Insulin Secretory Actions of Ethanol Extract of Eucalyptus citriodora Leaf, including Plasma DPP-IV and GLP-1 Levels in High-Fat-Fed Rats, as Well as Characterization of Biologically Effective Phytoconstituents




Abstract
:1. Introduction
2. Materials and Methods
2.1. Collection and Preparation of Plant Extracts
2.2. In Vitro Insulin-Releasing Studies
2.3. Membrane Potential and Intracellular Calcium ([Ca2+]i)
2.4. Cellular Glucose Uptake
2.5. Glycation of Insulin
2.6. DPP-IV Enzyme Activity In Vitro
2.7. Starch Digestion
2.8. Glucose Diffusion In Vitro
2.9. Animals
2.10. Oral Glucose Tolerance Test
2.11. DPP-IV Enzyme Activity
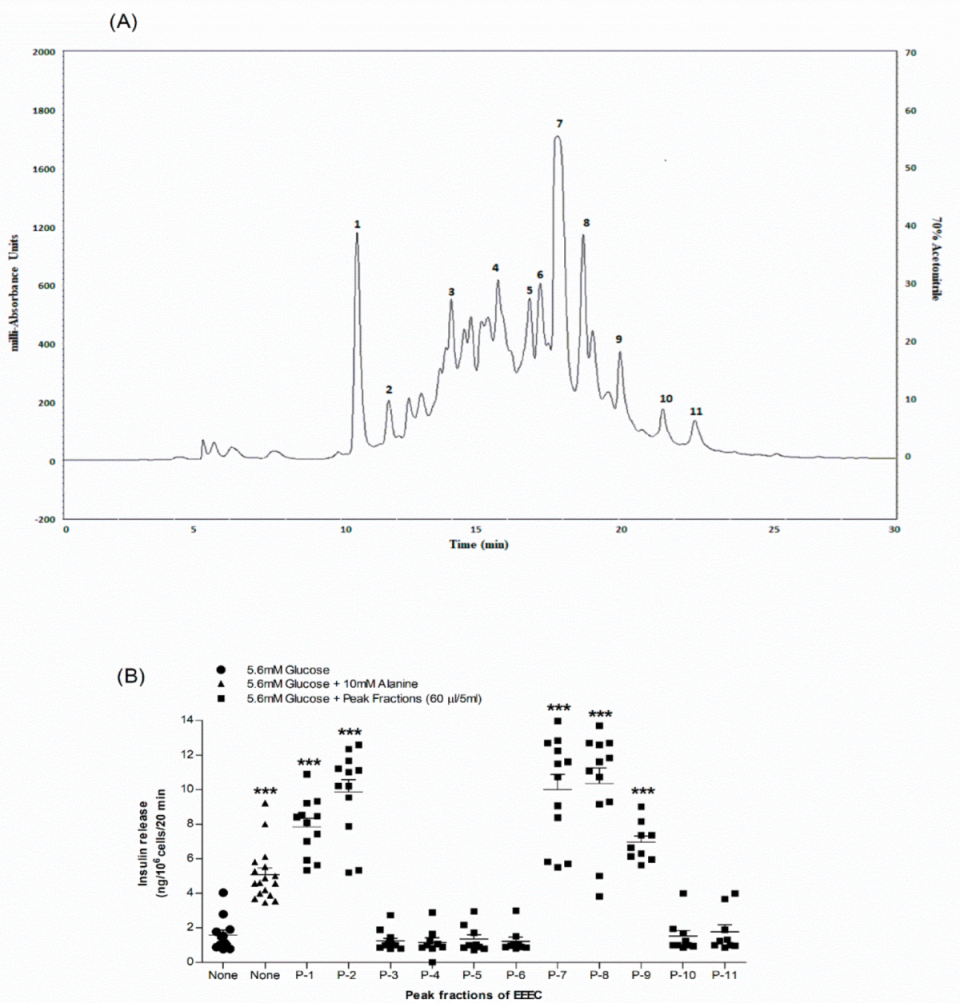
2.12. Purification of Crude Extract
2.13. Mass Spectroscopy
2.14. Statistical Analysis
3. Results
3.1. EEEC and Insulin Release
3.2. EEEC and Known Modulators of Insulin Release, Inhibitors and Free Ca2+ Conditions
3.3. EEEC and Membrane Depolarization and [Ca2+]i
3.4. EEEC and Glycation of Insulin
3.5. EEEC and Glucose Uptake and Insulin Action
3.6. EEEC and Starch Digestion
3.7. EEEC and Glucose Diffusion In Vitro
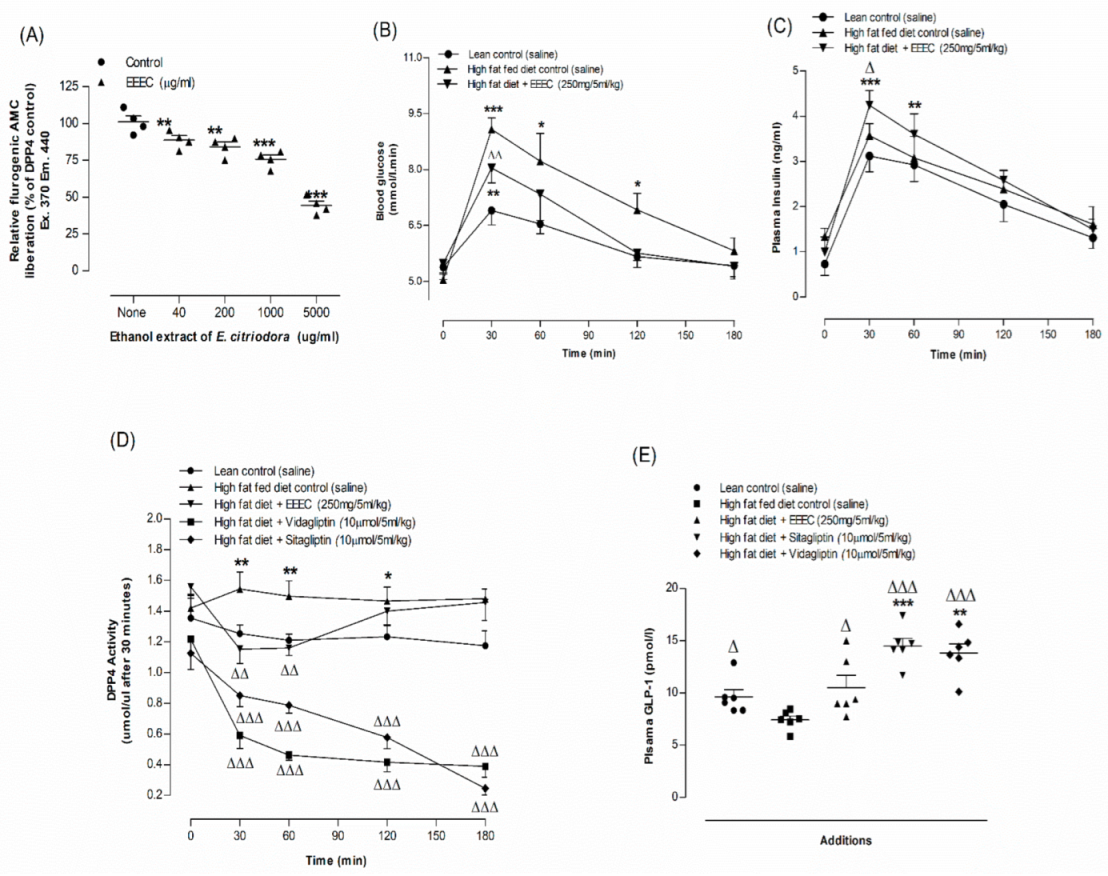
3.8. EEEC and DPP-IV Enzyme Activity In Vitro
3.9. EEEC and Oral Glucose Tolerance and Plasma Insulin, DPP-IV and Active GLP-1 (7-36) Levels
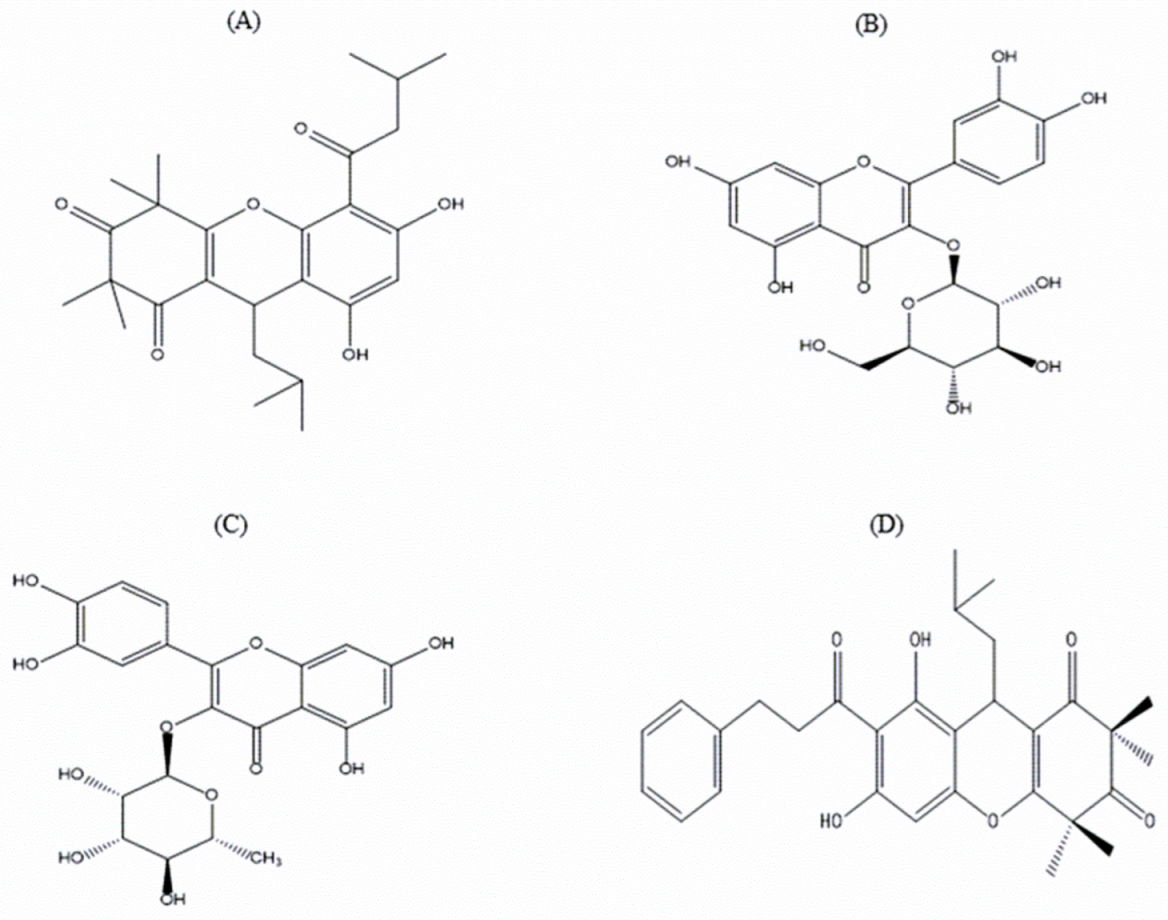
3.10. Identification of Purified Extract
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| AGE | Advanced glycation end-products |
| AMPK | AMP-activated protein kinase |
| BMI | Body mass index |
| CEDT | Cellulose ester dialysis tube |
| cAMP | Cyclic adenosine monophosphate. |
| COPD | Chronic obstructive pulmonary disease |
| CVD | Cardiovascular diseases |
| DM | Diabetes Mellitus |
| DPP-IV | Dipeptidyl peptidase IV |
| EEEC | Ethanol extract of Eucalyptus citriodora |
| GIP | Glucose-dependent insulinotropic polypeptide |
| GLP-1 | Glucagon-like peptide-1 |
| GLUT4 | Glucose transporter type 4 |
| HFF | High-fat-fed |
| IBMX | 3-isobutyl-1-methylxanthine |
| KCl | Potassium chloride |
| LC-MS | Liquid chromatography–mass spectrometry |
| MAPK | Mitogen-activated protein kinase |
| NEFA | Non-esterified fatty acids |
| NMR | Nuclear magnetic resonance |
| PI3K | Phosphoinositide 3-kinase (PI3K) |
| RP-HPLC | Reverse-phase high performance liquid chromatography |
| T2DM | Type 2 Diabetes Mellitus |
References
- Lipscombe, L.L.; Austin, P.C.; Manuel, D.G.; Shah, B.R.; Hux, J.E.; Booth, G.L. Income-related differences in mortality among people with diabetes mellitus. Can. Med. Assoc. J. 2010, 182, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Trivedi, N.A.; Mazumdar, B.; Bhatt, J.D.; Hemavathi, K.G. Effect of Shilajit on blood glucose and lipid profile in alloxan-induced diabetic rats. Indian J. Pharmacol. 2004, 36, 373. [Google Scholar]
