
Lipidome Alterations following Mild Traumatic Brain Injury in the Rat

 ,
, 
Abstract
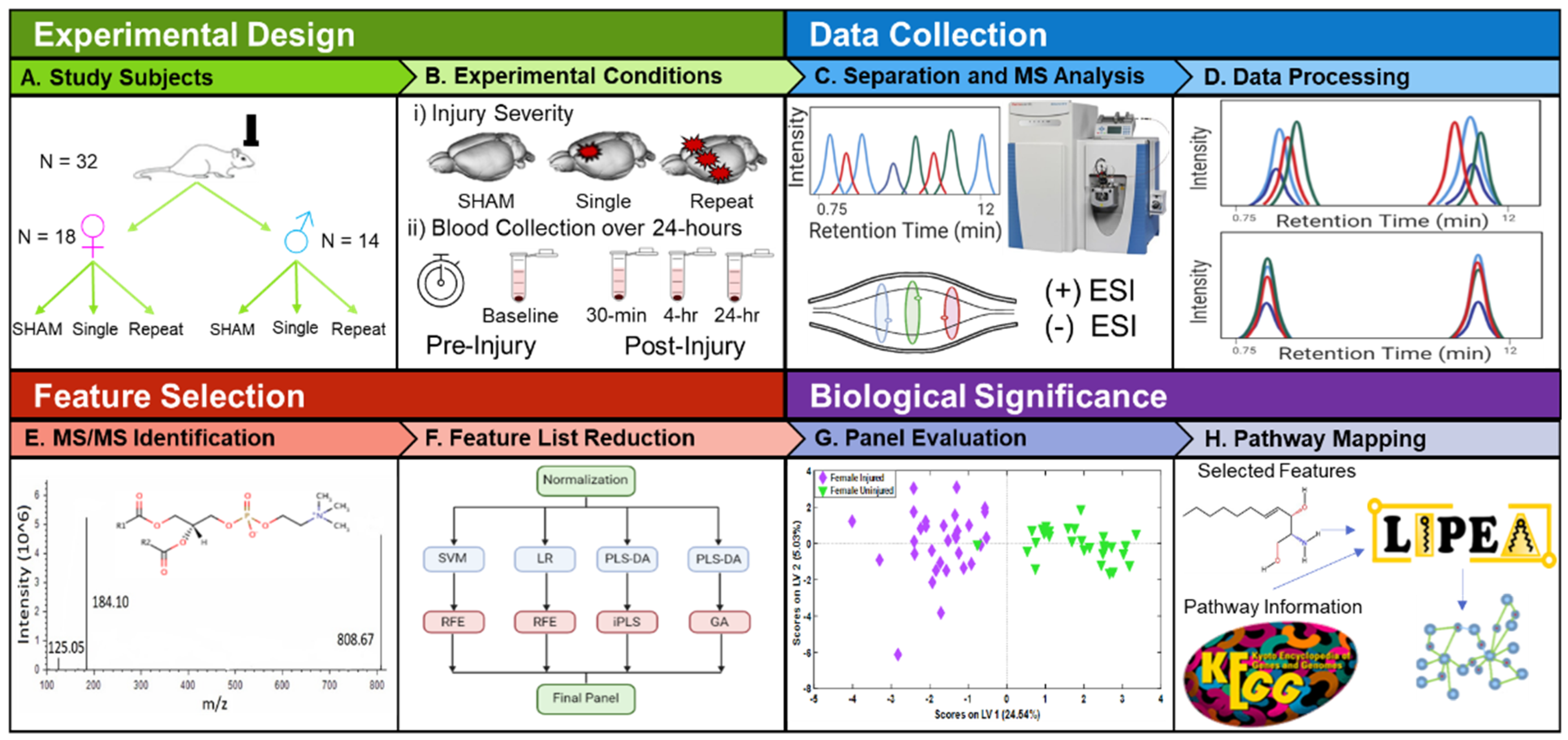
:1. Introduction
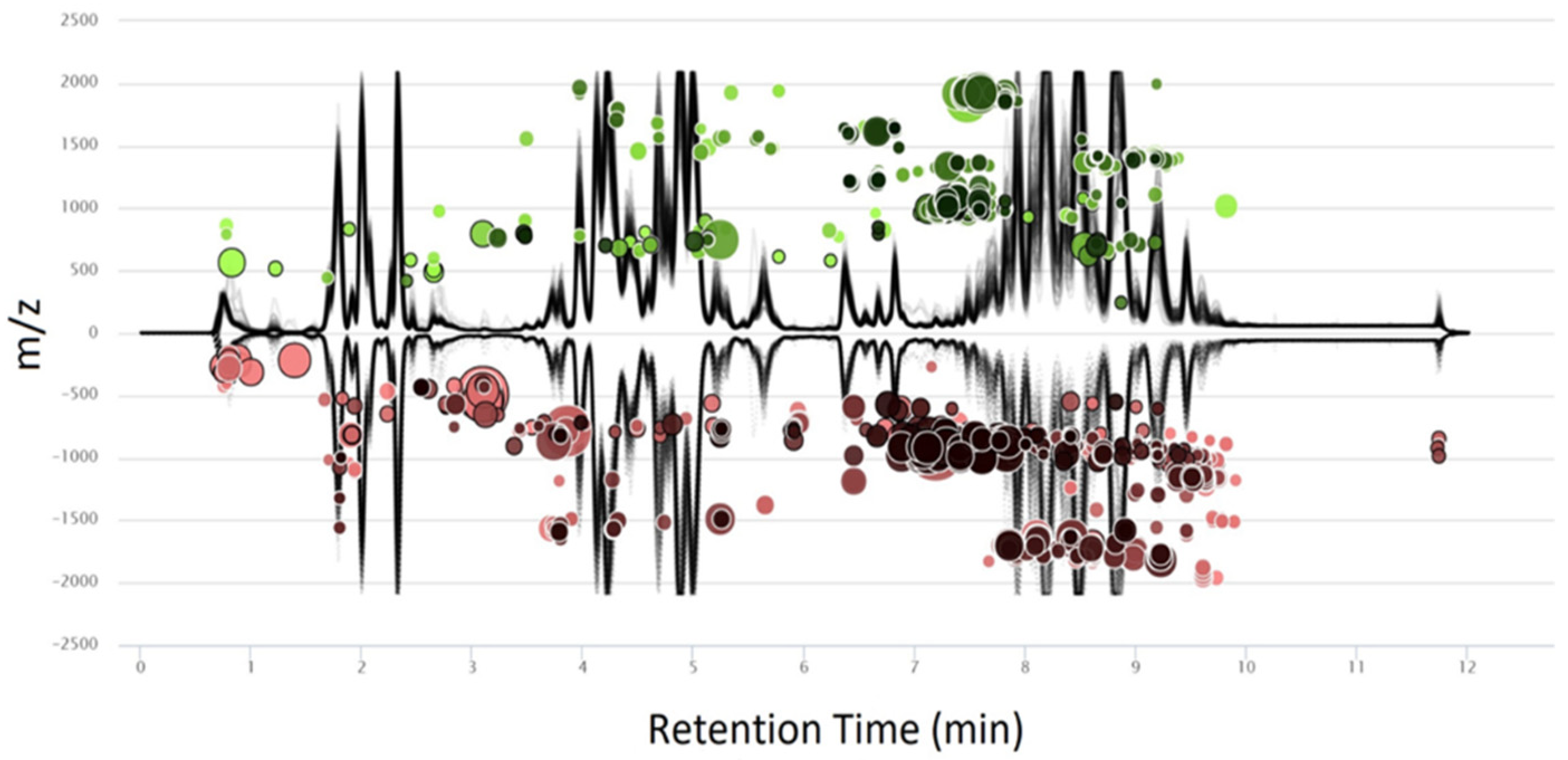
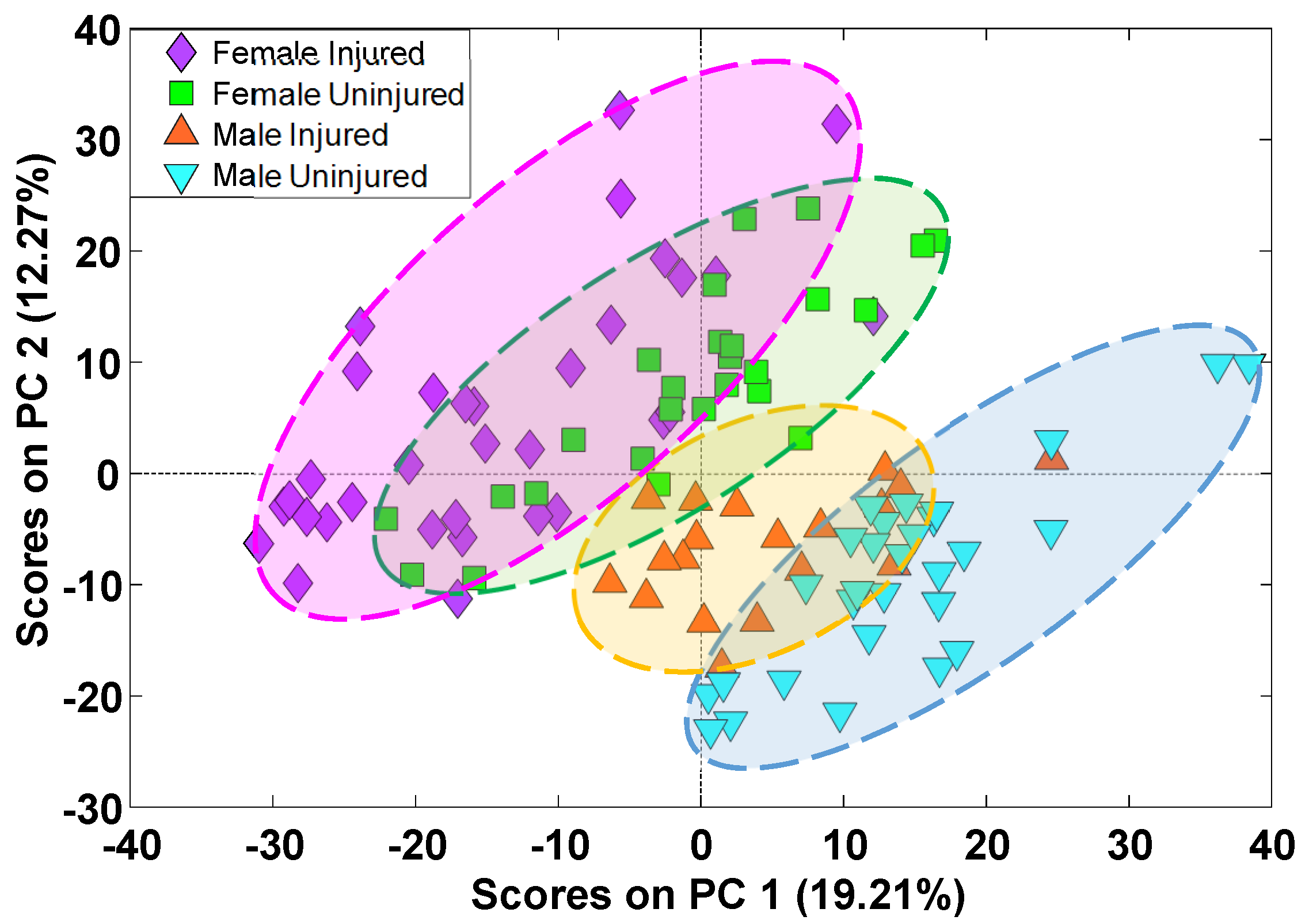
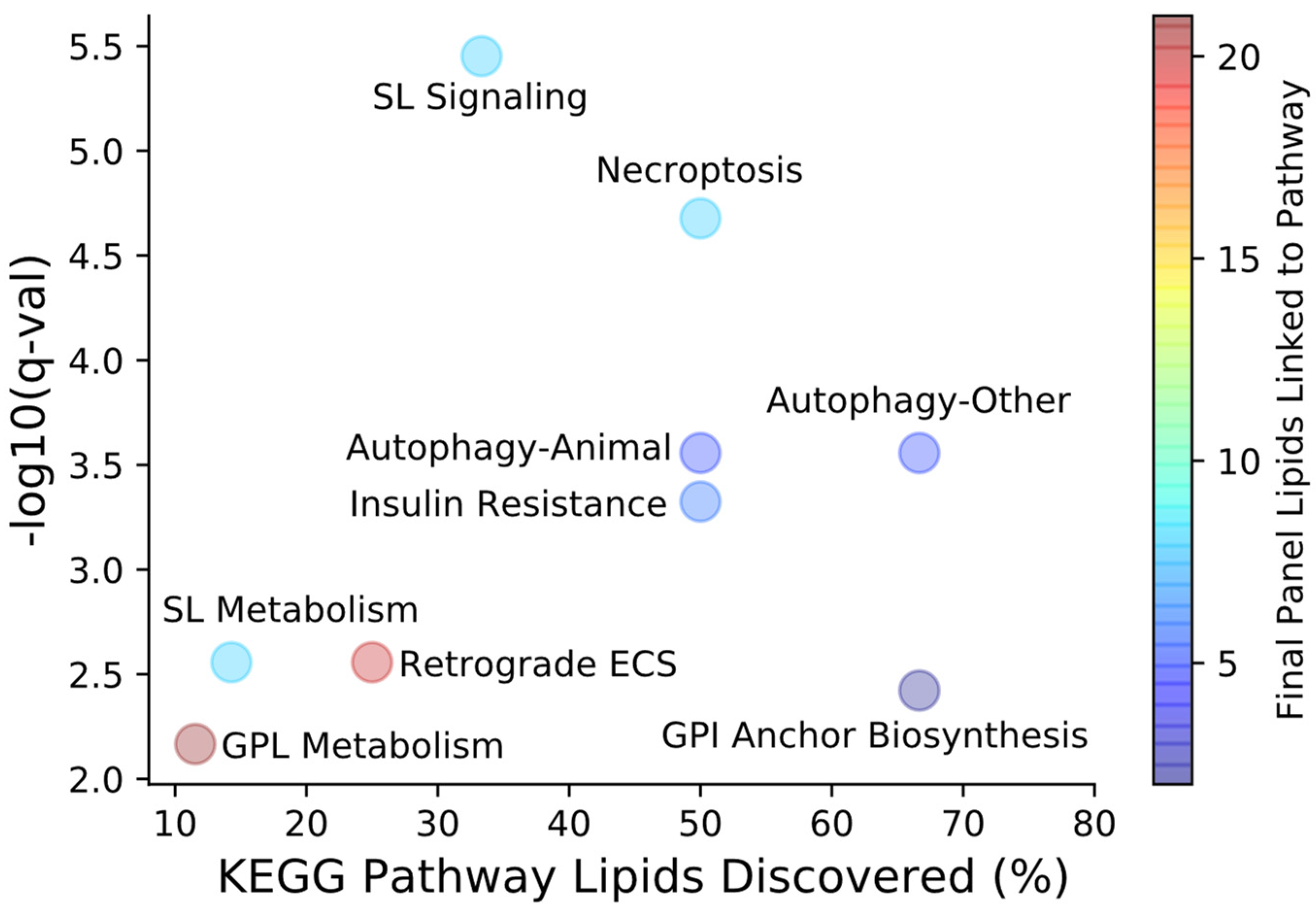
2. Results
3. Discussion
4. Materials and Methods
4.1. Chemicals
4.2. Injury Protocol and Blood Collection
4.3. Sample Preparation and Ultrahigh Performance Liquid Chromatography-Mass Spectrometry (UHPLC-MS) Analysis
4.4. Data Processing
4.5. Feature Selection and Pathway Mapping
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Peterson, A.; Xu, L.; Daugherty, J.; Breiding, M. Surveillance Report of Traumatic Brain Injury-Related Emergency Department Visits, Hospitalizations, and Deaths. Centers for Disease Control and Prevention. 2014. Available online: https://stacks.cdc.gov/view/cdc/78062 (accessed on 10 May 2020).
- Faul, M.; Xu, L.; Wald, M.M.; Coronado, V.G. Traumatic Brain Injury in the United States: Emergency Department Visits, Hospitalizations and Deaths. Centers for Disease Control and Prevention. 2010. Available online: https://www.cdc.gov/traumaticbraininjury/pubs/index.html (accessed on 10 May 2020).
- Arciniegas, D.; Held, K.; Wagner, P. Cognitive Impairment Following Traumatic Brain Injury. Curr. Treat. Options Neurol. 2002, 4, 43–57. [Google Scholar] [CrossRef] [PubMed]
- Rigon, A.; Turkstra, L.S.; Mutlu, B.; Duff, M.C. Facial-Affect Recognition Deficit as a Predictor of Different Aspects of Social-Communication Impairment in Traumatic Brain Injury. Neuropsychology 2018, 4, 476–483. [Google Scholar] [CrossRef] [PubMed]
- Dams-O’Connor, K.; Guetta, G.; Hahn-Ketter, A.E.; Fedor, A. Traumatic Brain Injury as a Risk Factor for Alzheimer’s Disease: Current Knowledge and Future Directions. Neurodegener. Dis. Manag. 2016, 6, 417–429. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fleminger, S.; Oliver, D.; Lovestone, S.; Rabe-Hesketh, S.; Giora, A. Head Injury as a Risk Factor for Alzheimer’s Disease: The Evidence 10 Years on a Partial Replication. J. Neurol. Neurosurg. Psychiatry 2003, 74, 857–862. [Google Scholar] [CrossRef]
- Salinsky, M.; Storzbach, D.; Goy, E.; Evrard, C. Traumatic Brain Injury and Psychogenic Seizures in Veterans. J. Head Trauma Rehabil. 2015, 30, E65–E70. [Google Scholar] [CrossRef]
- Lucke-Wold, B.P.; Nguyen, L.; Turner, R.C.; Logsdon, A.F.; Chen, Y.-W.; Smith, K.E.; Huber, J.D.; Matsumo, R.; Rosen, C.L.; Tucker, E.S.; et al. Traumatic Brain Injury and Epilepsy: Underlying Mechanisms Leading to Seizure. Seizure 2015, 33, 13–23. [Google Scholar] [CrossRef] [Green Version]
- Qureshi, K.L.; Upthegrove, R.; Toman, E.; Sawlani, V.; Davies, D.J.; Belli, A. Post-Traumatic Stress Disorder in UK Civilians with Traumatic Brain Injury: An Observational Study of Traumatic Brain Injury Clinic Attendees to Estimate PTSD Prevalence and its Relationship with Radiological Markers of Brain Injury Severity. BMJ Open 2019, 9, e021675. [Google Scholar] [CrossRef] [Green Version]
