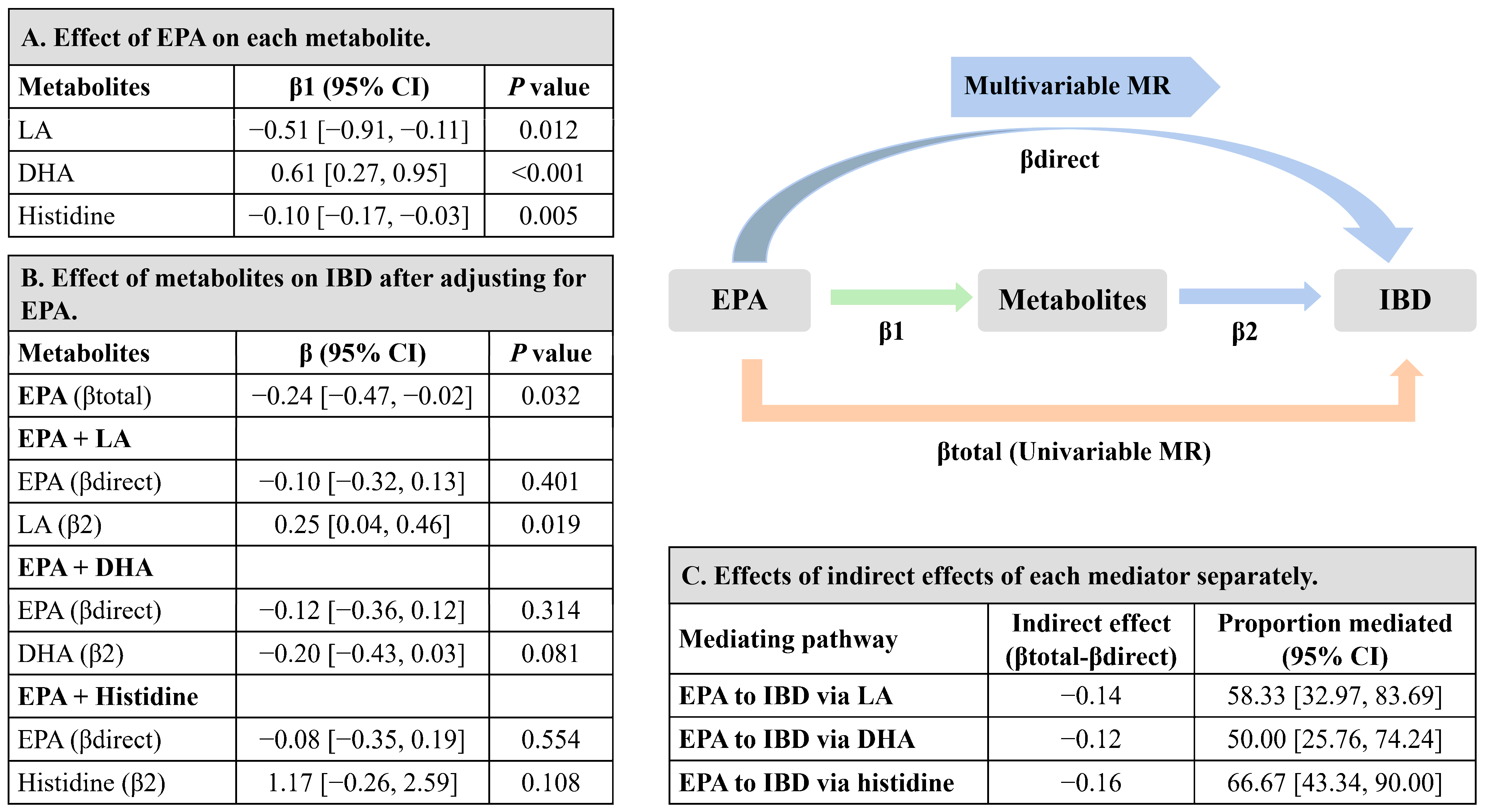
3.4. Mediation MR of EPA, Metabolites, and IBD Risk
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Baumgart, D.C.; Carding, S.R. Inflammatory bowel disease: Cause and immunobiology. Lancet 2007, 369, 1627–1640. [Google Scholar] [CrossRef] [PubMed]
- Kaplan, G.G. The global burden of IBD: From 2015 to 2025. Nat. Rev. Gastroenterol. Hepatol. 2015, 12, 720–727. [Google Scholar] [CrossRef] [PubMed]
- Xavier, R.J.; Podolsky, D.K. Unravelling the pathogenesis of inflammatory bowel disease. Nature 2007, 448, 427–434. [Google Scholar] [CrossRef] [PubMed]
- Del Gobbo, L.C.; Imamura, F.; Aslibekyan, S.; Marklund, M.; Virtanen, J.K.; Wennberg, M.; Yakoob, M.Y.; Chiuve, S.E.; dela Cruz, L.; Frazier-Wood, A.C.; et al. ω-3 Polyunsaturated Fatty Acid Biomarkers and Coronary Heart Disease: Pooling Project of 19 Cohort Studies. JAMA Intern. Med. 2016, 176, 1155–1166. [Google Scholar] [CrossRef]
- Talaei, M.; Sdona, E.; Calder, P.C.; Jones, L.R.; Emmett, P.M.; Granell, R.; Bergström, A.; Melén, E.; Shaheen, S.O. Intake of n-3 polyunsaturated fatty acids in childhood, FADS genotype and incident asthma. Eur. Respir. J. 2021, 58, 2003633. [Google Scholar] [CrossRef]
- Calder, P.C. Polyunsaturated fatty acids, inflammatory processes and inflammatory bowel diseases. Mol. Nutr. Food Res. 2008, 52, 885–897. [Google Scholar] [CrossRef]
- Glaser, C.; Heinrich, J.; Koletzko, B. Role of FADS1 and FADS2 polymorphisms in polyunsaturated fatty acid metabolism. Metabolism 2010, 59, 993–999. [Google Scholar] [CrossRef]
- Gorjão, R.; Azevedo-Martins, A.K.; Rodrigues, H.G.; Abdulkader, F.; Arcisio-Miranda, M.; Procopio, J.; Curi, R. Comparative effects of DHA and EPA on cell function. Pharmacol. Ther. 2009, 122, 56–64. [Google Scholar] [CrossRef]
- Belluzzi, A.; Brignola, C.; Campieri, M.; Pera, A.; Boschi, S.; Miglioli, M. Effect of an enteric-coated fish-oil preparation on relapses in Crohn’s disease. N. Engl. J. Med. 1996, 334, 1557–1560. [Google Scholar] [CrossRef]
- Hahn, J.; Cook, N.R.; Alexander, E.K.; Friedman, S.; Walter, J.; Bubes, V.; Kotler, G.; Lee, I.-M.; Manson, J.E.; Costenbader, K.H. Vitamin D and marine omega 3 fatty acid supplementation and incident autoimmune disease: VITAL randomized controlled trial. BMJ 2022, 376, e066452. [Google Scholar] [CrossRef]
- Chan, S.S.; Luben, R.; Olsen, A.; Tjonneland, A.; Kaaks, R.; Lindgren, S.; Grip, O.; Bergmann, M.M.; Boeing, H.; Hallmans, G.; et al. Association between high dietary intake of the n-3 polyunsaturated fatty acid docosahexaenoic acid and reduced risk of Crohn’s disease. Aliment Pharmacol. Ther. 2014, 39, 834–842. [Google Scholar] [CrossRef] [PubMed]