- Sun, H.; Saeedi, P.; Karuranga, S.; Pinkepank, M.; Ogurtsova, K.; Duncan, B.B.; Stein, C.; Basit, A.; Chan, J.C.; Mbanya, J.C. IDF Diabetes Atlas: Global, regional and country-level diabetes prevalence estimates for 2021 and projections for 2045. Diabetes. Res. Clin. Pract. 2022, 183, 109119. [Google Scholar] [CrossRef] [PubMed]
- King, H.; Aubert, R.E.; Herman, W.H. Global burden of diabetes, 1995–2025: Prevalence, numerical estimates, and projections. Diabetes Care 1998, 21, 1414–1431. [Google Scholar] [CrossRef]
- Ansari, P.; Flatt, P.R.; Harriott, P.; Abdel-Wahab, Y.H.A. Anti-Hyperglycaemic and Insulin-Releasing Effects of Camellia Sinensis Leaves and Isolation and Characterisation of Active Compounds. Brit. J. Nutr. 2020, 126, 1149–1163. [Google Scholar] [CrossRef]
- Al-Goblan, A.S.; Al-Alfi, M.A.; Khan, M.Z. Mechanism linking diabetes mellitus and obesity. Diabetes Metab. Syndr. Obes. Targets Ther. 2014, 7, 587. [Google Scholar] [CrossRef]
- Piché, M.-E.; Tchernof, A.; Després, J.-P. Obesity Phenotypes, Diabetes, and cardiovascular diseases. Circ. Res. 2020, 126, 1477–1500. [Google Scholar] [CrossRef]
- Ansari, P.; Flatt, P.R.; Harriott, P.; Hannan, J.M.A.; Abdel-Wahab, Y.H.A. Identification of Multiple Pancreatic and Extra-Pancreatic Pathways Underlying the Glucose-Lowering Actions of Acacia Arabica Bark in Type-2 Diabetes and Isolation of Active Phytoconstituents. Plants 2021, 10, 1190. [Google Scholar] [CrossRef]
- Flatt, P.R. Dipeptidyl Peptidase IV (DPP IV) and Related Molecules in Type 2 Diabetes. Front. Biosci. 2008, 13, 3648. [Google Scholar] [CrossRef]
- Seino, Y.; Fukushima, M.; Yabe, D. GIP and GLP-1, the Two Incretin Hormones: Similarities and Differences. J. Diabetes Investig. 2010, 1, 8–23. [Google Scholar] [CrossRef]
- Inzucchi, S.E.; Bergenstal, R.M.; Buse, J.B.; Diamant, M.; Ferrannini, E.; Nauck, M.; Peters, A.L.; Tsapas, A.; Wender, R.; Matthews, D.R. Management of Hyperglycemia in Type 2 Diabetes, 2015: A Patient-Centered Approach: Update to a Position Statement of the American Diabetes Association and the European Association for the Study of Diabetes. Diabetes Care 2015, 38, 140–149. [Google Scholar] [CrossRef] [PubMed]
- Jung, M.; Park, M.; Lee, H.C.; Kang, Y.-H.; Kang, E.S.; Kim, S.K. Antidiabetic Agents from Medicinal Plants. Curr. Med. Chem. 2006, 13, 1203–1218. [Google Scholar] [CrossRef] [PubMed]