- Prasad, K.N.; Bondy, S.C. Common Biochemical Defects Linkage Between Post-Traumatic Stress Disorders, Mild Traumatic Brain Injury (TBI) and Penetrating TBI. Brain Res. 2015, 1599, 103–114. [Google Scholar] [CrossRef] [Green Version]
- Sternbach, G.L. The Glasgow Coma Scale. J. Emerg. Med. 2000, 19, 67–71. [Google Scholar] [CrossRef]
- Amyot, F.; Arciniegas, D.B.; Brazaitis, M.P.; Curley, K.C.; Diaz-Arrastia, R.; Gandjbakhche, A.; Herscovitch, P.; Hinds, S.R.; Manley, G.T.; Pacifico, A.; et al. A Review of the Effectiveness of Neuroimaging Modalities for the Detection of Traumatic Brain Injury. J. Neurotrauma 2015, 32, 1693–1721. [Google Scholar] [CrossRef] [Green Version]
- Mondello, S.; Schmid, K.; Rachel, B.P.; Kobeissy, F.; Italiano, D.; Jeromin, A.; Hayes, R.L.; Tortella, F.C.; Buki, A. The Challenge of Mild Traumatic Brain Injury: Role of Biochemical Markers in Diagnosis of Brain Damage. Med. Res. Rev. 2014, 34, 503–531. [Google Scholar] [CrossRef] [PubMed]
- Hajiaghamemar, M.; Seidi, M.; Oeur, R.A.; Margulies, S.S. Toward Development of Clinically Translatable Diagnostic and Prognostic Metrics of Traumatic Brain Injury Using Animal Models: A Review and a Look Forward. Exp. Neurol. 2019, 318, 101–123. [Google Scholar] [CrossRef] [PubMed]
- Rusnak, M. Traumatic Brain Injury: Giving Voice to a Silent Epidemic. Nat. Rev. Neurol. 2013, 9, 186–187. [Google Scholar] [CrossRef] [PubMed]
- Gerberding, J.; Binder, S. Report to Congress on Mild Traumatic Brain Injury in the United States: Steps to Prevent a Serious Public Health Problem; Centers for Disease Control and Prevention: Atlanta, GA, USA, 2003.
- Oris, C.; Pereira, B.; Duriff, J.; Simon-Pimmel, J.; Castellani, C.; Manzano, S.; Sapin, M.; Damien, B. The Biomarker S100B and Mild Traumatic Brain Injury: A Meta-analysis. Pediatrics 2018, 141, e20180037. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thelin, E.; Nimer, F.A.; Frostell, A.; Zetterberg, H.; Blennow, K.; Nystrom, H.; Svensson, M.; Bellander, B.-M.; Piehl, F.; Nelson, D.W. A Serum Protein Biomarker Panel Improves Outcome Prediction in Human Traumatic Brain Injury. Neurotrauma 2019, 2862, 2850–2862. [Google Scholar] [CrossRef] [Green Version]
- US Food and Drug Administration. Evaluation of Automatic Class III Designation for Banyan Brain Trauma Indicator: Decision Memorandum; US Food and Drug Administration: Silver Spring, MD, USA, 2018.
- Lagerstedt, L.; Egea-Guerrero, J.J.; Bustamante, A.; Rodriguez-Rodriguez, A.; Rahal, A.E.; Quintana-Diaz, M.; García-Armengol, R.; Prica, C.M.; Andereggen, E.; Rinaldi, L.; et al. Combining H-FABP and GFAP Increases the Capacity to Differentiate Detween CT-positive and CT-negative Patients with Mild Traumatic Brain Injury. PLoS ONE 2018, 13, e0200394. [Google Scholar] [CrossRef]
- Obermeier, B.; Daneman, R.; Ransohoff, R.M. Development, Maintenance and Disruption of the Blood-Brain Barrier. Nat. Med. 2013, 19, 1584–1596. [Google Scholar] [CrossRef]
- Hamilton, J.A.; Hillard, C.J.; Spector, A.A.; Watkins, P.A. Brain Uptake and Utilization of Fatty Acids, Lipids and Lipoproteins: Application to Neurological Disorders. J. Mol. Neurosci. 2007, 33, 2–11. [Google Scholar] [CrossRef]