- Rashvand, S.; Somi, M.H.; Rashidkhani, B.; Hekmatdoost, A. Dietary fatty acid intakes are related to the risk of ulcerative colitis: A case–control study. Int. J. Color. Dis. 2015, 30, 1255–1260. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, Y.; Ohfuji, S.; Kondo, K.; Fukushima, W.; Sasaki, S.; Kamata, N.; Yamagami, H.; Fujiwara, Y.; Suzuki, Y.; Hirota, Y.; et al. Association of Dietary Fatty Acid Intake with the Development of Ulcerative Colitis: A Multicenter Case-Control Study in Japan. Inflamm. Bowel Dis. 2021, 27, 617–628. [Google Scholar] [CrossRef] [PubMed]
- Lawlor, D.A.; Harbord, R.M.; Sterne, J.A.; Timpson, N.; Davey Smith, G. Faculty Opinions recommendation of Mendelian randomization: Using genes as instruments for making causal inferences in epidemiology. Stat. Med. 2008, 27, 1133–1163. [Google Scholar] [CrossRef] [PubMed]
- He, J.; Luo, X.; Xin, H.; Lai, Q.; Zhou, Y.; Bai, Y. The Effects of Fatty Acids on Inflammatory Bowel Disease: A Two-Sample Mendelian Randomization Study. Nutrients 2022, 14, 2883. [Google Scholar] [CrossRef]
- Zuber, V.; Grinberg, N.F.; Gill, D.; Manipur, I.; Slob, E.A.; Patel, A.; Wallace, C.; Burgess, S. Combining evidence from Mendelian randomization and colocalization: Review and comparison of approaches. Am. J. Hum. Genet. 2022, 109, 767–782. [Google Scholar] [CrossRef]
- Borges, M.C.; Haycock, P.C.; Zheng, J.; Hemani, G.; Holmes, M.V.; Davey Smith, G.; Hingorani, A.D.; Lawlor, D.A. Role of circulating polyunsaturated fatty acids on cardiovascular diseases risk: Analysis using Mendelian randomization and fatty acid genetic association data from over 114,000 UK Biobank participants. BMC Med. 2022, 20, 210. [Google Scholar] [CrossRef]
- Lemaitre, R.N.; Tanaka, T.; Tang, W.; Manichaikul, A.; Foy, M.; Kabagambe, E.K.; Nettleton, J.A.; King, I.B.; Weng, L.-C.; Bhattacharya, S.; et al. Genetic loci associated with plasma phospholipid n-3 fatty acids: A meta-analysis of genome-wide association studies from the CHARGE consortium. PLoS Genet. 2011, 7, e1002193. [Google Scholar] [CrossRef]
- de Lange, K.M.; Moutsianas, L.; Lee, J.C.; Lamb, C.A.; Luo, Y.; Kennedy, N.A.; Jostins, L.; Rice, D.L.; Gutierrez-Achury, J.; Ji, S.-G.; et al. Genome-wide association study implicates immune activation of multiple integrin genes in inflammatory bowel disease. Nat. Genet. 2017, 49, 256–261. [Google Scholar] [CrossRef]