- Zulcafli, A.S.; Lim, C.; Ling, A.P.; Chye, S.; Koh, R. Antidiabetic Potential of Syzygium Sp.: An Overview. Yale J. Biol. Med. 2020, 93, 307–325. [Google Scholar] [PubMed]
- Unuofin, J.O.; Lebelo, S.L. Antioxidant Effects and Mechanisms of Medicinal Plants and Their Bioactive Compounds for the Prevention and Treatment of Type 2 Diabetes: An Updated Review. Oxid. Med. Cell. Longev. 2020, 2020, 1356893. [Google Scholar] [CrossRef] [PubMed]
- Ansari, P.; Hannon-Fletcher, M.P.; Flatt, P.R.; Abdel-Wahab, Y.H.A. Effects of 22 Traditional Anti-Diabetic Medicinal Plants on DPP-IV Enzyme Activity and Glucose Homeostasis in High-Fat Fed Obese Diabetic Rats. Biosci. Rep. 2021, 41, BSR20203824. [Google Scholar] [CrossRef]
- Ramezani, H.; Singh, H.P.; Batish, D.R.; Kohli, R.K. Antifungal Activity of the Volatile Oil of Eucalyptus Citriodora. Fitoterapia 2002, 73, 261–262. [Google Scholar] [CrossRef]
- Kesharwani, V.; Gupta, S.; Kushwaha, N.; Kesharwani, R.; Patel, D.K. A review on therapeutics application of Eucalyptus oil. Int. J. Herb. Med. 2018, 6, 110–115. [Google Scholar]
- Patra, A.; Jha, S.; Sahu, A.N. Antidiabetic activity of aqueous extract of Eucalyptus citriodora hook. in alloxan induced diabetic rats. Pharmacogn. Mag. 2009, 5, 51–54. [Google Scholar]
- Ho, C.-L.; Li, L.-H.; Weng, Y.-C.; Hua, K.-F.; Ju, T.-C. Eucalyptus Essential Oils Inhibit the Lipopolysaccharide-Induced Inflammatory Response in RAW264.7 Macrophages through Reducing MAPK and NF-ΚB Pathways. BMC Complement. Altern. Med. 2020, 20, 200. [Google Scholar] [CrossRef]
- Saba, I.; Iqbal, M.J.; Iqbal, M. Bioactivity of Eucalyptus citriodora leaves essential oil. J. Agrochimica. 2013, 57, 128. [Google Scholar]
- Gbenou, J.D.; Ahounou, J.F.; Akakpo, H.B.; Laleye, A.; Yayi, E.; Gbaguidi, F.; Baba-Moussa, L.; Darboux, R.; Dansou, P.; Moudachirou, M.; et al. Phytochemical Composition of Cymbopogon Citratus and Eucalyptus Citriodora Essential Oils and Their Anti-Inflammatory and Analgesic Properties on Wistar Rats. Mol. Biol. Rep. 2012, 40, 1127–1134. [Google Scholar] [CrossRef] [PubMed]
- Ansari, P.; Flatt, P.R.; Harriott, P.; Abdel-Wahab, Y.H. Insulinotropic and antidiabetic properties of Eucalyptus citriodora leaves and isolation of bioactive phytomolecules. J. Pharm. Pharmacol. 2021, 73, 1049–1061. [Google Scholar] [CrossRef] [PubMed]
- Ansari, P.; Azam, S.; Seidel, V.; Abdel-Wahab, Y.H.A. In vitro and in vivo antihyperglycemic activity of the ethanol extract of Heritiera fomes bark and characterization of pharmacologically active phytomolecules. J. Pharm. Pharmacol. 2022, 3, 415–425. [Google Scholar] [CrossRef] [PubMed]
