
Direct In Vivo Analysis of CBD- and THC-Acid Cannabinoids and Classification of Cannabis Cultivars Using SpiderMass

 , ,
, , 
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Result
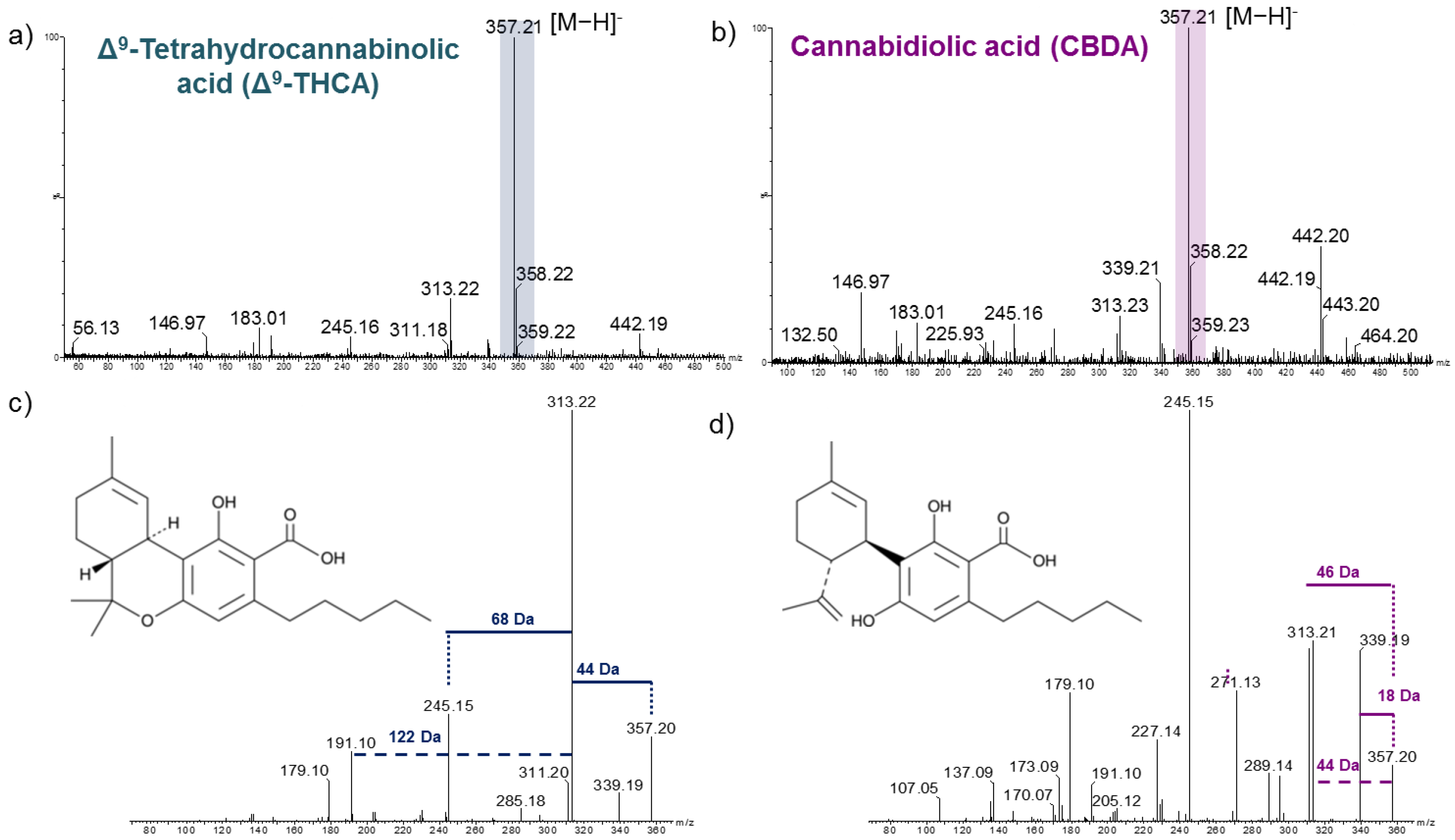
2.1. Analysis of Cannabinoid Standards
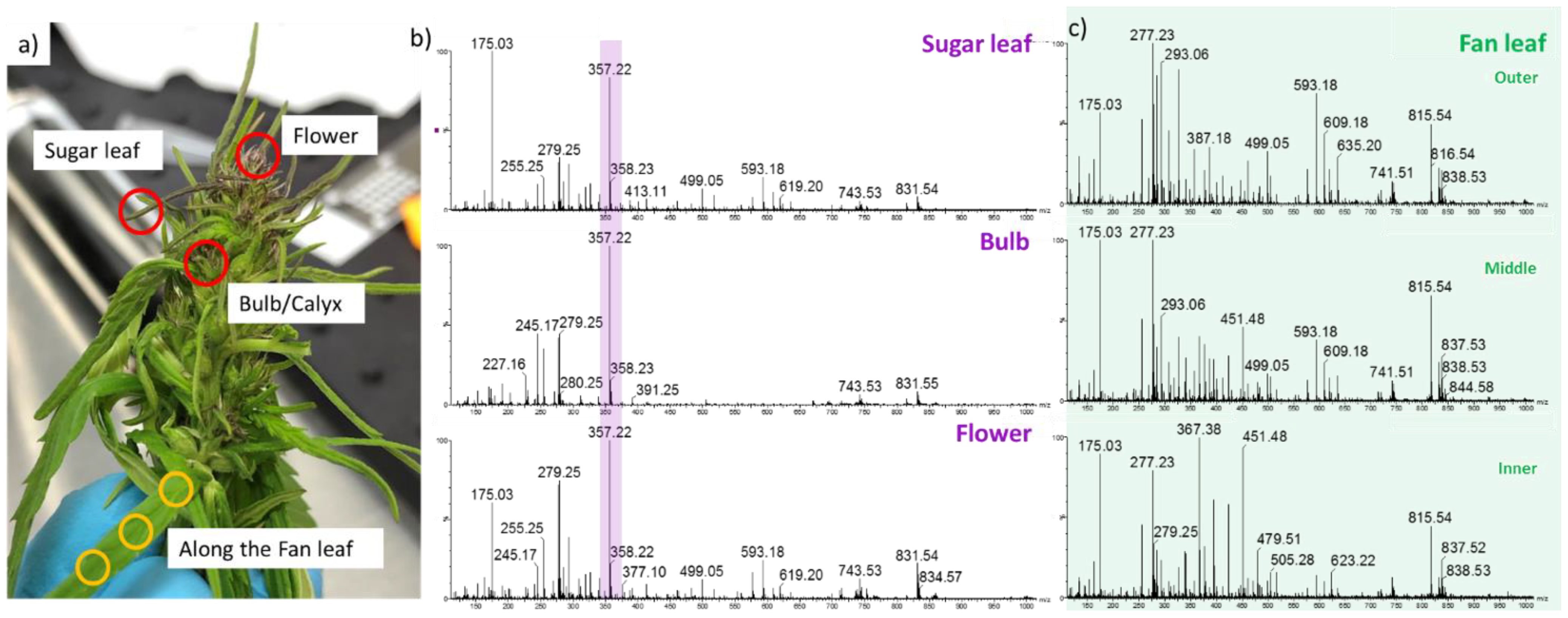
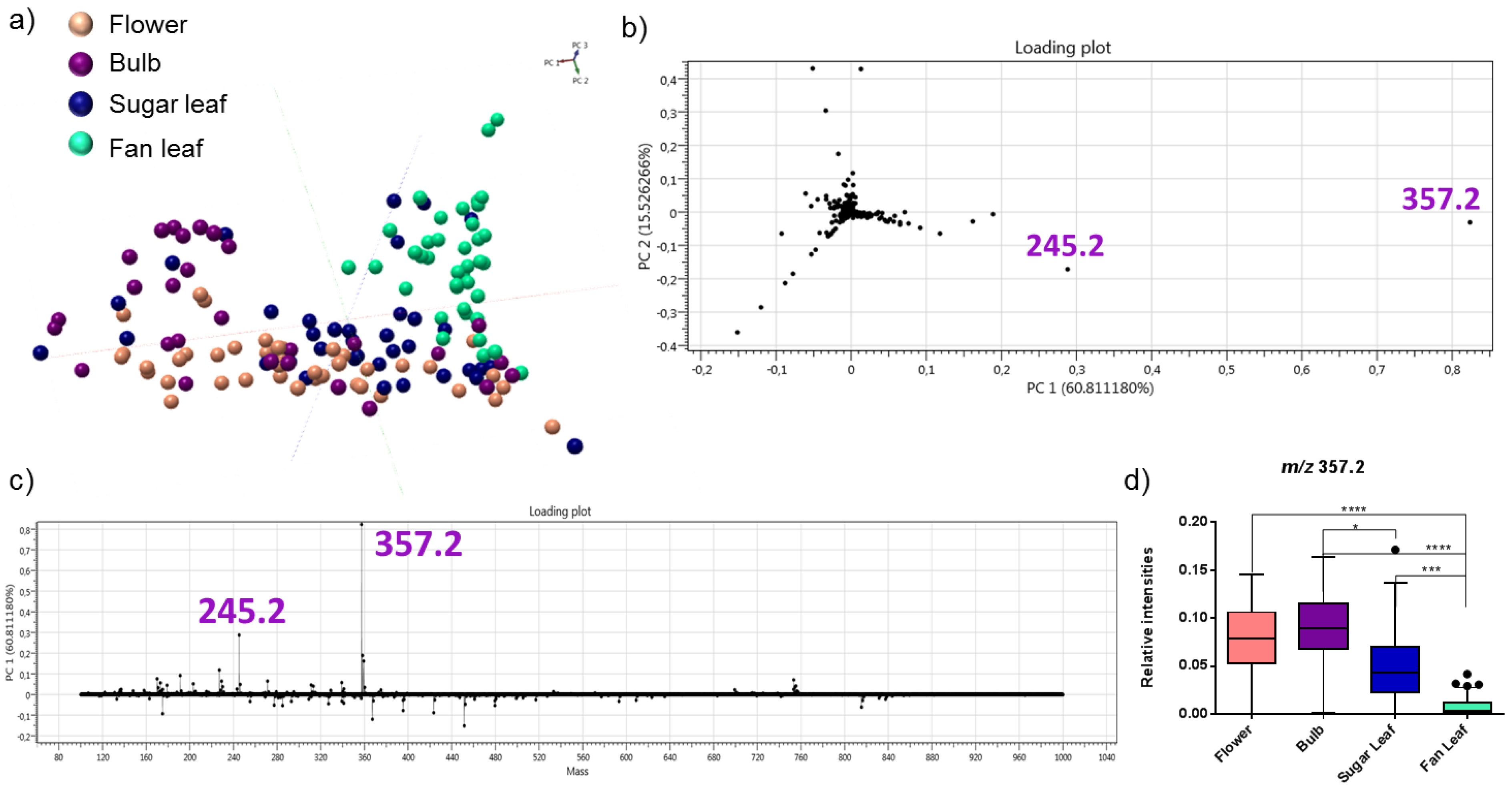
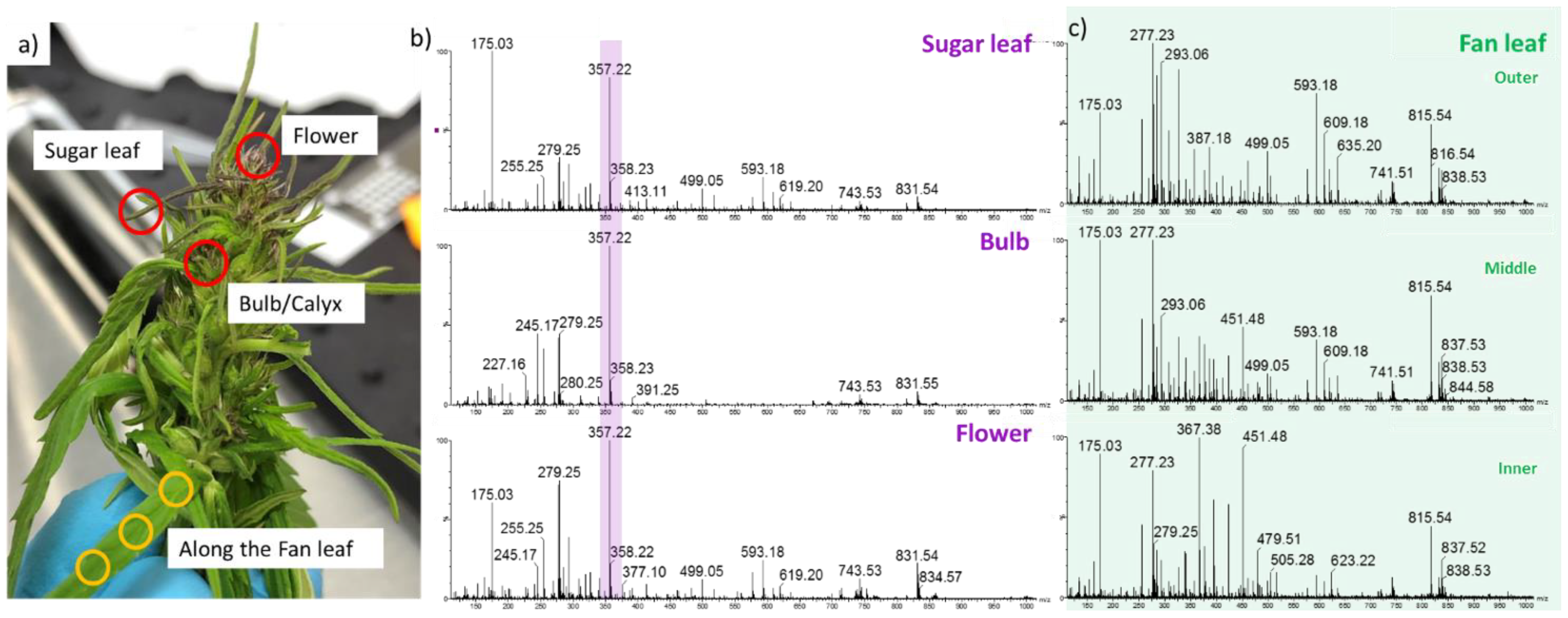
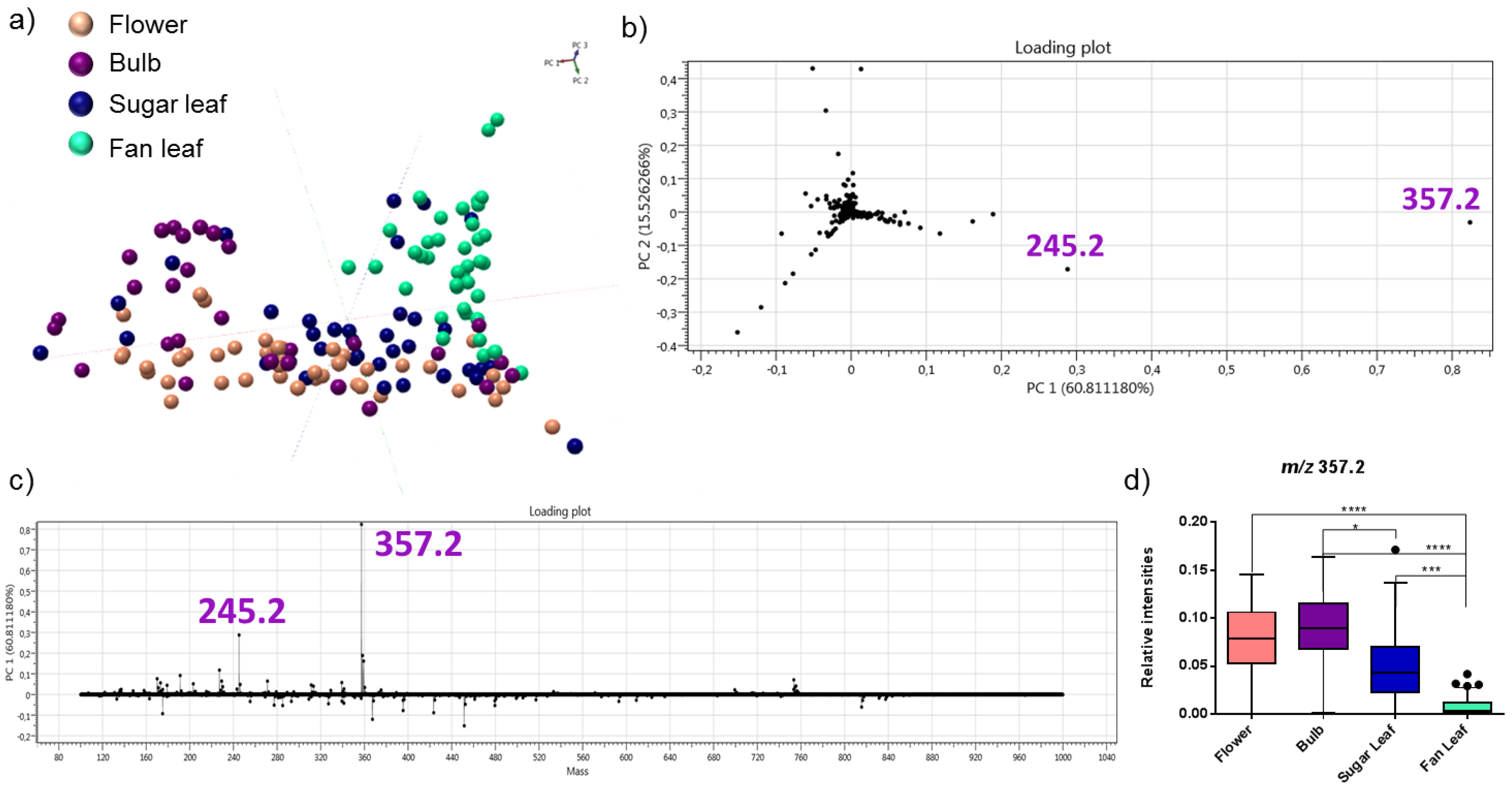