- Pardridge, W.M. The Blood-Brain Barrier: Bottleneck in Brain Drug Development. NeuroRx 2005, 2, 3–14. [Google Scholar] [CrossRef]
- Roux, A.; Muller, L.; Jackson, S.N.; Post, J.; Baldwin, K.; Hoffer, B.; Balaban, C.D.; Barbacci, D.; Schultz, J.A.; Gouty, S.; et al. Mass Spectrometry Imaging of Rat Brain Lipid Profile Changes Over Time Following Traumatic Brain Injury. J. Neurosci. Methods 2016, 272, 19–32. [Google Scholar] [CrossRef] [Green Version]
- Sheth, S.A.; Iavarone, A.T.; Liebeskind, D.S.; Won, S.K.; Swanson, R.A. Targeted Lipid Profiling Discovers Plasma Biomarkers of Acute Brain Injury. PLoS ONE 2015, 10, e0129735. [Google Scholar] [CrossRef] [PubMed]
- Fiandaca, M.S.; Mapstone, M.; Mahmoodi, A.; Gross, T.; Macciardi, F.; Cheema, A.K.; Merchant-Borna, K.; Bazarian, J.; Federoff, H.J. Plasma Metabolomic Biomarkers Accurately Classify Acute Mild Traumatic Brain Injury from Controls. PLoS ONE 2018, 13, e0195318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hogan, S.R.; Phan, J.H.; Alvarado-Velez, M.; Wang, M.D.; Bellamkonda, R.V.; Fernández, F.M.; LaPlaca, M.C. Discovery of Lipidome Alterations Following Traumatic Brain Injury via High-Resolution Metabolomics. J. Proteome Res. 2018, 17, 2131–2143. [Google Scholar] [CrossRef]
- Gowda, H.; Ivanisevic, J.; Johnson, C.H.; Kurczy, M.E.; Benton, P.H.; Rinehart, D.; Nguyen, T.; Ray, J.; Kuehl, J.; Arevalo, B.; et al. Interactive XCMS online: Simplifying advanced metabolomic data processing and subsequent statistical analyses. Anal. Chem. 2014, 86, 6931–6939. [Google Scholar] [CrossRef] [PubMed]
- Bahado-Singh, R.O.; Grahm, S.; Han, B.; Turkoglu, O.; Ziadeh, J.; Mandal, R.; Er, A.; Wishart, D.S.; Stahel, P.L. Serum Metabolomic Markers for Traumatic Brain Injury: A Mouse Model. Metabolomics 2016, 12, 100. [Google Scholar] [CrossRef]
- Mallah, K.; Quanico, J.; Trede, D.; Kobeissy, F.; Zibara, K.; Salzet, M.; Fournier, I. Lipid Changes Associated with Traumatic Brain Injury Revealed by 3D MALDI-MSI. Anal. Chem. 2018, 90, 10568–10576. [Google Scholar] [CrossRef]
- Miller, M.R.; Robinson, M.; Bartha, R.; Stewart, T.C.; Fischer, L.; Dekaban, G.A.; Menson, R.S.; Shoemaker, K.J.; Fraser, D.D. Concussion Acutely Decreases Plasma Glycerophospholipids in Adolescent Male Athletes. J. Neurotrauma 2021, 38, 1608–1614. [Google Scholar] [CrossRef]
- Daley, M.; Dekaban, G.; Bartha, R.; Brown, A.; Stewart, T.C.; Doherty, T.; Fischer, L.; Holmes, J.; Menon, R.S.; Rupar, C.A.; et al. Metabolomics Profiling of Concussion in Adolescent Male Hockey Players: A Novel Diagnostic Method. Metabolomics 2016, 12, 185. [Google Scholar] [CrossRef]
- Homayoun, P.; Parkins, N.E.; Soblosky, J.; Carey, M.E.; Rodriguez De Turco, E.B.; Bazan, N.G. Cortical Impact Injury in Rats Promotes a Rapid and Sustained Increase in Polyunsaturated Free Fatty Acids and Diacylglycerols. Neurochem. Res. 2000, 25, 269–276. [Google Scholar] [CrossRef]
- Acevedo, A.; Durán, C.; Ciucci, S.; Gerl, M.; Cannistraci, C.V. LIPEA: Lipid Pathway Enrichment Analysis. bioRxiv 2018, biorxiv:274969. [Google Scholar] [CrossRef] [Green Version]