- Hannan, J.M.A.; Ansari, P.; Haque, A.; Sanju, A.; Huzaifa, A.; Rahman, A.; Ghosh, A.; Azam, S. Nigella Sativa Stimulates Insulin Secretion from Isolated Rat Islets and Inhibits the Digestion and Absorption of (CH2O)n in the Gut. Biosci. Rep. 2019, 39, BSR20190723. [Google Scholar] [CrossRef]
- Ansari, P.; Azam, S.; Hannan, J.M.A.; Flatt, P.R.; Abdel Wahab, Y.H.A. Anti-hyperglycemic activity of H. rosa-sinensis leaves is partly mediated by inhibition of carbohydrate digestion and absorption, and enhancement of insulin secretion. J. Ethnopharmacol. 2020, 253, 112647. [Google Scholar] [CrossRef]
- Hannan, J.M.A.; Ansari, P.; Azam, S.; Flatt, P.R.; Abdel Wahab, Y.H.A. Effects of Spirulina Platensis on Insulin Secretion, Dipeptidyl Peptidase IV Activity and Both Carbohydrate Digestion and Absorption Indicate Potential as an Adjunctive Therapy for Diabetes. Brit. J. Nutr. 2020, 124, 1021–1034. [Google Scholar] [CrossRef]
- Flatt, P.R.; Bailey, C.J. Abnormal plasma glucose and insulin responses in heterozygous lean (ob/+) mice. Diabetologia 1981, 20, 573–577. [Google Scholar] [CrossRef]
- Abdel-Wahab, Y.H.A.; Marenah, L.; Flatt, P.R.; Conlon, J.M. Insulin releasing properties of the Temporin family of antimicrobial peptides. Protein Pept. Lett. 2007, 14, 702–707. [Google Scholar] [CrossRef]
- Hannan, J.M.A.; Ali, L.; Khaleque, J.; Akhter, M.; Flatt, P.R.; Abdel-Wahab, Y.H.A. Antihyperglycemic activity of Asparagus racemosus roots is partly mediated by inhibition of carbohydrate digestion and absorption, and enhancement of cellular insulin action. Brit. J. Nutri. 2012, 107, 1316–1323. [Google Scholar] [CrossRef]
- Duffy, N.A.; Green, B.D.; Irwin, N.; Gault, V.A.; McKillop, A.M.; O’Harte, F.P.M.; Flatt, P.R. Effects of Antidiabetic Drugs on Dipeptidyl Peptidase IV Activity: Nateglinide Is an Inhibitor of DPP IV and Augments the Antidiabetic Activity of Glucagon-like Peptide-1. Eur. J. Pharmacol. 2007, 568, 278–286. [Google Scholar] [CrossRef]
- Thomson, H.; Ojo, O.; Flatt, P.; AbdelWahab, Y. Antidiabetic actions of aqueous bark extract of Swertia chirayita on insulin secretion, cellular glucose uptake and protein glycation. J. Exp. Integrat. Med. 2014, 4, 268. [Google Scholar] [CrossRef]
- O’Harte, F.P.M.; Højrup, P.; Barnett, C.R.; Flatt, P.R. Identification of the Site of Glycation of Human Insulin. Peptides 1996, 17, 1323–1330. [Google Scholar] [CrossRef]
- Gallagher, A.M.; Flatt, P.R.; Duffy, G.; Abdel-Wahab, Y.H.A. The effects of traditional antidiabetic plants on in vitro glucose diffusion. Nutr. Res. 2003, 23, 413–424. [Google Scholar] [CrossRef]