- Burgess, S.; Thompson, S.G. Avoiding bias from weak instruments in Mendelian randomization studies. Int. J. Epidemiol. 2011, 40, 755–764. [Google Scholar] [CrossRef]
- Bowden, J.; Smith, G.D.; Haycock, P.C.; Burgess, S. Consistent Estimation in Mendelian Randomization with Some Invalid Instruments Using a Weighted Median Estimator. Genet. Epidemiol. 2016, 40, 304–314. [Google Scholar] [CrossRef]
- Bowden, J.; Davey Smith, G.; Burgess, S. Mendelian randomization with invalid instruments: Effect estimation and bias detection through Egger regression. Int. J. Epidemiol. 2015, 44, 512–525. [Google Scholar] [CrossRef]
- Verbanck, M.; Chen, C.Y.; Neale, B.; Do, R. Detection of widespread horizontal pleiotropy in causal relationships inferred from Mendelian randomization between complex traits and diseases. Nat. Genet. 2018, 50, 693–698. [Google Scholar] [CrossRef]
- Greco, M.F.; Minelli, C.; Sheehan, N.A.; Thompson, J.R. Detecting pleiotropy in Mendelian randomisation studies with summary data and a continuous outcome. Stat. Med. 2015, 34, 2926–2940. [Google Scholar] [CrossRef]
- Hemani, G.; Zheng, J.; Elsworth, B.; Wade, K.H.; Haberland, V.; Baird, D.; Laurin, C.; Burgess, S.; Bowden, J.; Langdon, R.; et al. The MR-Base platform supports systematic causal inference across the human phenome. Elife 2018, 7, e34408. [Google Scholar] [CrossRef]
- Timpson, N.J.; Nordestgaard, B.G.; Harbord, R.M.; Zacho, J.; Frayling, T.M.; Tybjærg-Hansen, A.; Smith, G.D. C-reactive protein levels and body mass index: Elucidating direction of causation through reciprocal Mendelian randomization. Int. J. Obes. 2011, 35, 300–308. [Google Scholar] [CrossRef]
- Carter, A.R.; Sanderson, E.; Hammerton, G.; Richmond, R.C.; Smith, G.D.; Heron, J.; Taylor, A.E.; Davies, N.M.; Howe, L.D. Mendelian randomisation for mediation analysis: Current methods and challenges for implementation. Eur. J. Epidemiol. 2021, 36, 465–478. [Google Scholar] [CrossRef]
- Pang, Z.; Chong, J.; Zhou, G.; de Lima Morais, D.A.; Chang, L.; Barrette, M.; Gauthier, C.; Jacques, P.-É.; Li, S.; Xia, J. MetaboAnalyst 5.0: Narrowing the gap between raw spectra and functional insights. Nucleic Acids Res. 2021, 49, W388–W396. [Google Scholar] [CrossRef]
- Cabré, E.; Mañosa, M.; Gassull, M.A. Omega-3 fatty acids and inflammatory bowel diseases—A systematic review. Br. J. Nutr. 2012, 107 (Suppl. S2), S240–S252. [Google Scholar] [CrossRef]
