
Interaction between Plasma Metabolomics and Intestinal Microbiome in db/db Mouse, an Animal Model for Study of Type 2 Diabetes and Diabetic Kidney Disease



Abstract
:1. Introduction
2. Results
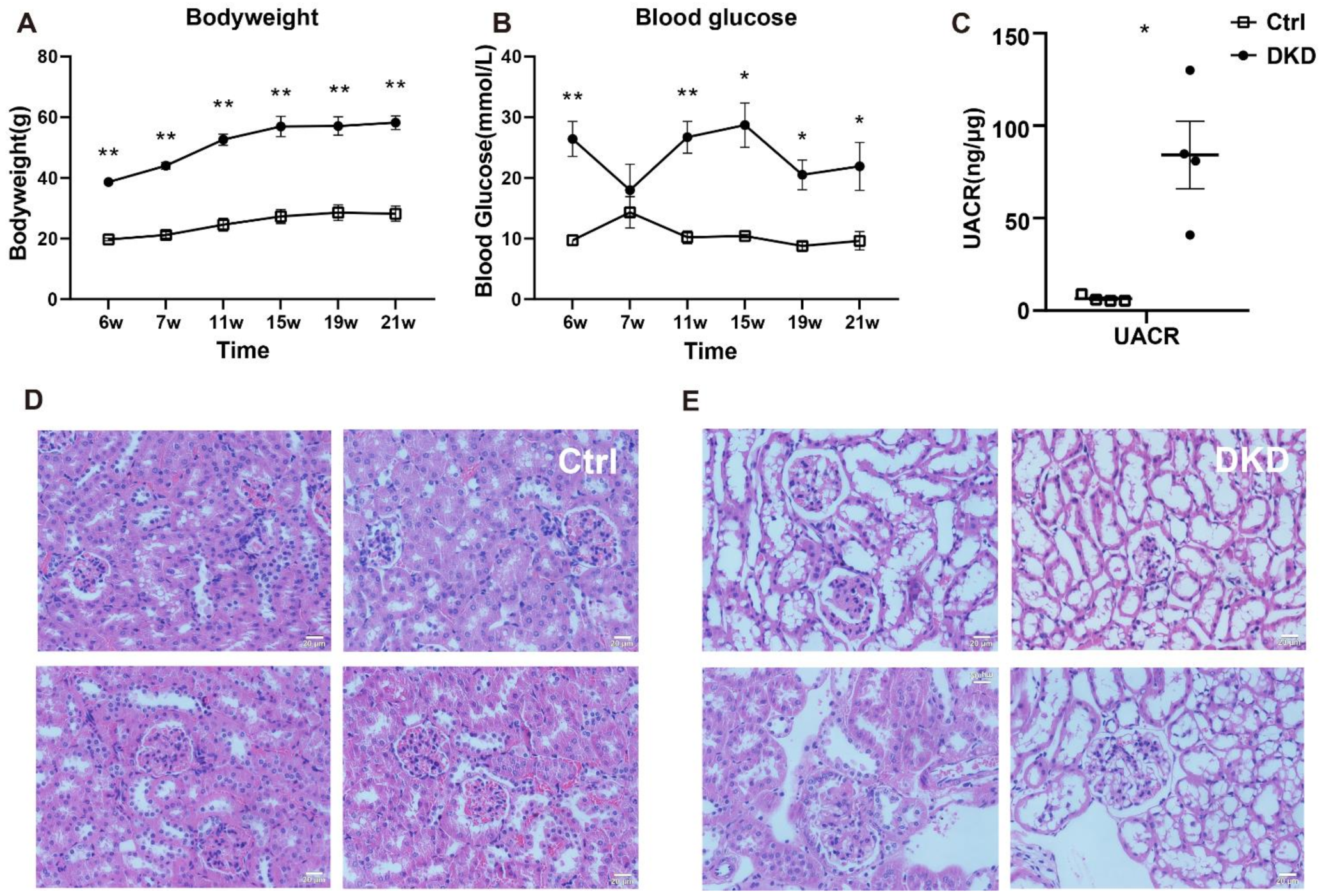
2.1. Basic Physio-Pathological Indicators
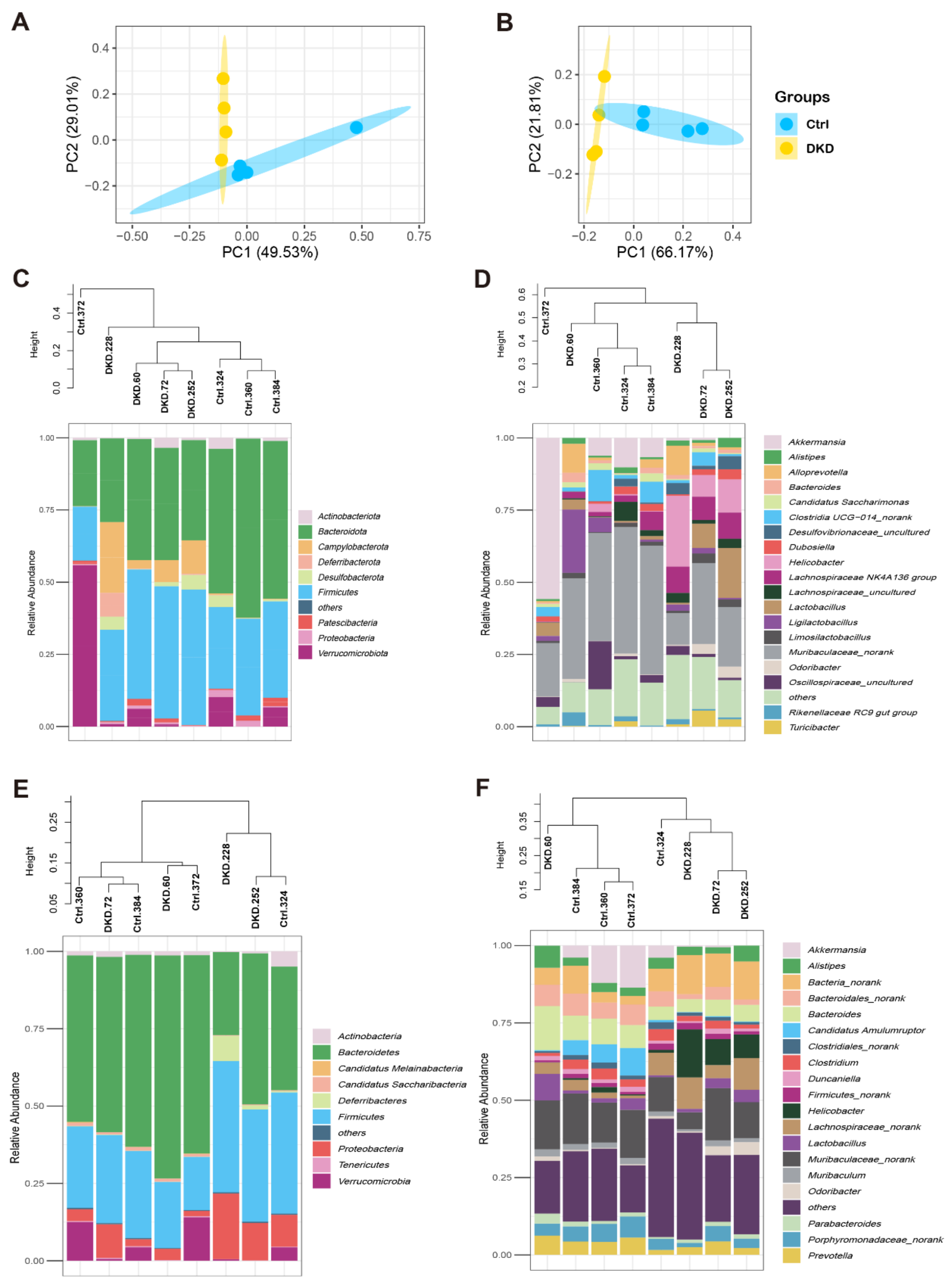
2.2. Identification of Intestinal Prokaryotes
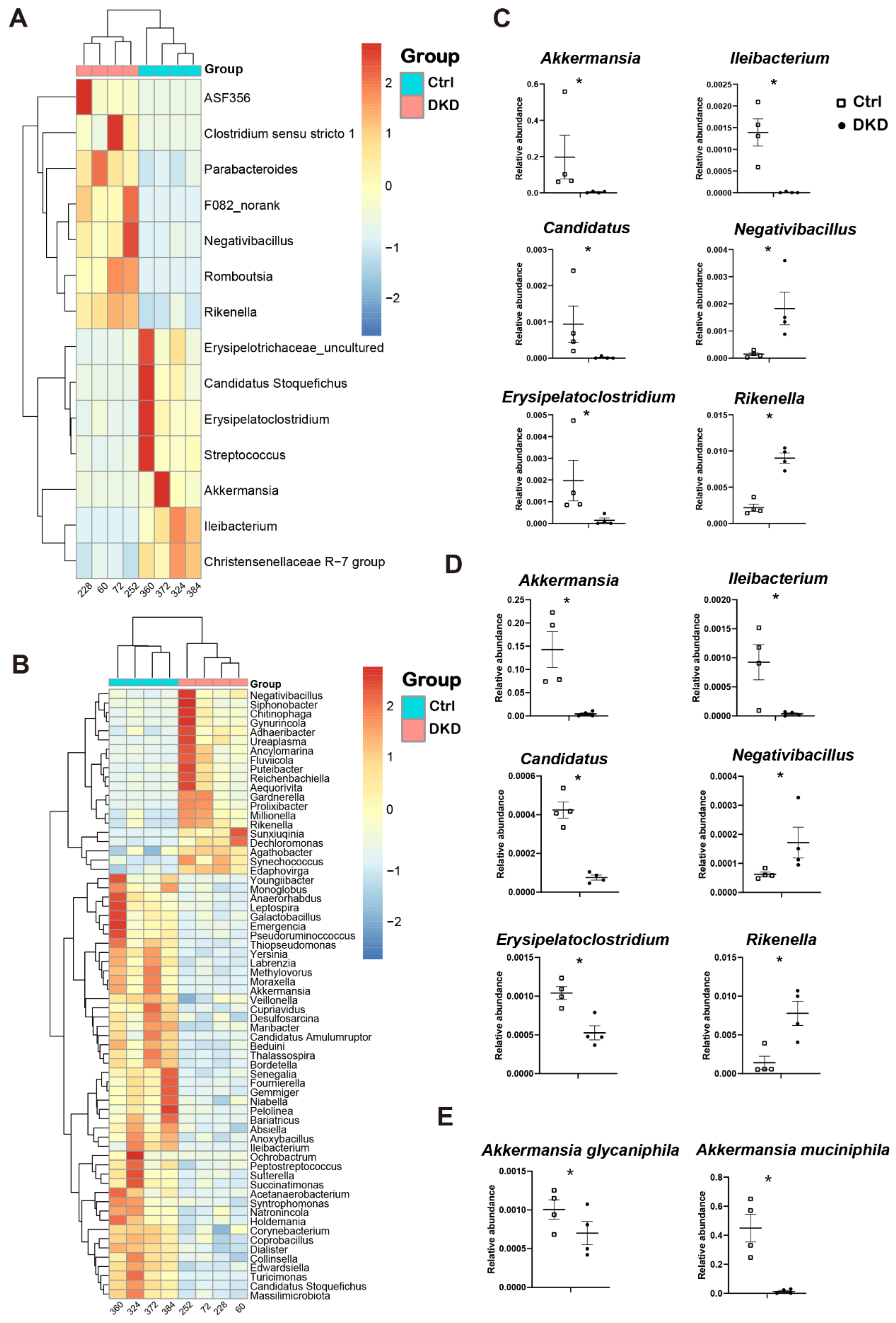
2.3. The Genus Levels of Microbes in db/db Mice of DKD Group
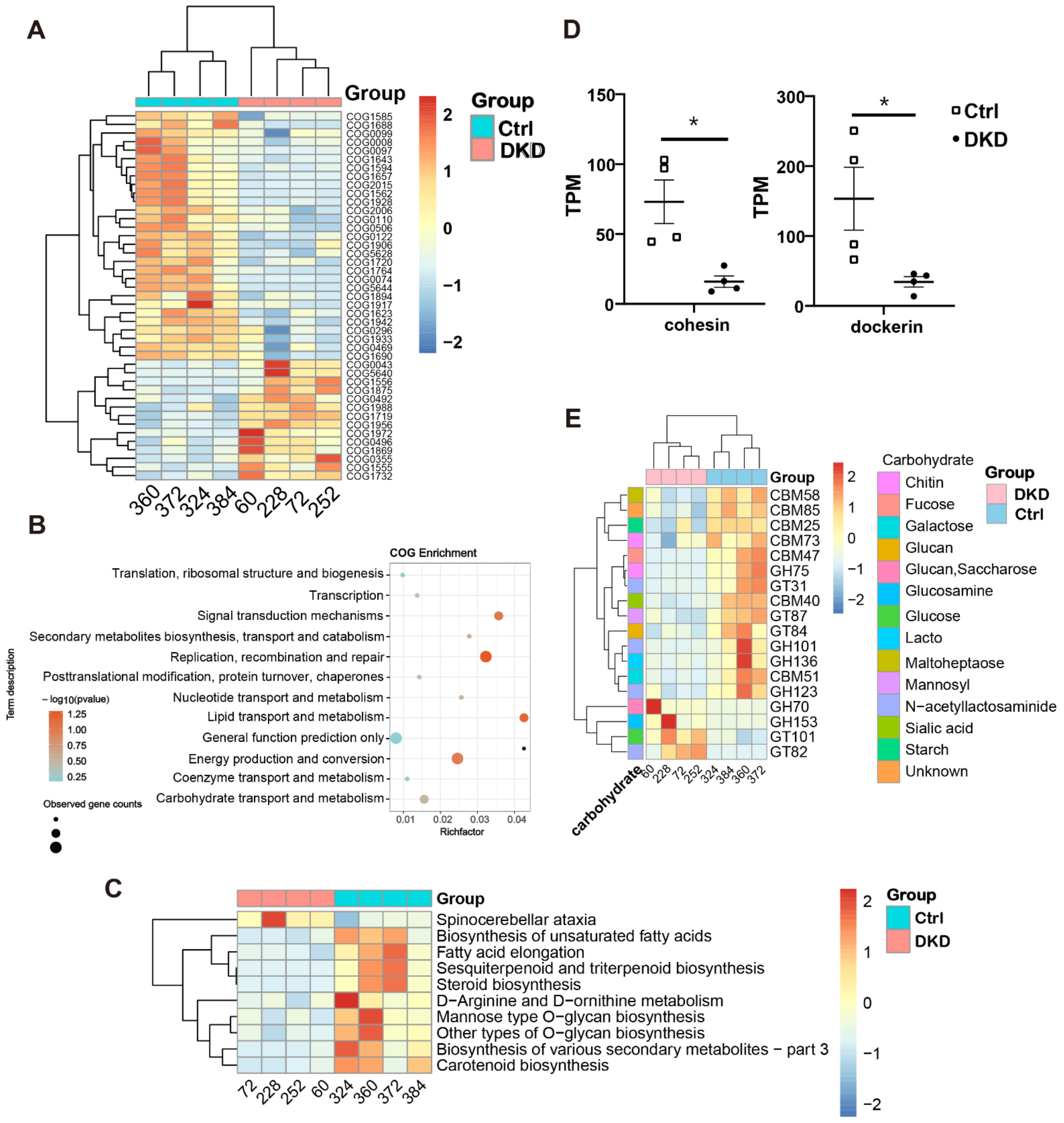
2.4. Intergroup Differences in Functional Annotations
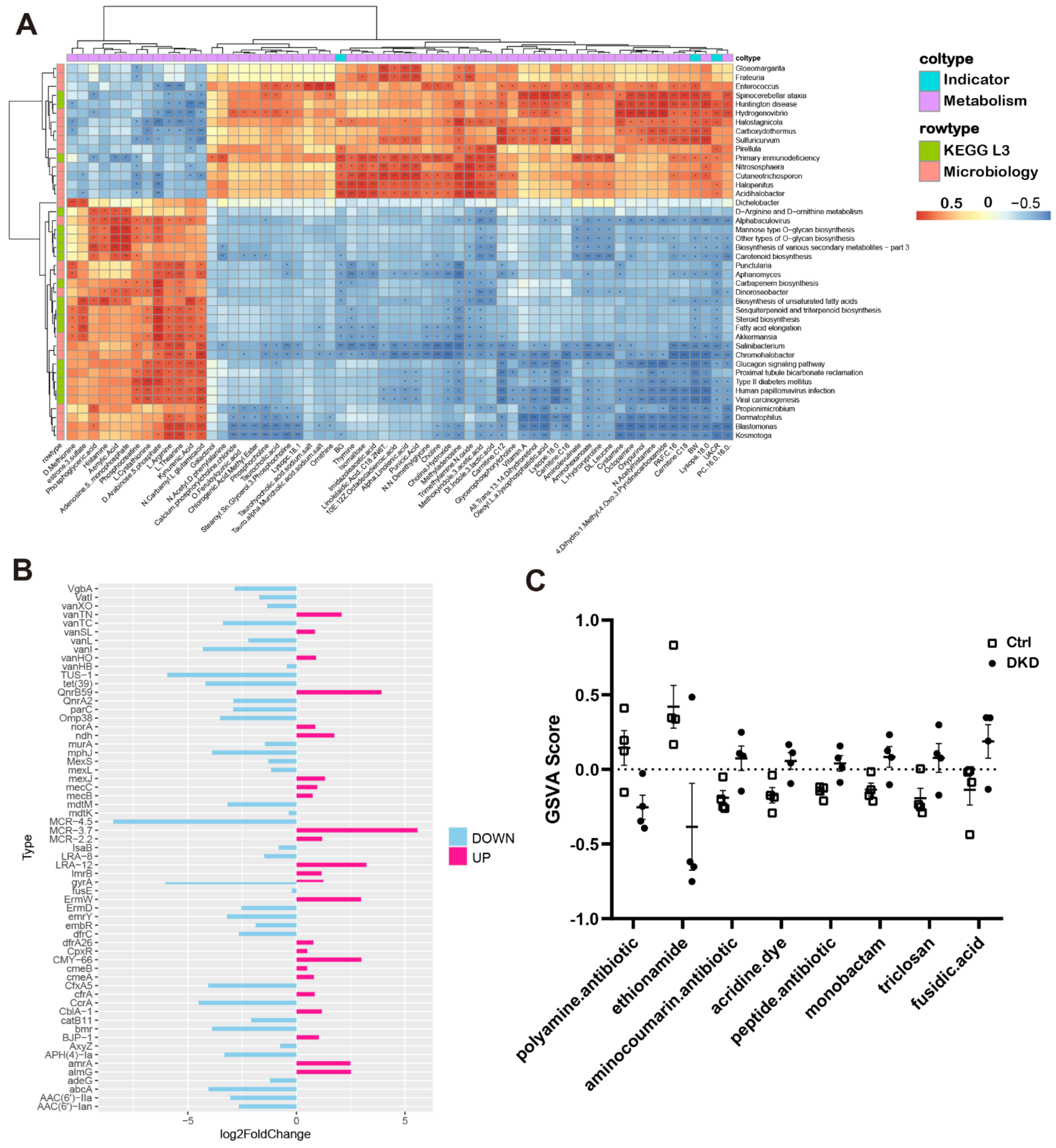
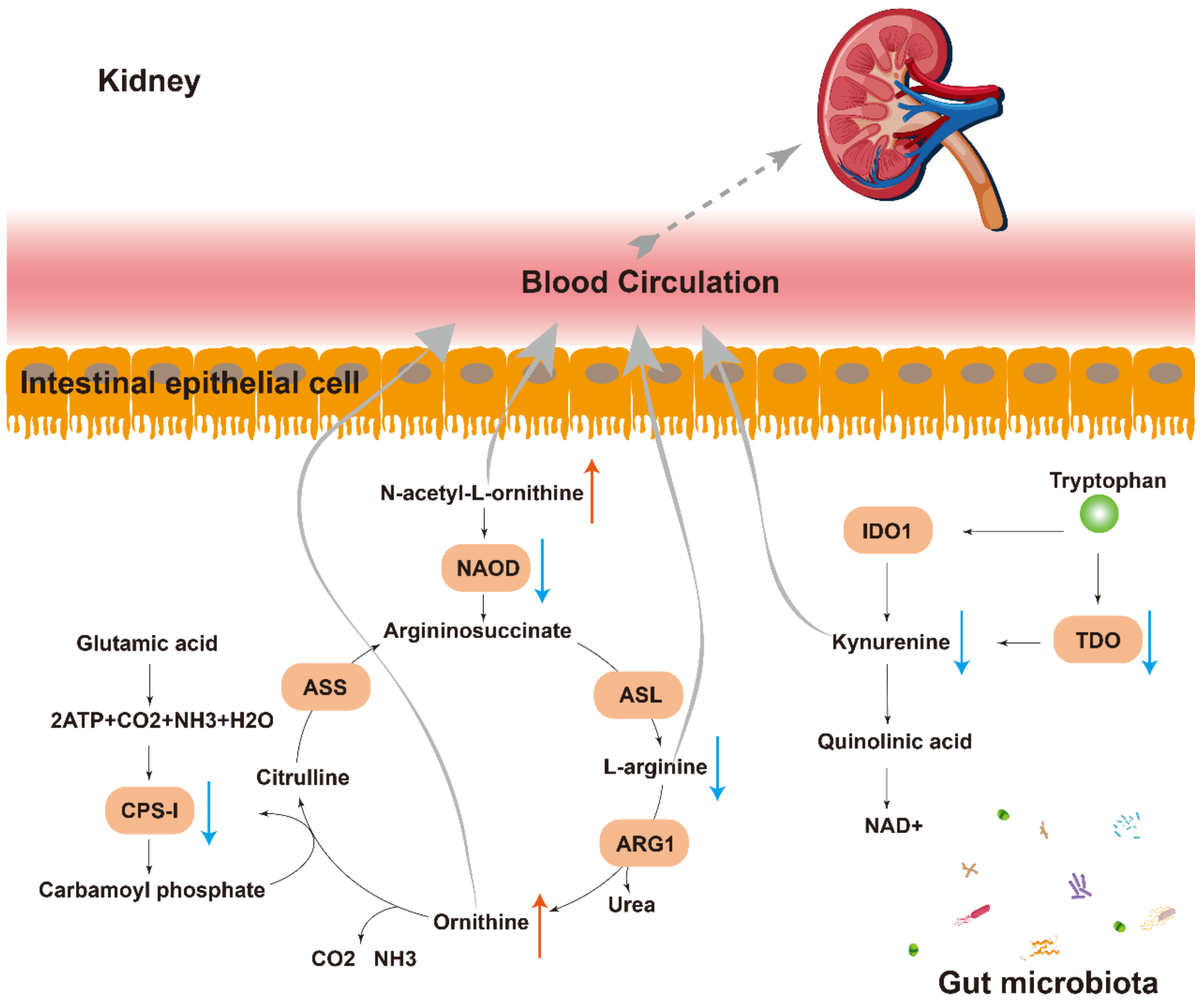
2.5. Interaction between Plasma Metabolomics and Intestinal Microbiome
3. Discussion
4. Material and Methods
4.1. Animals and Physio-Pathological Parameters
4.2. Metabolomic Analysis with Plasma Samples
4.3. Precondition of 16S rDNA and Metagenomics Sequencing
4.4. Bioinformatical and Statistical Analyses
Supplementary Materials
Author Contributions
Funding
Institutional Review Bord Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| 7-KCHO | 7-Ketocholesterol |
| AGE | agarose gel electrophoresis |
| AMR | antimicrobial resistance |
| ARO | antibiotic resistance ontology |
| CARD | comprehensive antibiotic resistance database |
| CAZy | carbohydrate-active enzymes databases |
| COG | clusters of orthologous groups of proteins |
| COXs | cyclooxygenases |
| CPAI | chromatographic peak area integration |
| Cr | creatinine |
| DKD | diabetic kidney disease |
| ECM | extracellular matrix |
| ESKD | end-stage kidney disease |
| G-3-P | glycerol-3-phosphate |
| GBM | glomerular basement membrane |
| HE | hematoxylin-eosin |
| HMC | human mesangial cells |
| IDF | International Diabetes Federation |
| LC-MS | liquid mass spectrometry |
| L-Kyn | L-Kynureniure |
| LOXs | lipoxygenases |
| LPAR | lysophosphatidic acid receptor |
| Lysopa | lysophosphatidic acid |
| Lysopc | lysophosphatidyl choline |
| Lysope | lysophosphatidyl ethanolamine |