- Barbacci, D.C.; Roux, A.; Muller, L.; Jackson, S.N.; Post, J.; Baldwin, K.; Hoffer, B.; Balaban, C.D.; Schultz, J.A.; Gouty, S.; et al. Mass Spectrometric Imaging of Ceramide Biomarkers Tracks Therapeutic Response in Traumatic Brain Injury. ACS Chem. Neurosci. 2017, 8, 2266–2274. [Google Scholar] [CrossRef] [PubMed]
- Kitatani, K.; Idkowiak-Baldys, J.; Hannun, Y.A. The Sphingolipid Salvage Pathway in Ceramide Metabolism and Signaling. Cell Signal. 2008, 20, 1010–1018. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.K.; Jin, H.E.; Park, M.H.; Kiom, B.; Lee, P.H.; Nakauchi, H.; Carter, J.E.; He, X.; Schuchman, E.H.; Bae, J.S. Acid Sphingomyelinase Modulates the Autophagic Process by Controlling Lysosomal Biogenesis in Alzheimer’s Disease. J. Exp. Med. 2014, 211, 1551–1570. [Google Scholar] [CrossRef] [Green Version]
- Gulbins, E.; Palmada, M.; Reichel, M.; Lüth, A.; Böhmer, C.; Amato, D.; Müller, C.P.; Tischbirek, C.H.; Groemer, T.W.; Tabatabai, G.; et al. Acid Sphingomyelinase–Ceramide System Mediates Effects of Antidepressant Drugs. Nat. Med. 2013, 19, 2–9. [Google Scholar] [CrossRef] [Green Version]
- Niziolek, G.M.; Hoehn, R.S.; Seitz, A.P.; Jernigan, P.L.; Makley, A.T.; Gulbins, E.; Edwards, M.J.; Goodman, M.D. The Role of Acid Sphingomyelinase Inhibition in Repetitive Mild Traumatic Brain Injury. J. Surg. Res. 2020, 259, 296–304. [Google Scholar] [CrossRef]
- Dawson, G. Measuring Brain Lipids. Biochim. Biophys. Acta. 2016, 1851, 1026–1039. [Google Scholar] [CrossRef] [Green Version]
- Xie, Y.; Li, J.; Kang, R.; Tang, D. Interplay Between Lipid Metabolism and Autophagy. Front Cell Dev. Biol. 2020, 8, 431. [Google Scholar] [CrossRef]
- Saito, T.; Kuma, A.; Sugiura, Y.; Ichimura, Y.; Obata, M.; Kitamura, H.; Okuda, S.; Lee, H.C.; Ikeda, K.; Kanegae, Y.; et al. Autophagy Regulates Lipid Metabolism through Selective Turnover of NCoR1. Nat. Commun. 2019, 10, 1567. [Google Scholar] [CrossRef] [Green Version]
- Luo, C.L.; Li, B.X.; Li, Q.Q.; Chen, X.P.; Sun, Y.X.; Bao, H.J.; Dai, D.K.; Shen, Y.W.; Xu, H.F.; Ni, H.; et al. Autophagy is Involved in Traumatic Brain Injury-Induced Cell Death and Contributes to Functional Outcome Deficits in Mice. Neuroscience 2011, 184, 54–63. [Google Scholar] [CrossRef]
- Liu, C.L.; Chen, S.; Dietrich, D.; Hu, B.R. Changes in Autophagy After Traumatic Brain Injury. J. Cereb. Blood Flow Metab. 2008, 28, 674–683. [Google Scholar] [CrossRef]
- Clark, R.S.B.; Bayir, H.; Chu, C.T.; Alber, S.M.; Kochanek, P.M.; Watkins, S.C. Autophagy is Increased in Mice After Traumatic Brain Injury and is Detectable in Human Brain After Trauma and Critical Illness. Autophagy 2008, 8627, 88–90. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Wang, H. Autophagy in Traumatic Brain Injury: A New Target for Therapeutic Intervention. Front Mol. Neurosci. 2018, 11, 190. [Google Scholar] [CrossRef]
- Zhang, M.; Shan, H.; Chang, P.; Wang, T.; Dong, W.; Chen, X.; Tao, L. Hydrogen Sulfide Offers Neuroprotection on Traumatic Brain Injury in Parallel with Reduced Apoptosis and Autophagy in Mice. PLoS ONE 2014, 9, e87241. [Google Scholar]