2.2. Detection of Acid Cannabinoids across the Plants
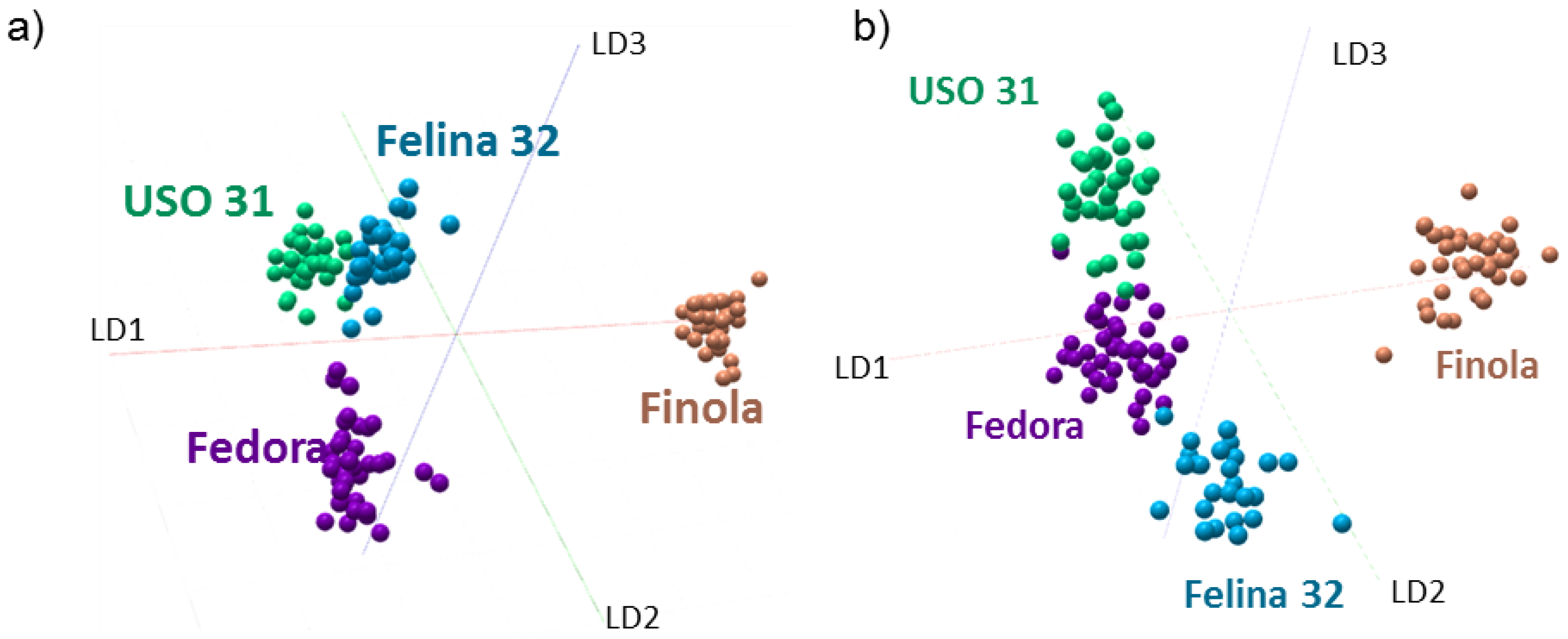
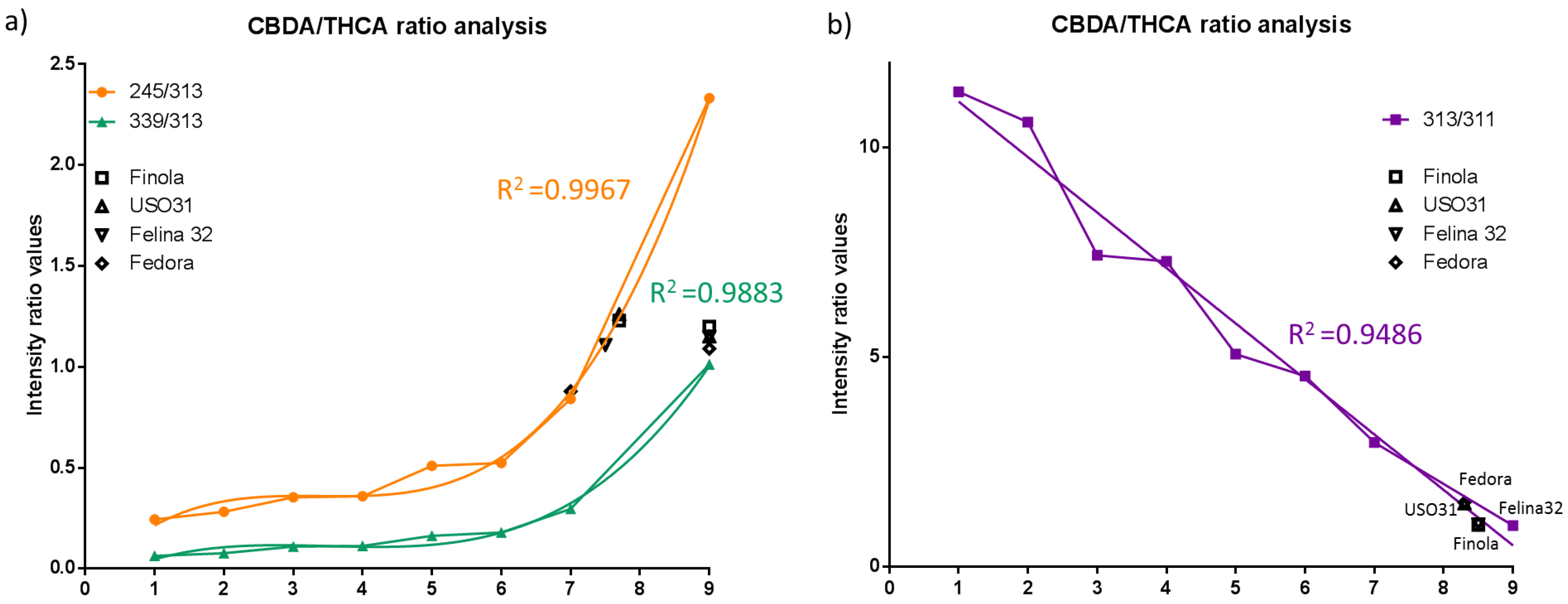
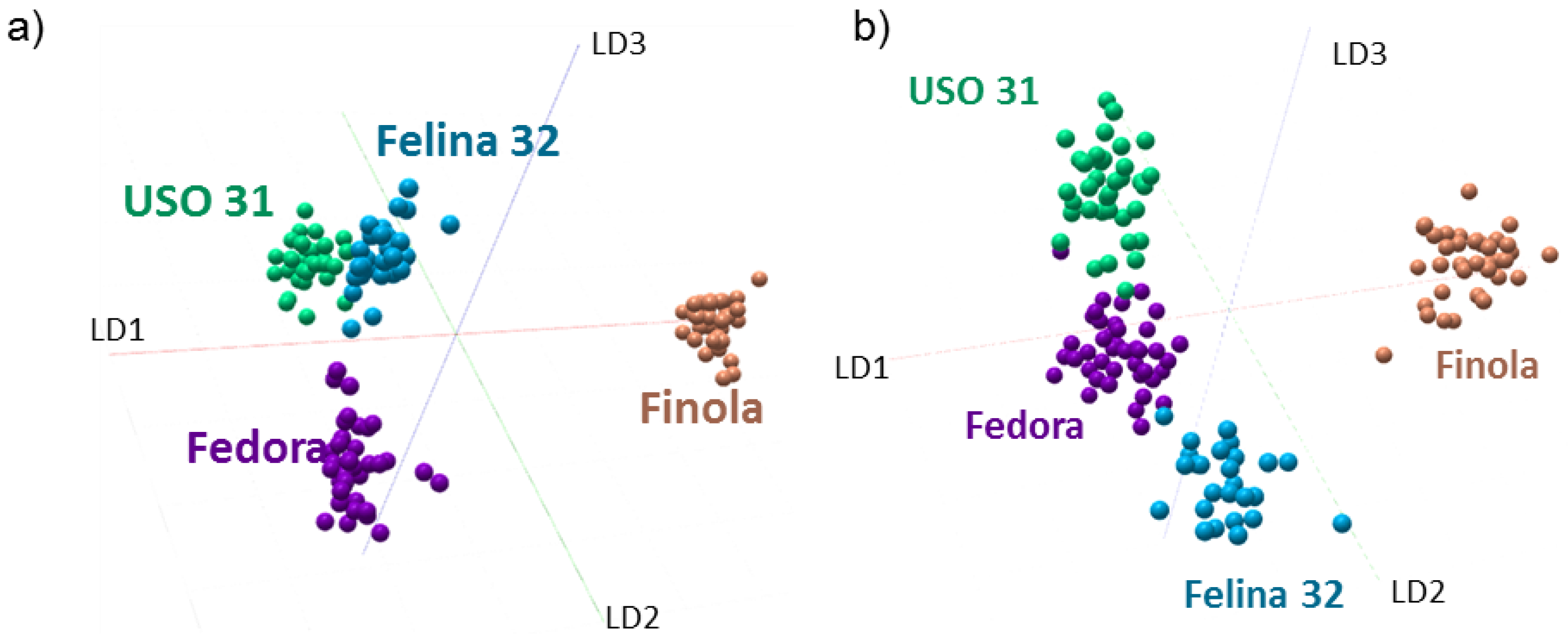
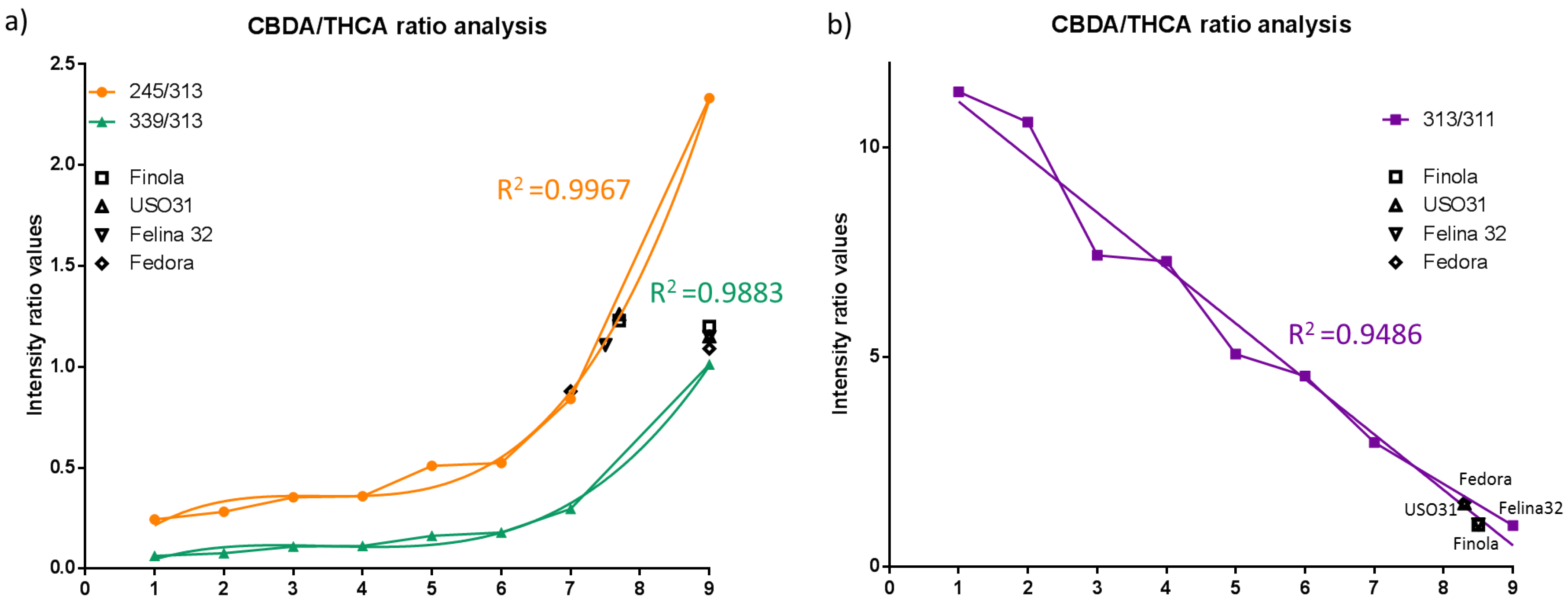
2.3. Discrimination of Cannabis Cultivars
3. Discussion
4. Materials and Methods
4.1. Cannabinoid Standards
4.2. Cultivation of Cannabis
4.3. SpiderMass and Data Analysis