- Ansari, P.; Flatt, P.R.; Harriott, P.; Abdel-Wahab, Y.H.A. Evaluation of the Antidiabetic and Insulin Releasing Effects of A. Squamosa, Including Isolation and Characterization of Active Phytochemicals. Plants 2020, 9, 1348. [Google Scholar] [CrossRef]
- Ansari, P.; Flatt, P.R.; Harriott, P.; Abdel-Wahab, Y.H.A. Insulin secretory and antidiabetic actions of Heritiera fomes bark together with isolation of active phytomolecules. PLoS ONE 2022, 17, e0264632. [Google Scholar] [CrossRef]
- Sola, D.; Rossi, L.; Schianca, G.P.C.; Maffioli, P.; Bigliocca, M.; Mella, R.; Corlianò, F.; Fra, G.P.; Bartoli, E.; Derosa, G. Sulfonylureas and Their Use in Clinical Practice. Arch. Med. Sci. 2015, 4, 840–848. [Google Scholar] [CrossRef]
- Park, J.E.; Han, J.S. A Portulaca oleracea L. extract promotes insulin secretion via a K+ATP channel dependent pathway in INS-1 pancreatic β-cells. Nutr. Res. Pract. 2018, 12, 183–190. [Google Scholar] [CrossRef]
- Dey, B.; Mitra, A. Chemo-Profiling of Eucalyptus and Study of Its Hypoglycemic Potential. World J. Diabetes 2013, 4, 170. [Google Scholar] [CrossRef]
- Billington, C.K.; Ojo, O.O.; Penn, R.B.; Ito, S. CAMP Regulation of Airway Smooth Muscle Function. Pulm. Pharmacol. Ther. 2013, 26, 112–120. [Google Scholar] [CrossRef]
- Lambadiari, V.; Triantafyllou, K.; Dimitriadis, G.D. Insulin Action in Muscle and Adipose Tissue in Type 2 Diabetes: The Significance of Blood Flow. World J. Diabetes 2015, 6, 626. [Google Scholar] [CrossRef]
- Xu, P.-T.; Song, Z.; Zhang, W.-C.; Jiao, B.; Yu, Z.-B. Impaired Translocation of GLUT4 Results in Insulin Resistance of Atrophic Soleus Muscle. BioMed Res. Int. 2015, 2015, 291987. [Google Scholar] [CrossRef] [PubMed]
- Fang, X.-K.; Gao, J.; Zhu, D.-N. Kaempferol and Quercetin Isolated from Euonymus alatus Improve Glucose Uptake of 3T3-L1 Cells without Adipogenesis Activity. Life Sci. 2008, 82, 615–622. [Google Scholar] [CrossRef] [PubMed]
- Telapolu, S.; Kalachavedu, M.; Punnoose, A.M.; Bilikere, D. MD-1, a Poly Herbal Formulation Indicated in Diabetes Mellitus Ameliorates Glucose Uptake and Inhibits Adipogenesis–an in Vitro Study. BMC Complement. Altern. Med. 2018, 18, 113. [Google Scholar] [CrossRef]
- Russo, B.; Picconi, F.; Malandrucco, I.; Frontoni, S. Flavonoids and Insulin-Resistance: From Molecular Evidences to Clinical Trials. Int. J. Mol. Sci. 2019, 20, 2061. [Google Scholar] [CrossRef] [PubMed]
- Amakura, Y.; Yoshimura, M.; Sugimoto, N.; Yamazaki, T.; Yoshida, T. Marker Constituents of the Natural Antioxidant Eucalyptus Leaf Extract for the Evaluation of Food Additives. Biosci. Biotechnol. Biochem. 2009, 73, 1060–1065. [Google Scholar] [CrossRef]