| Lysops | lysophosphatidyl serine |
| MAU | microalbuminuria |
| MS | mass spectrometry |
| NAOD | N2-acetylornithine deacetylase |
| NMR | nuclear magnetic resonance spectroscopy |
| novoDB | novogene database |
| NR | non-redundant protein sequence database |
| PCA | principal component analysis |
| PE | pair-end |
| QC | quality control |
| RAAS | renin-angiotensin-aldosterone system |
| RDP | ribosomal database project |
| SCFA | short-chain fatty acids |
| TDO | tryptophan 2,3-dioxygenase |
| UACR | urinary albumin/creatinine ratio |
References
- Saeedi, P.; Petersohn, I.; Salpea, P.; Malanda, B.; Karuranga, S.; Unwin, N.; Colagiuri, S.; Guariguata, L.; Motala, A.A.; Ogurtsova, K.; et al. Global and regional diabetes prevalence estimates for 2019 and projections for 2030 and 2045: Results from the International Diabetes Federation Diabetes Atlas, 9th edition. Diabetes Res. Clin. Pract. 2019, 157, 107843. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ebrahimzadeh Leylabadlo, H.; Sanaie, S.; Sadeghpour Heravi, F.; Ahmadian, Z.; Ghotaslou, R. From role of gut microbiota to microbial-based therapies in type 2-diabetes. Infect. Genet. Evol. 2020, 81, 104268. [Google Scholar] [CrossRef] [PubMed]
- Ojo, O.; Feng, Q.Q.; Ojo, O.O.; Wang, X.H. The Role of Dietary Fibre in Modulating Gut Microbiota Dysbiosis in Patients with Type Diabetes: A Systematic Review and Meta-Analysis of Randomised Controlled Trials. Nutrients 2020, 12, 3239. [Google Scholar] [CrossRef] [PubMed]
- Sharma, S.; Tripathi, P. Gut microbiome and type 2 diabetes: Where we are and where to go? J. Nutr. Biochem. 2019, 63, 101–108. [Google Scholar] [CrossRef]
- Thomas, M.C.; Brownlee, M.; Susztak, K.; Sharma, K.; Jandeleit-Dahm, K.A.; Zoungas, S.; Rossing, P.; Groop, P.H.; Cooper, M.E. Diabetic kidney disease. Nat. Rev. Dis. Primers 2015, 1, 15018. [Google Scholar] [CrossRef]
- Cole, J.B.; Florez, J.C. Genetics of diabetes mellitus and diabetes complications. Nat. Rev. Nephrol. 2020, 16, 377–390. [Google Scholar] [CrossRef]