- Turner, D.; Shah, P.S.; Steinhart, A.H.; Zlotkin, S.; Griffiths, A.M. Maintenance of remission in inflammatory bowel disease using omega-3 fatty acids (fish oil): A systematic review and meta-analyses. Inflamm. Bowel Dis. 2011, 17, 336–345. [Google Scholar] [CrossRef]
- Ananthakrishnan, A.N.; Khalili, H.; Konijeti, G.G.; Higuchi, L.M.; de Silva, P.; Fuchs, C.S.; Willett, W.C.; Richter, J.M.; Chan, A.T. Long-term, intake of dietary fat and risk of ulcerative colitis and Crohn’s disease. Gut 2014, 63, 776–784. [Google Scholar] [CrossRef]
- Calder, P.C.; Yaqoob, P.; Thies, F.; Wallace, F.A.; Miles, E.A. Fatty acids and lymphocyte functions. Br. J. Nutr. 2002, 87 (Suppl. S1), S31–S48. [Google Scholar] [CrossRef]
- Endres, S.; Ghorbani, R.; Kelley, V.E.; Georgilis, K.; Lonnemann, G.; van der Meer, J.W.M.; Cannon, J.G.; Rogers, T.S.; Klempner, M.S.; Weber, P.C.; et al. The effect of dietary supplementation with n-3 polyunsaturated fatty acids on the synthesis of interleukin-1 and tumor necrosis factor by mononuclear cells. N. Engl. J. Med. 1989, 320, 265–271. [Google Scholar] [CrossRef]
- Li, K.; Huang, T.; Zheng, J.; Wu, K.; Li, D. Effect of marine-derived n-3 polyunsaturated fatty acids on C-reactive protein, interleukin 6 and tumor necrosis factor α: A meta-analysis. PLoS ONE 2014, 9, e88103. [Google Scholar] [CrossRef]
- Koletzko, B.; Reischl, E.; Tanjung, C.; Gonzalez-Casanova, I.; Ramakrishnan, U.; Meldrum, S.; Simmer, K.; Heinrich, J.; Demmelmair, H. FADS1 and FADS2 Polymorphisms Modulate Fatty Acid Metabolism and Dietary Impact on Health. Annu. Rev. Nutr. 2019, 39, 21–44. [Google Scholar] [CrossRef]
- Mozaffari, H.; Daneshzad, E.; Larijani, B.; Bellissimo, N.; Azadbakht, L. Dietary intake of fish, n-3 polyunsaturated fatty acids, and risk of inflammatory bowel disease: A systematic review and meta-analysis of observational studies. Eur. J. Nutr. 2020, 59, 1–17. [Google Scholar] [CrossRef]
- Tjonneland, A.; Overvad, K.; Bergmann, M.M.; Nagel, G.; Linseisen, J.; Hallmans, G.; Palmqvist, R.; Sjodin, H.; Hagglund, G.; Berglund, G.; et al. Linoleic acid, a dietary n-6 polyunsaturated fatty acid, and the aetiology of ulcerative colitis: A nested case-control study within a European prospective cohort study. Gut 2009, 58, 1606–1611. [Google Scholar]
- Morin, C.; Blier, P.U.; Fortin, S. Eicosapentaenoic acid and docosapentaenoic acid monoglycerides are more potent than docosahexaenoic acid monoglyceride to resolve inflammation in a rheumatoid arthritis model. Arthritis Res. Ther. 2015, 17, 142. [Google Scholar] [CrossRef]