- Singh, V.P.; Bali, A.; Singh, N.; Jaggi, A.S. Advanced Glycation End Products and Diabetic Complications. Korean J. Physiol. Pharmacol. 2014, 18, 1025926. [Google Scholar] [CrossRef]
- Singh, H.P.; Kaur, S.; Negi, K.; Kumari, S.; Saini, V.; Batish, D.R.; Kohli, R.K. Assessment of in Vitro Antioxidant Activity of Essential Oil of Eucalyptus Citriodora (Lemon-Scented Eucalypt; Myrtaceae) and Its Major Constituents. LWT–Food Sci. Technol. 2012, 48, 237–241. [Google Scholar] [CrossRef]
- Miguel, M.; Gago, C.; Antunes, M.; Lagoas, S.; Faleiro, M.; Megías, C.; Cortés-Giraldo, I.; Vioque, J.; Figueiredo, A. Antibacterial, Antioxidant, and Antiproliferative Activities of Corymbia citriodora and the Essential Oils of Eight Eucalyptus Species. Medicines 2018, 5, 61. [Google Scholar] [CrossRef]
- Al-Sayed, E.; El-Naga, R.N. Protective Role of Ellagitannins from Eucalyptus Citriodora against Ethanol-Induced Gastric Ulcer in Rats: Impact on Oxidative Stress, Inflammation and Calcitonin-Gene Related Peptide. Phytomedicine 2015, 22, 5–15. [Google Scholar] [CrossRef]
- Ceriello, A.; Davidson, J.; Hanefeld, M.; Leiter, L.; Monnier, L.; Owens, D.; Tajima, N.; Tuomilehto, J. Postprandial Hyperglycaemia and Cardiovascular Complications of Diabetes: An Update. Nutr. Metab. Cardiovasc. Dis. 2006, 16, 453–456. [Google Scholar] [CrossRef]
- Dey, B.; Mitra, A.; Katakam, P.; Singla, R.K. Exploration of Natural Enzyme Inhibitors with Hypoglycemic Potentials amongst Eucalyptus Spp. By in Vitro Assays. World J. Diabetes 2014, 5, 209. [Google Scholar] [CrossRef] [PubMed]
- Kidane, Y.; Bokrezion, T.; Mebrahtu, J.; Mehari, M.; Gebreab, Y.B.; Fessehaye, N.; Achila, O.O. In Vitro Inhibition of α-Amylase and α-Glucosidase by Extracts from Psiadia Punctulata and Meriandra Bengalensis. Evid. Based Complement. Altern. Med. 2018, 2018, 2164345. [Google Scholar] [CrossRef] [PubMed]
- Ansari, P.; Akther, S.; Hannan, J.M.A.; Seidel, V.; Nujat, N.J.; Abdel-Wahab, Y.H.A. Pharmacologically Active Phytomolecules Isolated from Traditional Antidiabetic Plants and Their Therapeutic Role for the Management of Diabetes Mellitus. Molecules 2022, 27, 4278. [Google Scholar] [CrossRef] [PubMed]
- Lattimer, J.M.; Haub, M.D. Effects of Dietary Fiber and Its Components on Metabolic Health. Nutrients 2010, 2, 1266–1289. [Google Scholar] [CrossRef] [PubMed]
- Brownlee, M. The Pathobiology of Diabetic Complications: A Unifying Mechanism. Diabetes 2005, 54, 1615–1625. [Google Scholar] [CrossRef]