- Doshi, S.M.; Friedman, A.N. Diagnosis and Management of Type 2 Diabetic Kidney Disease. Clin. J. Am. Soc. Nephrol. 2017, 12, 1366–1373. [Google Scholar] [CrossRef]
- Pugliese, G.; Penno, G.; Natali, A.; Barutta, F.; Di Paolo, S.; Reboldi, G.; Gesualdo, L.; De Nicola, L.; Italian Diabetes Society and the Italian Society of Nephrology. Diabetic kidney disease: New clinical and therapeutic issues. Joint position statement of the Italian Diabetes Society and the Italian Society of Nephrology on “The natural history of diabetic kidney disease and treatment of hyperglycemia in patients with type 2 diabetes and impaired renal function”. Nutr. Metab. Cardiovasc. Dis. 2019, 29, 1127–1150. [Google Scholar]
- De Boer, I.H.; Caramori, M.L.; Chan, J.C.N.; Heerspink, H.J.L.; Hurst, C.; Khunti, K.; Liew, A.; Michos, E.D.; Navaneethan, S.D.; Olowu, W.A.; et al. Executive summary of the 2020 KDIGO Diabetes Management in CKD Guideline: Evidence-based advances in monitoring and treatment. Kidney Int. 2020, 98, 839–848. [Google Scholar] [CrossRef]
- Wang, B.; Chandrasekera, P.C.; Pippin, J.J. Leptin- and leptin receptor-deficient rodent models: Relevance for human type 2 dia-betes. Curr. Diabetes Rev. 2014, 10, 131–145. [Google Scholar] [CrossRef] [Green Version]
- Sharma, K.; McCue, P.; Dunn, S.R. Diabetic kidney disease in the db/db mouse. Am. J. Physiol. Renal Physiol. 2003, 284, F1138–F1144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, M.; Yang, Z.; Zhang, C.; Shi, Y.; Han, W.; Song, S.; Mu, L.; Du, C.; Shi, Y. Inhibition of NLRP3 inflammasome ameliorates podocyte damage by suppressing lipid accumulation in diabetic nephropathy. Metabolism 2021, 118, 154748. [Google Scholar] [CrossRef] [PubMed]
- Alicic, R.Z.; Rooney, M.T.; Tuttle, K.R. Diabetic Kidney Disease: Challenges, Progress, and Possibilities. Clin. J. Am. Soc. Nephrol. 2017, 12, 2032–2045. [Google Scholar] [CrossRef] [PubMed]