- Mickleborough, T.D.; Tecklenburg, S.L.; Montgomery, G.S.; Lindley, M.R. Eicosapentaenoic acid is more effective than docosahexaenoic acid in inhibiting proinflammatory mediator production and transcription from LPS-induced human asthmatic alveolar macrophage cells. Clin. Nutr. 2009, 28, 71–77. [Google Scholar] [CrossRef]
- Mozaffarian, D.; Lemaitre, R.N.; King, I.B.; Song, X.; Spiegelman, D.; Sacks, F.M.; Rimm, E.B.; Siscovick, D.S. Circulating long-chain ω-3 fatty acids and incidence of congestive heart failure in older adults: The cardiovascular health study: A cohort study. Ann. Intern. Med. 2011, 155, 160–170. [Google Scholar] [CrossRef]
- Liu, F.; Smith, A.D.; Solano-Aguilar, G.; Wang, T.T.Y.; Pham, Q.; Beshah, E.; Tang, Q.; Urban, J.F.; Xue, C.; Li, R.W. Mechanistic insights into the attenuation of intestinal inflammation and modulation of the gut microbiome by krill oil using in vitro and in vivo models. Microbiome 2020, 8, 83. [Google Scholar] [CrossRef] [PubMed]
- Bosch, S.; Struys, E.A.; van Gaal, N.; Bakkali, A.; Jansen, E.W.; Diederen, K.; Benninga, M.A.; Mulder, C.J.; de Boer, N.K.; de Meij, T.G. Fecal Amino Acid Analysis Can Discriminate de Novo Treatment-Naïve Pediatric Inflammatory Bowel Disease from Controls. J. Pediatr. Gastroenterol. Nutr. 2018, 66, 773–778. [Google Scholar] [CrossRef] [PubMed]
- Ueda, Y.; Kawakami, Y.; Kunii, D.; Okada, H.; Azuma, M.; Le, D.S.N.; Yamamoto, S. Elevated concentrations of linoleic acid in erythrocyte membrane phospholipids in patients with inflammatory bowel disease. Nutr. Res. 2008, 28, 239–244. [Google Scholar] [CrossRef]
- O’Neill, C.M.; Minihane, A.-M. The impact of fatty acid desaturase genotype on fatty acid status and cardiovascular health in adults. Proc. Nutr. Soc. 2017, 76, 64–75. [Google Scholar] [CrossRef]
- Chu, X.; Jaeger, M.; Beumer, J.; Bakker, O.B.; Aguirre-Gamboa, R.; Oosting, M.; Smeekens, S.P.; Moorlag, S.; Mourits, V.P.; Koeken, V.A.C.M.; et al. Integration of metabolomics, genomics, and immune phenotypes reveals the causal roles of metabolites in disease. Genome Biol. 2021, 22, 198. [Google Scholar] [CrossRef] [PubMed]
- Costea, I.; Mack, D.R.; Lemaitre, R.N.; Israel, D.; Marcil, V.; Ahmad, A.; Amre, D.K. Interactions between the dietary polyunsaturated fatty acid ratio and genetic factors determine susceptibility to pediatric Crohn’s disease. Gastroenterology 2014, 146, 929–931. [Google Scholar] [CrossRef]