- Xu, J.; Wang, H.; Lu, X.; Ding, K.; Zhang, L.; He, J. Posttraumatic Administration of Luteolin Protects Mice from Traumatic Brain Injury: Implication of Autophagy and Inflammation. Brain Res. 2014, 1582, 237–246. [Google Scholar] [CrossRef]
- Zhang, L.; Wang, H.; Fan, W.; Gao, Y.; Li, X.; Hu, Z.; Ding, K.; Wang, Y.; Wang, X. Fucoxanthin Provides Neuroprotection in Models of Traumatic Brain Injury via the Nrf2- ARE and Nrf2-Autophagy Pathways. Sci. Rep. 2017, 7, 46763. [Google Scholar] [CrossRef] [Green Version]
- Pasvogel, A.E.; Miketova, P.; Moore, I.M.K. Cerebrospinal Fluid Phospholipid Changes Following Traumatic Brain Injury. Biol. Res. Nurs. 2008, 10, 113–120. [Google Scholar] [CrossRef]
- Pasvogel, A.E.; Miketova, P.; Moore, I.M.K. Differences in CSF Phospholipid Concentration by Traumatic Brain Injury Outcome. Biol. Res. Nurs. 2010, 11, 325–331. [Google Scholar] [CrossRef]
- Manning, Y.K.; Llera, A.; Dekaban, G.A.; Bartha, R.; Barreira, C.; Brown, A.; Fischer, L.; Jevremovic, T.; Blackney, K.; Doherty, T.J.; et al. Linked MRI Signatures of the Brain’s Acute and Persistent Response to Concussion in Female Varsity Rugby Players. NeuroImage Clin. 2019, 21, 101627. [Google Scholar] [CrossRef]
- Nessel, I.; Michael-Titus, A.T. Lipid Profiling of Brain Tissue and Blood After Traumatic Brain Injury: A Review of Human and Experimental Studies. Semin. Cell Dev. Biol. 2021, 112, 145–156. [Google Scholar] [CrossRef]
- Pasparakis, M.; Vandenabeele, P. Necroptosis and its Role in Inflammation. Nature 2015, 5, 311–320. [Google Scholar] [CrossRef]
- Parisi, L.R.; Li, N.; Atilla-Gokcumen, G.E. Very Long Chain Fatty Acids Are Functionally Involved in Necroptosis Very Long Chain Fatty Acids Are Functionally Involved in Necroptosis. Cell Chem. Biol. 2017, 24, 1445–1454. [Google Scholar] [CrossRef] [Green Version]
- Sawai, H.; Ogiso, H.; Okazaki, T. Differential Changes in Sphingolipids Between TNF-Induced Necroptosis and Apoptosis in U937 Cells and Necroptosis-Resistant Sublines. Leuk. Res. 2015, 39, 964–970. [Google Scholar] [CrossRef]
- Sekar, S.; Viswas, R.S.; Mahabadi, H.M.; Alizadeh, E.; Fonge, H.; Taghibiglou, C. Concussion/Mild Traumatic Brain Injury (TBI) Induces Brain Insulin Resistance: A Positron Emission Tomography (PET) Scanning Study. Int. J. Mol. Sci. 2021, 22, 9005. [Google Scholar] [CrossRef]
- Mowery, N.; Gunter, O.L.; Guillamondegui, O.; Dossett, L.A.; Dortch, M.J.; Morris, J.A.; May, A.K. Stress Insulin Resistance is a Marker for Mortality in Traumatic Brain Injury. Trauma Acute Care Surg. 2009, 66, 145–153. [Google Scholar] [CrossRef]
- Kotronen, A.; Velagapudi, V.R.; Yetukuri, L.; Westerbacka, J.; Bergholm, R.; Ekroos, K.; Makkonen, J.; Taskinen, M.R.; Orešič, M.; Yki-Järvinen, H.; et al. Serum Saturated Fatty Acids Containing Triacylglycerols are Better Markers of Insulin Resistance than Total Serum Triacylglycerol Concentrations. Diabetologia 2009, 52, 684–690. [Google Scholar] [CrossRef] [Green Version]
- Barzó, P.; Marmarou, A.; Fatouros, P.; Corwin, F.; Dunbar, J. Magnetic Resonance Imaging-Monitored Acute Blood-Brain Barrier Changes in Experimental Traumatic Brain Injury. J. Neurosurg. 1996, 85, 1113–1121. [Google Scholar] [CrossRef]