- Vella, A. Mechanism of Action of DPP-4 Inhibitors—New Insights. J. Clin. Endocrinol. Metab. 2012, 97, 2626–2628. [Google Scholar] [CrossRef]
- Deacon, C.F. Physiology and Pharmacology of DPP-4 in Glucose Homeostasis and the Treatment of Type 2 Diabetes. Front. Endocrinol. 2019, 10, 80. [Google Scholar] [CrossRef]
- Kim, B.-R.; Kim, H.; Choi, I.; Kim, J.-B.; Jin, C.; Han, A.-R. DPP-IV Inhibitory Potentials of Flavonol Glycosides Isolated from the Seeds of Lens Culinaris: In Vitro and Molecular Docking Analyses. Molecules 2018, 23, 1998. [Google Scholar] [CrossRef]
- Mohanty, I.R.; Borde, M.; Kumar, C.S.; Maheshwari, U. Dipeptidyl Peptidase IV Inhibitory Activity of Terminalia Arjuna Attributes to Its Cardioprotective Effects in Experimental Diabetes: In Silico, in Vitro and in Vivo Analyses. Phytomedicine 2019, 57, 158–165. [Google Scholar] [CrossRef]
- Hiranrat, A.; Mahabusarakam, W. New Acylphloroglucinols from the Leaves of Rhodomyrtus tomentosa. Tetrahedron 2008, 64, 11193–11197. [Google Scholar] [CrossRef]
- Gervais, A.; Lazarski, K.E.; Porco, J.A. Divergent Total Syntheses of Rhodomyrtosones A and B. J. Org. Chem. 2015, 80, 9584–9591. [Google Scholar] [CrossRef] [PubMed]
- Islam, M.; Al-Amin, M.; Siddiqi, M.M.A.; Akter, S.; Haque, M.M.; Sultana, N.; Chowdhury, A.S. Isolation of Quercetin-3-O-Beta-d-Glucopyranoside from the Leaves of Azadirachta indica and Antimicrobial and Cytotoxic Screening of the Crude Extracts. Dhaka Univ. J. Sci. 2012, 60, 11–14. [Google Scholar] [CrossRef]
- He, C.-Y.; Fu, J.; Ma, J.-Y.; Feng, R.; Tan, X.-S.; Huang, M.; Shou, J.-W.; Zhao, Z.-X.; Li, X.-Y.; Zhang, X.-F.; et al. Biotransformation and in Vitro Metabolic Profile of Bioactive Extracts from a Traditional Miao-Nationality Herbal Medicine, Polygonum capitatum. Molecules 2014, 19, 10291–10308. [Google Scholar] [CrossRef] [PubMed]
- Martos, I.; Ferreres, F.; Yao, L.; D’Arcy, B.; Caffin, N.; Tomás-Barberán, F.A. Flavonoids in Monospecific Eucalyptus Honeys from Australia. J. Agric. Food Chem. 2000, 48, 4744–4748. [Google Scholar] [CrossRef] [PubMed]
- Kharwar, R.N.; Gond, S.K.; Kumar, A.; Mishra, A. A Comparative Study of Endophytic and Epiphytic Fungal Association with Leaf of Eucalyptus citriodora Hook., and Their Antimicrobial Activity. World J. Microbiol. Biotechnol. 2010, 26, 1941–1948. [Google Scholar] [CrossRef]
- Vo, T.; Ngo, D. The Health Beneficial Properties of Rhodomyrtus tomentosa as Potential Functional Food. Biomolecules 2019, 9, 76. [Google Scholar] [CrossRef]