- Liu, R.; Qiao, S.; Shen, W.; Liu, Y.; Lu, Y.; Liangyu, H.; Guo, Z.; Gong, J.; Shui, G.; Li, Y.; et al. Disturbance of Fatty Acid Desaturation Mediated by FADS2 in Mesenteric Adipocytes Contributes to Chronic Inflammation of Crohn’s Disease. J. Crohn’s Colitis 2020, 14, 1581–1599. [Google Scholar] [CrossRef]
- Turner, D.; Steinhart, A.H.; Griffiths, A.M. Omega 3 fatty acids (fish oil) for maintenance of remission in ulcerative colitis. Cochrane Database Syst. Rev. 2007, 3, Cd006443. [Google Scholar]
- Pearl, D.S.; Masoodi, M.; Eiden, M.; Brümmer, J.; Gullick, D.; McKeever, T.M.; Whittaker, M.A.; Nitch-Smith, H.; Brown, J.F.; Shute, J.K.; et al. Altered colonic mucosal availability of n-3 and n-6 polyunsaturated fatty acids in ulcerative colitis and the relationship to disease activity. J. Crohns Colitis 2014, 8, 70–79. [Google Scholar] [CrossRef]
- Ungaro, F.; Tacconi, C.; Massimino, L.; Corsetto, P.A.; Correale, C.; Fonteyne, P.; Piontini, A.; Garzarelli, V.; Calcaterra, F.; Della Bella, S.; et al. MFSD2A Promotes Endothelial Generation of Inflammation-Resolving Lipid Mediators and Reduces Colitis in Mice. Gastroenterology 2017, 153, 1363–1377.e6. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Exposure | No. of SNPs | Methods | Estimate | Heterogeneity | Pleiotropy | ||||
|---|---|---|---|---|---|---|---|---|---|
| OR | 95% CI | P | Q | Ph | MR Egger int P | MR-PRESSO P | |||
| Total omega-3 fatty acid | 42 | IVW | 0.94 | (0.82, 1.07) | 0.35 | 232.9 | <0.001 | 0.06 | <0.001 |
| MR-Egger | 0.83 | (0.69, 0.99) | 0.05 | ||||||
| Weighted median | 0.85 | (0.80, 0.92) | <0.001 | ||||||
| MR-PRESSO outlier test | 0.88 | (0.81, 0.95) | 0.003 | ||||||
| α-linolenic acid | 12 | IVW | 1.54 | (0.72, 3.29) | 0.26 | 46.7 | <0.001 | 0.65 | <0.001 |
| MR-Egger | 1.40 | (0.58, 3.39) | 0.48 | ||||||
| Weighted median | 1.42 | (0.89, 2.28) | 0.14 | ||||||
| MR-PRESSO outlier test | 1.24 | (0.79, 1.95) | 0.38 | ||||||
| EPA | 23 | IVW | 0.78 | (0.63, 0.98) | 0.03 | 30.8 | 0.099 | 0.97 | 0.099 |
| MR-Egger | 0.78 | (0.45, 1.34) | 0.37 | ||||||
| Weighted median | 0.59 | (0.45, 0.78) | <0.001 | ||||||
| MR-PRESSO outlier test | NA | NA | NA | ||||||
| DHA | 6 | IVW | 1.05 | (0.86, 1.28) | 0.65 | 21.6 | <0.001 | 0.56 | 0.012 |
| MR-Egger | 1.20 | (0.75, 1.93) | 0.49 | ||||||
| Weighted median | 1.12 | (0.98, 1.28) | 0.09 | ||||||
| MR-PRESSO outlier test | 1.11 | (0.99, 1.25) | 0.43 | ||||||
| Exposure | No. of SNPs | Outcome | No. of SNPs | IVW | Heterogeneity | Pleiotropy | ||||
|---|---|---|---|---|---|---|---|---|---|---|
| Beta | 95% CI | P | Q | Ph | MR Egger int P | MR-PRESSOP | ||||