- Singer-Englar, T.; Barlow, G.; Mathur, R. Obesity, diabetes, and the gut microbiome: An updated review. Expert Rev. Gastroenterol. Hepatol. 2018, 13, 3–15. [Google Scholar] [CrossRef]
- Tsai, H.-J.; Tsai, W.-C.; Hung, W.-C.; Hung, W.-W.; Chang, C.-C.; Dai, C.-Y.; Tsai, Y.-C. Gut Microbiota and Subclinical Cardiovascular Disease in Patients with Type 2 Diabetes Mellitus. Nutrients 2021, 13, 2679. [Google Scholar] [CrossRef]
- Liu, Y.-X.; Qin, Y.; Chen, T.; Lu, M.; Qian, X.; Guo, X.; Bai, Y. A practical guide to amplicon and metagenomic analysis of microbiome data. Protein Cell 2020, 12, 315–330. [Google Scholar] [CrossRef]
- Galperin, M.Y.; Wolf, Y.I.; Makarova, K.S.; Alvarez, R.V.; Landsman, D.; Koonin, E.V. COG database update: Focus on microbial diversity, model organisms, and widespread pathogens. Nucleic Acids Res. 2020, 49, D274–D281. [Google Scholar] [CrossRef]
- Drula, E.; Garron, M.-L.; Dogan, S.; Lombard, V.; Henrissat, B.; Terrapon, N. The carbohydrate-active enzyme database: Functions and literature. Nucleic Acids Res. 2021, 50, D571–D577. [Google Scholar] [CrossRef]
- Alcock, B.P.; Raphenya, A.R.; Lau, T.T.Y.; Tsang, K.K.; Bouchard, M.; Edalatmand, A.; Huynh, W.; Nguyen, A.-L.V.; Cheng, A.A.; Liu, S.; et al. CARD 2020: Antibiotic resistome surveillance with the comprehensive antibiotic resistance database. Nucleic Acids Res. 2020, 48, D517–D525. [Google Scholar] [CrossRef]
- Yu, G.; Wang, L.-G.; Han, Y.; He, Q.-Y. clusterProfiler: An R Package for Comparing Biological Themes Among Gene Clusters. OMICS J. Integr. Biol. 2012, 16, 284–287. [Google Scholar] [CrossRef]
- Hänzelmann, S.; Castelo, R.; Guinney, J. GSVA: Gene set variation analysis for microarray and RNA-Seq data. BMC Bioinform. 2013, 14, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Depommier, C.; Everard, A.; Druart, C.; Plovier, H.; Van Hul, M.; Vieira-Silva, S.; Falony, G.; Raes, J.; Maiter, D.; Delzenne, N.M.; et al. Supplementation with Akkermansia muciniphila in over-weight and obese human volunteers: A proof-of-concept exploratory study. Nat. Med. 2019, 25, 1096–1103. [Google Scholar] [CrossRef] [PubMed]