- Huang, X.-L.; He, Y.; Ji, L.-L.; Wang, K.-Y.; Wang, Y.-L.; Chen, D.-F.; Geng, Y.; OuYang, P.; Lai, W.-M. Hepatoprotective Potential of Isoquercitrin against Type 2 Diabetes-Induced Hepatic Injury in Rats. Oncotarget 2017, 8, 101545–101559. [Google Scholar] [CrossRef]
- Babujanarthanam, R.; Kavitha, P.; Pandian, M.R. Quercitrin, a Bioflavonoid Improves Glucose Homeostasis in Streptozotocin-Induced Diabetic Tissues by Altering Glycolytic and Gluconeogenic Enzymes. Fundam. Clin. Pharmacol. 2009, 24, 357–364. [Google Scholar] [CrossRef]
- Wang, C.; Yang, J.; Zhao, P.; Zhou, Q.; Mei, Z.; Yang, G.; Yang, X.; Feng, Y. Chemical Constituents from Eucalyptus citriodora Hook Leaves and Their Glucose Transporter 4 Translocation Activities. Bioorg. Med. Chem. Lett. 2014, 24, 3096–3099. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peak Samples | Retention Time (min) | Theoretical Molecular Weight (Da) | Found Molecular Weight (Da) | Predicted Compounds |
|---|---|---|---|---|
| P1 | 10.5 | 442.5 | 442.1 | Rhodomyrtosone B |
| P2 | 12 | - | 476.7 | Unknown |
| P7 | 18 | 464.4 | 464.2 | Quercetin-3-O-β-D-glucopyranoside |
| P8 | 19 | - | 490.9 | Rhodomyrtosone E |
| P9 | 20.5 | 448.4 | 447.9 | Quercitroside |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ansari, P.; Choudhury, S.T.; Abdel-Wahab, Y.H.A. Insulin Secretory Actions of Ethanol Extract of Eucalyptus citriodora Leaf, including Plasma DPP-IV and GLP-1 Levels in High-Fat-Fed Rats, as Well as Characterization of Biologically Effective Phytoconstituents. Metabolites 2022, 12, 757. https://doi.org/10.3390/metabo12080757
Ansari P, Choudhury ST, Abdel-Wahab YHA. Insulin Secretory Actions of Ethanol Extract of Eucalyptus citriodora Leaf, including Plasma DPP-IV and GLP-1 Levels in High-Fat-Fed Rats, as Well as Characterization of Biologically Effective Phytoconstituents. Metabolites. 2022; 12(8):757. https://doi.org/10.3390/metabo12080757
Chicago/Turabian StyleAnsari, Prawej, Samara T. Choudhury, and Yasser H. A. Abdel-Wahab. 2022. "Insulin Secretory Actions of Ethanol Extract of Eucalyptus citriodora Leaf, including Plasma DPP-IV and GLP-1 Levels in High-Fat-Fed Rats, as Well as Characterization of Biologically Effective Phytoconstituents" Metabolites 12, no. 8: 757. https://doi.org/10.3390/metabo12080757
APA StyleAnsari, P., Choudhury, S. T., & Abdel-Wahab, Y. H. A. (2022). Insulin Secretory Actions of Ethanol Extract of Eucalyptus citriodora Leaf, including Plasma DPP-IV and GLP-1 Levels in High-Fat-Fed Rats, as Well as Characterization of Biologically Effective Phytoconstituents. Metabolites, 12(8), 757. https://doi.org/10.3390/metabo12080757