- Shapira, Y.; Setton, D.; Artru, A.A.; Shohami, E. Blood-Brain Barrier Permeability, Cerebral Edema, and Neurologic Function After Closed Head Injury in Rats. Anesth. Analg. 1993, 77, 141–148. [Google Scholar] [CrossRef]
- Jankevics, A.; Jenkins, A.; Dunn, W.B.; Najdekr, L. An Improved Strategy for Analysis of Lipid Molecules Utilising a Reversed Phase C30 UHPLC Column and Scheduled MS/MS Acquisition. Talanta 2021, 229, 122262. [Google Scholar] [CrossRef]
- Antignac, J.P.; De Wasch, K.; Monteau, F.; De Brabander, H.; Andre, F.; Le Bizec, B. The Ion Suppression Phenomenon in Liquid Chromatography-Mass Spectrometry and its Consequences in the Field of Residue Analysis. Anal. Chim. Acta. 2005, 529, 129–136. [Google Scholar] [CrossRef]
- Fahy, E.; Sud, M.; Cotter, D.; Subramaniam, S. LIPID MAPS Online Tools for Lipid Research. Nucleic Acids Res. 2007, 35, 606–612. [Google Scholar] [CrossRef] [Green Version]
- Fahy, E.; Subramaniam, S.; Murphy, R.C.; Nishijima, M.; Raetz, C.R.; Shimizu, T.; Spener, F.; van Meer, G.; Wakelam, M.J.; Dennis, E.A. Update of the LIPID MAPS Comprehensive Classification System for Lipids. J. Lipid Res. 2009, 50, 9–14. [Google Scholar] [CrossRef] [Green Version]
- Wishart, D.S.; Feunang, Y.D.; Marcu, A.; Guo, A.C.; Liang, K.; Vázquez-Fresno, R.; Sajed, T.; Johnson, D.; Li, C.; Karu, N.; et al. HMDB 4.0: The Human Metabolome Database for 2018. Nucleic Acids Res. 2018, 46, D608–D617. [Google Scholar] [CrossRef]
- Pedregosa, F.; Varoquaux, G.; Gramfort, A.; Michel, V.; Thirion, B. Scikit-learn: Machine Learning in Python. J. Mach. Learn. Res. 2011, 12, 2825–2830. [Google Scholar]
- Guenther, N.; Schonlau, M. Support Vector Machines. Stata J. 2016, 16, 917–937. [Google Scholar] [CrossRef] [Green Version]
- Li, F.; Yang, Y. Analysis of Recursive Feature Elimination Methods. In Proceedings of the 28th Annual International ACM SIGIR Conference on Research and Development in Information Retrieval (SIGIR), Salvador, BA, Brazil, 15–19 August 2005; pp. 633–634. [Google Scholar]
- Sato, T.; Takano, Y.; Miyashiro, R.; Yoshise, A. Feature Subset Selection for Logistic Regression via Mixed Integer Optimization. Comput. Optim. Appl. 2016, 64, 865–880. [Google Scholar] [CrossRef] [Green Version]
- Thissen, D.; Steinberg, L.; Kuang, D. Quick and Easy Implementation of the Benjamini-Hochberg Procedure for Controlling the False Positive Rate in Multiple Comparisons. J. Educ. Behav. Stat. 2002, 27, 77–83. [Google Scholar] [CrossRef]
- De Andrade, B.M.; De Gois, J.S.; Xavier, V.L.; Luna, A.S. Comparison of the Performance of Multiclass Classifiers in Chemical Data: Addressing the Problem of Overfitting with the Permutation Test. Chemom. Intell. Lab. Syst. 2020, 201, 104013. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Classifier | Feature Selection Method | Sex | Number of Features | Cross-Validation Estimate, AUC (SD) | All Samples, AUC | Selected Features |
|---|---|---|---|---|---|---|
| Linear SVM | RFE | M | 27 | 0.875 (0.133) | 0.980 | 63, 89, 244, 258, 365, 378, 417, 453, 457, 459, 476, 497, 527, 541, 543, 551, 570, 635, 651, 788, 792, 798, 808, 857, 967, 1095, 1114, |