4.4. HPLC-UV Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tahir, M.N.; Shahbazi, F.; Rondeau-Gagné, S.; Trant, J.F. The Biosynthesis of the Cannabinoids. J. Cannabis Res. 2021, 3, 7. [Google Scholar] [CrossRef] [PubMed]
- Nie, B.; Henion, J.; Ryona, I. The Role of Mass Spectrometry in the Cannabis Industry. J. Am. Soc. Mass. Spectrom. 2019, 30, 719–730. [Google Scholar] [CrossRef] [PubMed]
- Bonini, S.A.; Premoli, M.; Tambaro, S.; Kumar, A.; Maccarinelli, G.; Memo, M.; Mastinu, A. Cannabis Sativa: A Comprehensive Ethnopharmacological Review of a Medicinal Plant with a Long History. J. Ethnopharmacol. 2018, 227, 300–315. [Google Scholar] [CrossRef] [PubMed]
- Zekič, J.; Križman, M. Development of Gas-Chromatographic Method for Simultaneous Determination of Cannabinoids and Terpenes in Hemp. Molecules 2020, 25, 5872. [Google Scholar] [CrossRef] [PubMed]
- Capriotti, A.L.; Cannazza, G.; Catani, M.; Cavaliere, C.; Cavazzini, A.; Cerrato, A.; Citti, C.; Felletti, S.; Montone, C.M.; Piovesana, S.; et al. Recent Applications of Mass Spectrometry for the Characterization of Cannabis and Hemp Phytocannabinoids: From Targeted to Untargeted Analysis. J. Chromatogr. A 2021, 1655, 462492. [Google Scholar] [CrossRef]
- Commission Delegated Regulation (EU) No 639/2014 of 11 March 2014 Supplementing Regulation (EU) No 1307/2013 of the European Parliament and of the Council Establishing Rules for Direct Payments to Farmers under Support Schemes within the Framework of the Common Agricultural Policy and Amending Annex X to That Regulation. Available online: https://www.legislation.gov.uk/eur/2014/639/annex/iii (accessed on 18 March 2022).