- Kim, E.; Kim, A.H.; Lee, Y.; Ji, S.C.; Cho, J.; Yu, K.; Chung, J. Effects of vancomycin-induced gut microbiome alteration on the pharmacodynamics of metformin in healthy male subjects. Clin. Transl. Sci. 2021, 14, 1955–1966. [Google Scholar] [CrossRef] [PubMed]
- Wu, M.H.; Lin, C.N.; Chiu, D.T.; Chen, S.T. Kynurenine/Tryptophan Ratio Predicts Angiotensin Receptor Blocker Responsiveness in Patients with Diabetic Kidney Disease. Diagnostics 2020, 10, 20. [Google Scholar] [CrossRef] [Green Version]
- Wu, C.; Tao, Y.; Li, N.; Fei, J.; Wang, Y.; Wu, J.; Gu, H.F. Prediction of cellular targets in diabetic kidney diseases with single-cell transcriptomic analysis of db/db mouse kidneys. J. Cell Commun. Signal. 2022, 1–20. [Google Scholar] [CrossRef]
- Watanabe, Y.; Yamaguchi, T.; Ishihara, N.; Nakamura, S.; Tanaka, S.; Oka, R.; Imamura, H.; Sato, Y.; Ban, N.; Kawana, H.; et al. 7-Ketocholesterol induces ROS-mediated mRNA expression of 12-lipoxygenase, cyclooxygenase-2 and pro-inflammatory cytokines in human mesangial cells: Potential role in diabetic nephropathy. Prostaglandins Other Lipid Mediat. 2017, 134, 16–23. [Google Scholar] [CrossRef]
- Lyons, M.A.; Brown, A.J. 7-Ketocholesterol. Int. J. Biochem. Cell Biol. 1999, 31, 369–375. [Google Scholar] [CrossRef]
- Devi, S.; Nongkhlaw, B.; Limesh, M.; Pasanna, R.M.; Thomas, T.; Kuriyan, R.; Kurpad, A.V.; Mukhopadhyay, A. Acyl ethanolamides in Diabetes and Diabetic Nephropathy: Novel targets from untargeted plasma metabolomic profiles of South Asian Indian men. Sci. Rep. 2019, 9, 18117. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Metabolites | Classification | Roles | Mechanisms | Regulation in Current Study |
|---|---|---|---|---|
| γ-Glutamyl glutamic | Amino acid derivative | Alleviation | It may enhance mitochondrial metabolism and insulin secretion. | Down |