| Logistic Regression | RFE | M | 24 | 0.840 (0.174) | 0.992 | 88, 89, 183, 279, 365, 453, 457, 459, 473, 476, 486, 502, 527, 543, 551, 570, 601, 651, 652, 788, 792, 808, 1104, 1114 |
| oPLS-DA | GA | M | 31 | 0.941 (0.062) | 1.000 | 17, 63, 161, 171, 174, 209, 278, 316, 365, 407, 494, 497, 513, 527, 531, 543, 550, 551, 567, 589, 601, 616, 621, 626, 627, 652, 745, 774, 788, 1080, 1114 |
| oPLS-DA | iPLS | M | 20 | 0.891 (0.090) | 0.992 | 61, 101, 258, 273, 321, 346, 365, 473, 527, 543, 570, 617, 652, 851, 876, 951, 994, 998, 1008, 1095 |
| Linear SVM | RFE | F | 28 | 0.766 (0.140) | 0.953 | 8, 10, 35, 103, 104, 282, 328, 346, 348, 349, 388, 437, 457, 460, 490, 615, 757, 780, 784, 813, 825, 874, 875, 920, 989, 1026, 1044, 1110 |
| Logistic Regression | RFE | F | 29 | 0.752 (0.120) | 0.976 | 8, 35, 73, 81, 86, 103, 263, 282, 328, 346, 348, 388, 417, 437, 443, 455, 532, 620, 745, 757, 813, 825, 874, 875, 972, 988, 989, 1055, 1110 |
| oPLS-DA | GA | F | 29 | 0.949 (0.156) | 0.993 | 8, 27, 103, 154, 270, 378, 387, 408, 416, 455, 477, 531, 538, 550, 620, 647, 648, 652, 669, 712, 717, 719, 774, 825, 854, 869, 1082, 1095, 1110 |
| oPLS-DA | iPLS | F | 24 | 0.880 (0.110) | 0.943 | 27, 34, 141, 146, 149, 153, 299, 328, 381, 410, 425, 529, 590, 620, 621, 634, 675, 714, 751, 773, 842, 903, 936, 989 |
| Feature Number | Retention Time (min) | m/z Mass Error (ppm) | Detected Ion | Elemental Formula | Annotation | p-Value (TBI vs. Baseline) | Fold Change | Time |
|---|---|---|---|---|---|---|---|---|
| a | ||||||||
| 63 | 8.893 | 716.6343 −0.253 | [M+NH4]+ | C49H78O2 | CE(22:5) | 0.0655 | 1.340 | 4 h |
| 89 | 7.303 | 652.6605 −0.329 | [M+H]+ | C42H85NO3 | Cer(d18:0/24:0) | 0.0292 | −1.553 | 4 h |
| 258 | 2.186 | 601.3349 −0.518 | [M+H]+ | C27H53O12P | LysoPI(18:0) | 0.0147 | 1.190 | 24 h |
| 365 | 5.442 | 800.6168 −0.096 | [M+H]+ | C45H86NO8P | PC(18:2_19:0) | 0.245 | −1.042 | 30 min |
| 453 | 4.982 | 880.6071 0.446 | [M+HCO2]− | C48H86NO8P | PC(18:0_22:5) | 0.0483 | 1.207 | 4 h * |
| 459 | 4.742 | 878.5919 0.958 | [M+HCO2]− | C48H84NO8P | PC(18:0_22:6) | 0.0198 | 1.192 | 4 h |
| 476 | 4.337 | 846.6008 −0.494 | [M+H]+ | C49H84NO8P | PC(41:7) | 0.0323 | 1.425 | 4 h |
| 497 | 4.127 | 858.6014 0.153 | [M+H]+ | C50H84NO8P | PC(42:8) | 0.249 | 1.070 | 4 h |
| 527 | 4.773 | 718.5752 0.230 | [M+H]+ | C40H80NO7P | PC(O-16:1/16:0) | 0.146 | −1.603 | 4 h |
| 543 | 4.326 | 816.5910 0.387 | [M+H]+ | C48H82NO7P | PC(O-18:2_22:6) | 0.0375 | 1.493 | 30 min * |
| 551 | 5.557 | 772.6218 −0.197 | [M+H]+ | C44H86NO7P | PC(O-18:1/18:1) | 0.277 | 1.035 | 30 min |
| 570 | 5.768 | 798.6379 0.386 | [M+H]+ | C46H88NO7P | PC(O-38:3) | 0.221 | 1.054 | 24 h |
| 601 | 4.317 | 818.6062 −0.162 | [M+H]+ | C48H84NO7P | PC(O-18:1/22:6) | 0.00800 | 1.607 | 4 h ** |
| 651 | 5.588 | 704.5591 −0.433 | [M+H]+ | C39H78NO7P | PE(O-34:1) | 0.0156 | −1.605 | 4 h * |
| 652 | 6.297 | 704.5590 −0.504 | [M+H]+ | C39H78NO7P | PE(O-18:1/16:0) | 0.0144 | −1.656 | 4 h * |
| 788 | 3.676 | 689.5595 −0.217 | [M+H]+ | C38H77N2O6P | SM(d33:1) | 0.0953 | 1.153 | 24 h |
| 792 | 3.978 | 703.5759 0.797 | [M+H]+ | C39H79N2O6P | SM(d34:1) | 0.0335 | 1.296 | 24 h |