- Brown, A.K.; Xia, Z.; Bulloch, P.; Idowu, I.; Francisco, O.; Stetefeld, J.; Stout, J.; Zimmer, J.; Marvin, C.; Letcher, R.J.; et al. Validated Quantitative Cannabis Profiling for Canadian Regulatory Compliance—Cannabinoids, Aflatoxins, and Terpenes. Anal. Chim. Acta 2019, 1088, 79–88. [Google Scholar] [CrossRef]
- Di Marco Pisciottano, I.; Guadagnuolo, G.; Soprano, V.; Esposito, M.; Gallo, P. A Survey of Δ9-THC and Relevant Cannabinoids in Products from the Italian Market: A Study by LC–MS/MS of Food, Beverages and Feed. Food Chem. 2021, 346, 128898. [Google Scholar] [CrossRef]
- Christinat, N.; Savoy, M.-C.; Mottier, P. Development, Validation and Application of a LC-MS/MS Method for Quantification of 15 Cannabinoids in Food. Food Chem. 2020, 318, 126469. [Google Scholar] [CrossRef]
- Scheunemann, A.; Elsner, K.; Germerott, T.; Hess, C.; Zörntlein, S.; Röhrich, J. Extensive Phytocannabinoid Profiles of Seized Cannabis and Cannabis-Based Medicines—Identification of Potential Distinguishing Markers. Forensic Sci. Int. 2021, 322, 110773. [Google Scholar] [CrossRef]
- Meng, Q.; Buchanan, B.; Zuccolo, J.; Poulin, M.-M.; Gabriele, J.; Baranowski, D.C. A Reliable and Validated LC-MS/MS Method for the Simultaneous Quantification of 4 Cannabinoids in 40 Consumer Products. PLoS ONE 2018, 13, e0196396. [Google Scholar] [CrossRef]
- Meija, J.; McRae, G.; Miles, C.O.; Melanson, J.E. Thermal Stability of Cannabinoids in Dried Cannabis: A Kinetic Study. Anal. Bioanal. Chem. 2022, 414, 377–384. [Google Scholar] [CrossRef]
- Raharjo, T.J.; Verpoorte, R. Methods for the Analysis of Cannabinoids in Biological Materials: A Review. Phytochem. Anal. 2004, 15, 79–94. [Google Scholar] [CrossRef]
- Kauppila, T.J.; Flink, A.; Laakkonen, U.-M.; Aalberg, L.; Ketola, R.A. Direct Analysis of Cannabis Samples by Desorption Atmospheric Pressure Photoionization-Mass Spectrometry. Drug Test. Anal. 2013, 5, 186–190. [Google Scholar] [CrossRef]
- Huang, S.; Claassen, F.W.; van Beek, T.A.; Chen, B.; Zeng, J.; Zuilhof, H.; Salentijn, G.I.J. Rapid Distinction and Semiquantitative Analysis of THC and CBD by Silver-Impregnated Paper Spray Mass Spectrometry. Anal. Chem. 2021, 93, 3794–3802. [Google Scholar] [CrossRef]
- Rodriguez-Cruz, S.E. Rapid Analysis of Controlled Substances Using Desorption Electrospray Ionization Mass Spectrometry. Rapid Commun. Mass Spectrom. 2006, 20, 53–60. [Google Scholar] [CrossRef]
- Piccolella, S.; Formato, M.; Pecoraro, M.T.; Crescente, G.; Pacifico, S. Discrimination of CBD-, THC- and CBC-Type Acid Cannabinoids through Diagnostic Ions by UHPLC-HR-MS/MS in Negative Ion Mode. J. Pharm. Biomed. Anal. 2021, 201, 114125. [Google Scholar] [CrossRef]
- Fatou, B.; Saudemont, P.; Leblanc, E.; Vinatier, D.; Mesdag, V.; Wisztorski, M.; Focsa, C.; Salzet, M.; Ziskind, M.; Fournier, I. In Vivo Real-Time Mass Spectrometry for Guided Surgery Application. Sci. Rep. 2016, 6, 25919. [Google Scholar] [CrossRef]
- Ogrinc, N.; Saudemont, P.; Balog, J.; Robin, Y.-M.; Gimeno, J.-P.; Pascal, Q.; Tierny, D.; Takats, Z.; Salzet, M.; Fournier, I. Water-Assisted Laser Desorption/Ionization Mass Spectrometry for Minimally Invasive in Vivo and Real-Time Surface Analysis Using SpiderMass. Nat. Protoc. 2019, 14, 3162–3182. [Google Scholar] [CrossRef]
- Gülck, T.; Møller, B.L. Phytocannabinoids: Origins and Biosynthesis. Trends Plant Sci. 2020, 25, 985–1004. [Google Scholar] [CrossRef]
- Piccolella, S.; Crescente, G.; Formato, M.; Pacifico, S. A Cup of Hemp Coffee by Moka Pot from Southern Italy: An UHPLC-HRMS Investigation. Foods 2020, 9, 1123. [Google Scholar] [CrossRef]
- Rubino, F.M. Center-of-Mass Iso-Energetic Collision-Induced Decomposition in Tandem Triple Quadrupole Mass Spectrometry. Molecules 2020, 25, 2250. [Google Scholar] [CrossRef]
- Konopko Cooperative. Co-op for Sustainable Production & Processing of Hemp- Welcome. Available online: https://www.konopko.si/en (accessed on 18 March 2022).