| IBD | 117 | Total omega-3 fatty acid | 105 | −0.002 | (−0.012, 0.009) | 0.76 | 200.7 | <0.001 | 0.86 | <0.001 |
| α-linolenic acid | 39 | −0.001 | (−0.004, 0.001) | 0.36 | 35.9 | 0.57 | 0.94 | 0.416 | ||
| EPA | 39 | 0.001 | (−0.019, 0.020) | 0.92 | 57.9 | 0.02 | 0.87 | 0.009 | ||
| DHA | 39 | −0.010 | (−0.059, 0.040) | 0.71 | 47.7 | 0.13 | 0.69 | 0.036 | ||
| CD | 89 | Total omega-3 fatty acid | 83 | 0.004 | (−0.005, 0.013) | 0.39 | 174.5 | <0.001 | 0.28 | <0.001 |
| α-linolenic acid | 28 | −0.001 | (−0.003, 0.001) | 0.28 | 28.7 | 0.38 | 0.41 | 0.463 | ||
| EPA | 28 | 0.011 | (−0.005, 0.026) | 0.17 | 46.4 | 0.01 | 0.85 | 0.012 | ||
| DHA | 28 | 0.029 | (−0.011, 0.070) | 0.15 | 39.3 | 0.06 | 0.81 | 0.090 | ||
| UC | 62 | Total omega-3 fatty acid | 53 | −0.005 | (−0.018, 0.008) | 0.45 | 131.0 | <0.001 | 0.27 | <0.001 |
| α-linolenic acid | 27 | 0.002 | (−0.001, 0.004) | 0.23 | 28.5 | 0.34 | 0.34 | 0.446 | ||
| EPA | 27 | −0.005 | (−0.021, 0.010) | 0.50 | 26.5 | 0.44 | 0.38 | 0.088 | ||
| DHA | 27 | −0.045 | (−0.089, −0.002) | 0.04 | 26.7 | 0.42 | 0.61 | 0.095 | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jia, X.; Hu, C.; Wu, X.; Qi, H.; Lin, L.; Xu, M.; Xu, Y.; Wang, T.; Zhao, Z.; Chen, Y.; et al. Evaluating the Effects of Omega-3 Polyunsaturated Fatty Acids on Inflammatory Bowel Disease via Circulating Metabolites: A Mediation Mendelian Randomization Study. Metabolites 2023, 13, 1041. https://doi.org/10.3390/metabo13101041
Jia X, Hu C, Wu X, Qi H, Lin L, Xu M, Xu Y, Wang T, Zhao Z, Chen Y, et al. Evaluating the Effects of Omega-3 Polyunsaturated Fatty Acids on Inflammatory Bowel Disease via Circulating Metabolites: A Mediation Mendelian Randomization Study. Metabolites. 2023; 13(10):1041. https://doi.org/10.3390/metabo13101041
Chicago/Turabian StyleJia, Xiaojing, Chunyan Hu, Xueyan Wu, Hongyan Qi, Lin Lin, Min Xu, Yu Xu, Tiange Wang, Zhiyun Zhao, Yuhong Chen, and et al. 2023. "Evaluating the Effects of Omega-3 Polyunsaturated Fatty Acids on Inflammatory Bowel Disease via Circulating Metabolites: A Mediation Mendelian Randomization Study" Metabolites 13, no. 10: 1041. https://doi.org/10.3390/metabo13101041
APA StyleJia, X., Hu, C., Wu, X., Qi, H., Lin, L., Xu, M., Xu, Y., Wang, T., Zhao, Z., Chen, Y., Li, M., Zheng, R., Lin, H., Wang, S., Wang, W., Bi, Y., Zheng, J., & Lu, J. (2023). Evaluating the Effects of Omega-3 Polyunsaturated Fatty Acids on Inflammatory Bowel Disease via Circulating Metabolites: A Mediation Mendelian Randomization Study. Metabolites, 13(10), 1041. https://doi.org/10.3390/metabo13101041