| N-Carbamyl-L-glutamicacid | Amino acid derivative | Alleviation | Activates the urea cycle to prevent high blood ammonia. | Down |
| L-Methionine | Amino acid derivative | Alleviation/Deterioration | Moderate intake can maintain glucose and lipid metabolism homeostasis in T2D. However, excessive intake can lead to increased insulin resistance, oxidative stress and inflammation. | Down |
| D-Methionine | Amino acid derivative | Alleviation | Protects the kidneys through antioxidants. | Down |
| L-arginine | Amino acid derivative | Alleviation | Protects renal endothelial cells by participating in nitric oxide synthesis, subsequently increasing glomerular filtration rate and tubular reabsorption and reducing proteinuria. | Down |
| L-theanine | Amino acid derivative | Alleviation | L-theanine relieves liver and kidney damage by reducing oxidative stress, inflammatory response and apoptosis. | Down |
| Glu-Glu | Amino acid derivative | Alleviation | Similar to Glutamyl glutamic. | Down |
| L-Cystathionine | Amino acid derivative | Alleviation | L-cystathionine can inhibit mitochondria-dependent apoptosis. | Down |
| L-Kynurenine | Amino acid derivative | Alleviation | L-kynurenine can hinder oxidative stress and immune response. | Down |
| Histamine | Amino acid derivative | Alleviation/Deterioration | Although histamine is a well-known inflammatory mediator, there is also evidence that it may reduce kidney damage in glomerular basement membrane glomerulonephritis. | Down |
| 3-Aminoisobutyric acid | Amino acid derivative | Alleviation | It has several effects, including improving inflammation, insulin resistance, glucose homeostasis and lipid metabolism. | Up |
| N-acetyl-L-ornithine | Amino acid derivative | Deterioration | The content of N-acetyl-L-ornithine in T2D patients with DKD increased significantly, and it is an important progressive factor of DKD. | Up |
| Ornithine | Amino acid derivative | Indirect effects | Polyamines formed after ornithine decarboxylation led to renal hypertrophy. | Up |