| 808 | 3.613 | 727.5758 0.606 | [M+H]+ | C41H79N2O6P | SM(d36:3) | 0.000266 | 1.632 | 4 h * |
| 1095 | 8.758 | 984.8954 −0.423 | [M+NH4]+ | C63H114O6 | TG(60:4) | 0.221 | −1.648 | 24 h |
| 1114 | 9.754 | 1014.9420 −0.782 | [M+NH4]+ | C65H120O6 | TG(18:1_20:1_24:1) | 0.0275 | −1.446 | 4 h |
| b | ||||||||
| 8 | 0.811 | 246.1701 0.284 | [M+H]+ | C12H23NO4 | Car(5:0) | 0.0137 | −1.374 | 30 min |
| 27 | 1.325 | 414.3215 0.313 | [M+H]+ | C23H43NO5 | Car(16:1-OH) | 0.0932 | 1.660 | 4 h |
| 35 | 1.701 | 442.3528 0.316 | [M+H]+ | C25H47NO5 | Car(18:1-OH) | 0.00239 | 2.019 | 4 h |
| 103 | 7.349 | 708.6512 0.868 | [M+HCO2]− | C43H85NO3 | Cer(d18:1/25:0) | 0.182 | 1.260 | 4 h |
| 282 | 3.997 | 718.5386 0.654 | [M+H]+ | C39H76NO8P | PE(16:0_18:1) | 0.00113 | −2.354 | 4 h ** |
| 328 | 4.598 | 772.5858 0.286 | [M+H]+ | C43H82NO8P | PC(17:0_18:2) | 0.156 | −1.407 | 24 h |
| 346 | 4.434 | 784.5856 −0.036 | [M+H]+ | C44H82NO8P | PC(18:1_18:2) | 0.0471 | −1.381 | 24 h |
| 348 | 4.471 | 828.5764 −1.111 | [M+HCO2]− | C44H82NO8P | PC(16:0_20:3) | 0.215 | 1.183 | 24 h |
| 388 | 5.752 | 814.6323 −0.277 | [M+H]+ | C46H88NO8P | PC(18:0_20:2) | 0.113 | 1.077 | 24 h |
| 437 | 4.243 | 864.5763 0.996 | [M+HCO2]− | C47H82NO8P | PC(17:0_22:6) | 0.0510 | 1.112 | 24 h |
| 455 | 4.646 | 880.6078 1.206 | [M+HCO2]− | C48H86NO8P | PC(18:0_22:5) | 0.125 | 1.382 | 30 min |
| 620 | 4.510 | 742.5392 0.712 | [M+H]+ | C41H76NO8P | PE(18:1_18:2) | 0.00600 | −2.169 | 24 h ** |
| 757 | 1.899 | 838.5572 −0.148 | [M+Na]+ | C44H82NO10P | PS(38:2) | 0.150 | −1.357 | 24 h |
| 813 | 5.578 | 759.6379 −0.112 | [M+H]+ | C43H87N2O6P | SM(d16:0_ 22:1) | 0.00597 | 1.456 | 4 h * |
| 825 | 5.316 | 771.6381 −0.113 | [M+H]+ | C44H87N2O6P | SM(d39:2) | 0.0143 | 1.619 | 24 h |
| 874 | 1.840 | 302.3054 −1.448 | [M+H]+ | C18H39NO2 | Sphinganine (C18) | 0.131 | −1.281 | 4 h |
| 875 | 1.699 | 300.2898 0.299 | [M+H]+ | C18H37NO2 | Sphingosine (C18) | 0.205 | −1.508 | 24 h |
| 989 | 8.176 | 898.7861 −0.280 | [M+NH4]+ | C57H100O6 | TG(18:1_18:2_18:2) | 0.0128 | −2.530 | 4 h |
| 1110 | 8.837 | 998.9114 −0.076 | [M+NH4]+ | C64H116O6 | TG(61:4) | 0.000911 | −2.341 | 4 h * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gier, E.C.; Pulliam, A.N.; Gaul, D.A.; Moore, S.G.; LaPlaca, M.C.; Fernández, F.M. Lipidome Alterations following Mild Traumatic Brain Injury in the Rat. Metabolites 2022, 12, 150. https://doi.org/10.3390/metabo12020150
Gier EC, Pulliam AN, Gaul DA, Moore SG, LaPlaca MC, Fernández FM. Lipidome Alterations following Mild Traumatic Brain Injury in the Rat. Metabolites. 2022; 12(2):150. https://doi.org/10.3390/metabo12020150
Chicago/Turabian StyleGier, Eric C., Alexis N. Pulliam, David A. Gaul, Samuel G. Moore, Michelle C. LaPlaca, and Facundo M. Fernández. 2022. "Lipidome Alterations following Mild Traumatic Brain Injury in the Rat" Metabolites 12, no. 2: 150. https://doi.org/10.3390/metabo12020150
APA StyleGier, E. C., Pulliam, A. N., Gaul, D. A., Moore, S. G., LaPlaca, M. C., & Fernández, F. M. (2022). Lipidome Alterations following Mild Traumatic Brain Injury in the Rat. Metabolites, 12(2), 150. https://doi.org/10.3390/metabo12020150