- Glivar, T.; Eržen, J.; Kreft, S.; Zagožen, M.; Čerenak, A.; Čeh, B.; Tavčar Benković, E. Cannabinoid Content in Industrial Hemp (Cannabis Sativa L.) Varieties Grown in Slovenia. Ind. Crops Prod. 2020, 145, 112082. [Google Scholar] [CrossRef]
- Russo, E.B. Taming THC: Potential Cannabis Synergy and Phytocannabinoid-Terpenoid Entourage Effects. Br. J. Pharmacol. 2011, 163, 1344–1364. [Google Scholar] [CrossRef]
- Nissen, L.; Zatta, A.; Stefanini, I.; Grandi, S.; Sgorbati, B.; Biavati, B.; Monti, A. Characterization and Antimicrobial Activity of Essential Oils of Industrial Hemp Varieties (Cannabis Sativa L.). Fitoterapia 2010, 81, 413–419. [Google Scholar] [CrossRef]
- Hädener, M.; Kamrath, M.Z.; Weinmann, W.; Groessl, M. High-Resolution Ion Mobility Spectrometry for Rapid Cannabis Potency Testing. Anal. Chem. 2018, 90, 8764–8768. [Google Scholar] [CrossRef]
- Dos Santos, N.A.; Tose, L.V.; da Silva, S.R.C.; Murgu, M.; Kuster, R.M.; Ortiz, R.S.; Camargo, F.A.O.; Vaz, B.G.; Lacerda, V.; Romão, W. Analysis of Isomeric Cannabinoid Standards and Cannabis Products by UPLC-ESI-TWIM-MS: A Comparison with GC-MS and GC × GC-QMS. J. Braz. Chem. Soc. 2019, 30, 60–70. [Google Scholar] [CrossRef]
- Balog, J.; Sasi-Szabó, L.; Kinross, J.; Lewis, M.R.; Muirhead, L.J.; Veselkov, K.; Mirnezami, R.; Dezső, B.; Damjanovich, L.; Darzi, A.; et al. Intraoperative Tissue Identification Using Rapid Evaporative Ionization Mass Spectrometry. Sci. Transl. Med. 2013, 5, 194ra93. [Google Scholar] [CrossRef]
- Smith, C.A.; O’Maille, G.; Want, E.J.; Qin, C.; Trauger, S.A.; Brandon, T.R.; Custodio, D.E.; Abagyan, R.; Siuzdak, G. METLIN: A Metabolite Mass Spectral Database. Ther. Drug Monit. 2005, 27, 747–751. [Google Scholar] [CrossRef]
- Pauling, J.K.; Hermansson, M.; Hartler, J.; Christiansen, K.; Gallego, S.F.; Peng, B.; Ahrends, R.; Ejsing, C.S. Proposal for a Common Nomenclature for Fragment Ions in Mass Spectra of Lipids. PLoS ONE 2017, 12, e0188394. [Google Scholar] [CrossRef] [Green Version]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ogrinc, N.; Schneider, S.; Bourmaud, A.; Gengler, N.; Salzet, M.; Fournier, I. Direct In Vivo Analysis of CBD- and THC-Acid Cannabinoids and Classification of Cannabis Cultivars Using SpiderMass. Metabolites 2022, 12, 480. https://doi.org/10.3390/metabo12060480
Ogrinc N, Schneider S, Bourmaud A, Gengler N, Salzet M, Fournier I. Direct In Vivo Analysis of CBD- and THC-Acid Cannabinoids and Classification of Cannabis Cultivars Using SpiderMass. Metabolites. 2022; 12(6):480. https://doi.org/10.3390/metabo12060480
Chicago/Turabian StyleOgrinc, Nina, Serge Schneider, Adèle Bourmaud, Nicolas Gengler, Michel Salzet, and Isabelle Fournier. 2022. "Direct In Vivo Analysis of CBD- and THC-Acid Cannabinoids and Classification of Cannabis Cultivars Using SpiderMass" Metabolites 12, no. 6: 480. https://doi.org/10.3390/metabo12060480
APA StyleOgrinc, N., Schneider, S., Bourmaud, A., Gengler, N., Salzet, M., & Fournier, I. (2022). Direct In Vivo Analysis of CBD- and THC-Acid Cannabinoids and Classification of Cannabis Cultivars Using SpiderMass. Metabolites, 12(6), 480. https://doi.org/10.3390/metabo12060480