| L-Homocitrulline | Amino acid derivative | Deterioration | The formation of L-homocitrulline produces ammonia, which triggers cytotoxicity of macrophages. | Up |
| L-Hydroxylysine | Amino acid derivative | Deterioration | Excessive amounts of L-hydroxylysine can cause collagen deposition, which leads to thickening of the glomerular basement membrane. | Up |
| DL-Valine | Amino acid derivative | Marker | It is a marker of DKD mouse plasma induced by STZ. | Up |
| DL-Leucine | Amino acid derivative | Marker | Like DL-valine, it can be identified as a marker of DKD. | Up |
| Isoleucine | Amino acid derivative | Marker | Similar to DL-Valine and DL-Leucine. | Up |
| L-Dopa | Amino acid derivative | Deterioration | L-dopa can normalize filtration fraction and correct pre- and post-glomerular resistance by means of preferential post-glomerular vasodilatation as a precursor of dopamine synthesis. | Up |
| Alpha-Linolenic acid | Lipid derivative | Marker | Linolenic acid intake was negatively correlated with DKD in Brazilian T2D patients. | Up |
| 13-Hotre(R) | Lipid derivative | Alleviation | 13-Hotre(R) improves inflammation and oxidative stress, and low long-term 13-Hotre(R) intake is associated with the development of chronic kidney disease in T2D. | Down |
| Carnitine and its derivative | Lipid derivative | Marker and Alleviation | Carnitine, a derivative of methionine, increases circulatory levels in chronic renal failure, but inhibits tubular oxidative stress, interstitial fibrosis and apoptosis. | Up |
| Lysophosphatidylcholine and derivatives | Lipid derivative | Marker and Deterioration | As a pro-inflammatory signal, these substances can recruit macrophages to attack. The accumulation level and activation degree of receptors are positively correlated with UACR and glomerular hypertrophy. | Up |
| Isoproterenol | Lipid derivative | Alleviation/Deterioration | On the one hand, there is evidence that isoproterenol can promote vascular dilation and stimulate proximal tubule proliferation to protect kidney; on the other hand, there are studies supporting that isoproterenol can cause renal tubule injury by triggering oxidative stress and endoplasmic reticulum stress. | Up |
| 7-KCHO | Lipid derivative | Deterioration | It is a kind of cholesterol derivative, which can induce vascular cell apoptosis by promoting oxygen and inflammation. | Up |
| 2-deoxyglucose-6-phosphate | Carbohydrate derivative | Deterioration | It increases the activity of glucokinase causing hyperactivation of glucose metabolism. | Up |
| D−Arabinose 5−phosphate | Carbohydrate derivative | Marker | Circulation level of it increased in STZ modelled diabetic rats. | Up |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, C.; Fei, J.; Xu, Q.; Tao, Y.; Zhou, Z.; Wang, Y.; Wu, J.; Gu, H.F. Interaction between Plasma Metabolomics and Intestinal Microbiome in db/db Mouse, an Animal Model for Study of Type 2 Diabetes and Diabetic Kidney Disease. Metabolites 2022, 12, 775. https://doi.org/10.3390/metabo12090775
Wu C, Fei J, Xu Q, Tao Y, Zhou Z, Wang Y, Wu J, Gu HF. Interaction between Plasma Metabolomics and Intestinal Microbiome in db/db Mouse, an Animal Model for Study of Type 2 Diabetes and Diabetic Kidney Disease. Metabolites. 2022; 12(9):775. https://doi.org/10.3390/metabo12090775
Chicago/Turabian StyleWu, Chenhua, Jingjing Fei, Qing Xu, Yingjun Tao, Ziqi Zhou, Yurong Wang, Jie Wu, and Harvest F. Gu. 2022. "Interaction between Plasma Metabolomics and Intestinal Microbiome in db/db Mouse, an Animal Model for Study of Type 2 Diabetes and Diabetic Kidney Disease" Metabolites 12, no. 9: 775. https://doi.org/10.3390/metabo12090775
APA StyleWu, C., Fei, J., Xu, Q., Tao, Y., Zhou, Z., Wang, Y., Wu, J., & Gu, H. F. (2022). Interaction between Plasma Metabolomics and Intestinal Microbiome in db/db Mouse, an Animal Model for Study of Type 2 Diabetes and Diabetic Kidney Disease. Metabolites, 12(9), 775. https://doi.org/10.3390/metabo12090775