
Investigations into the In Vitro Metabolism of hGH and IGF-I Employing Stable-Isotope-Labelled Drugs and Monitoring Diagnostic Immonium Ions by High-Resolution/High-Accuracy Mass Spectrometry


Abstract
:1. Introduction

2. Results and Discussion
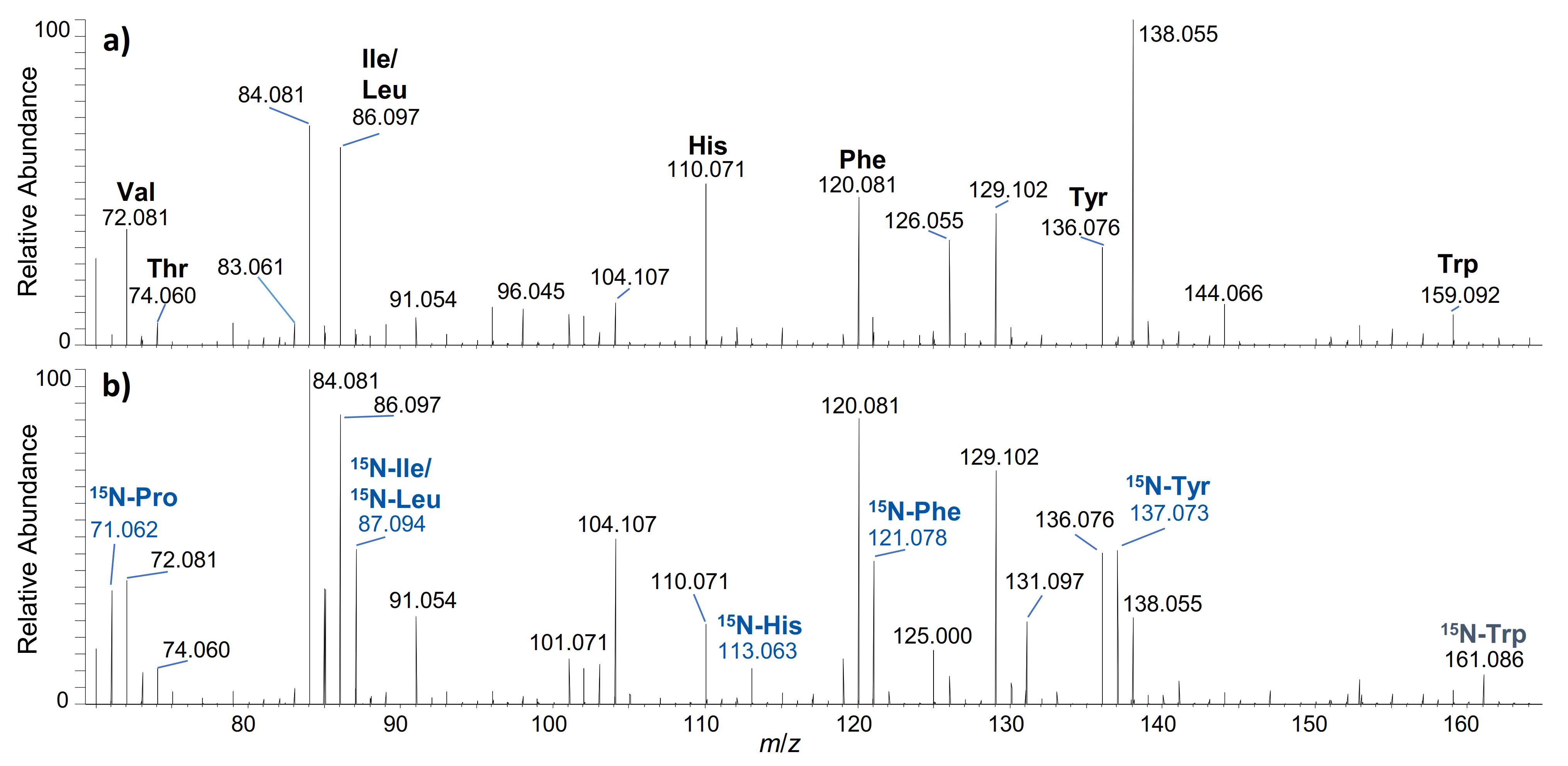
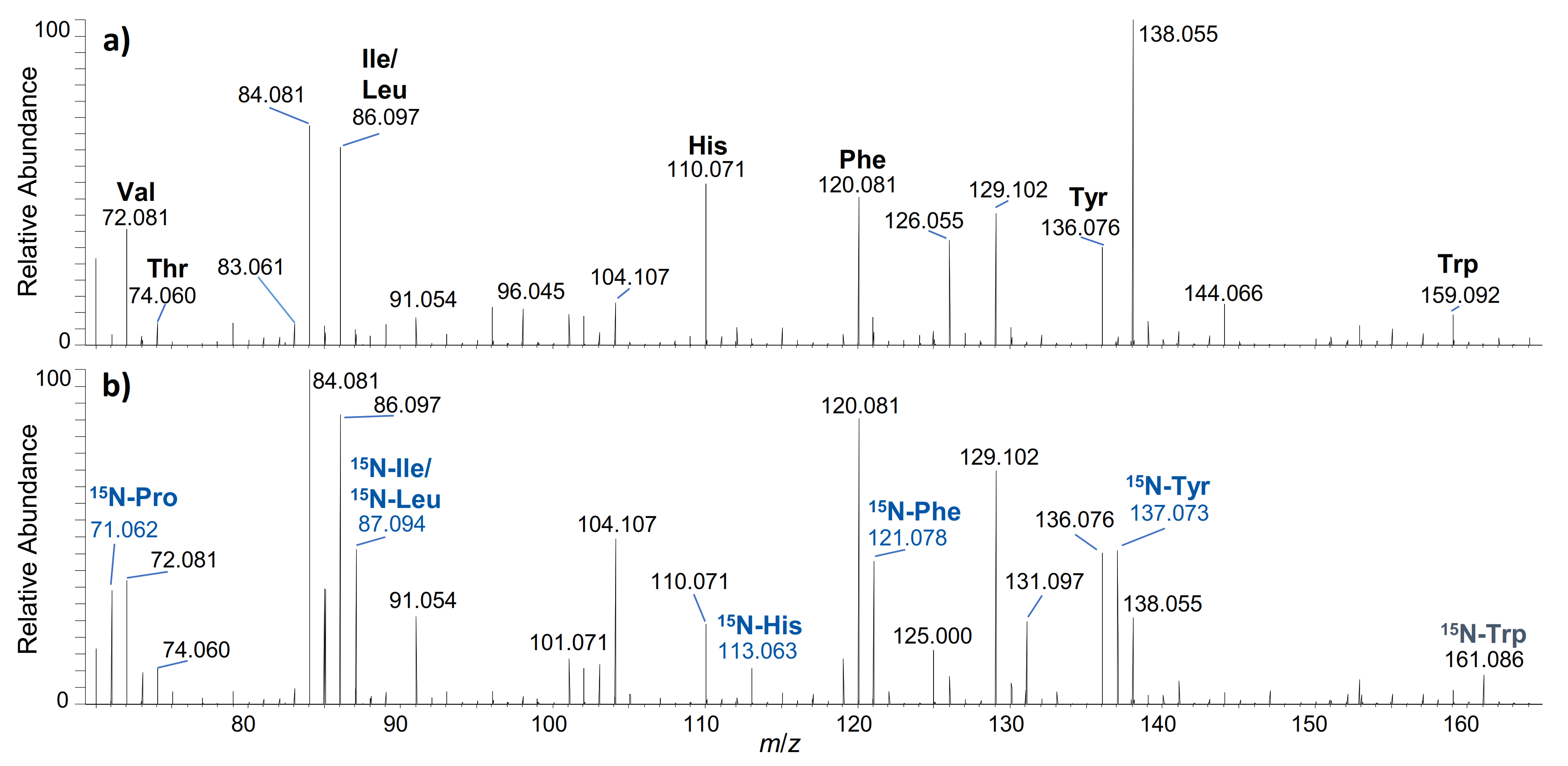
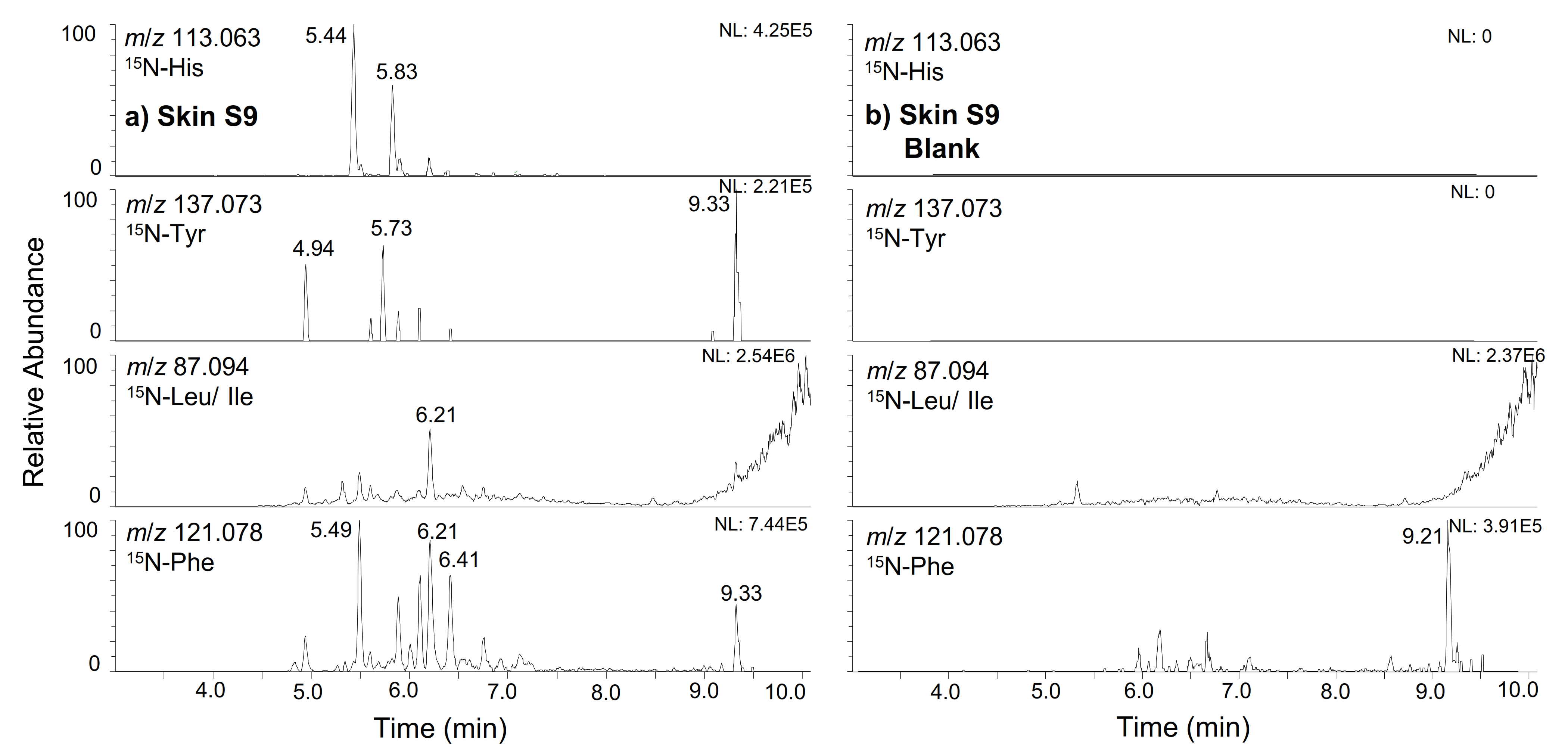
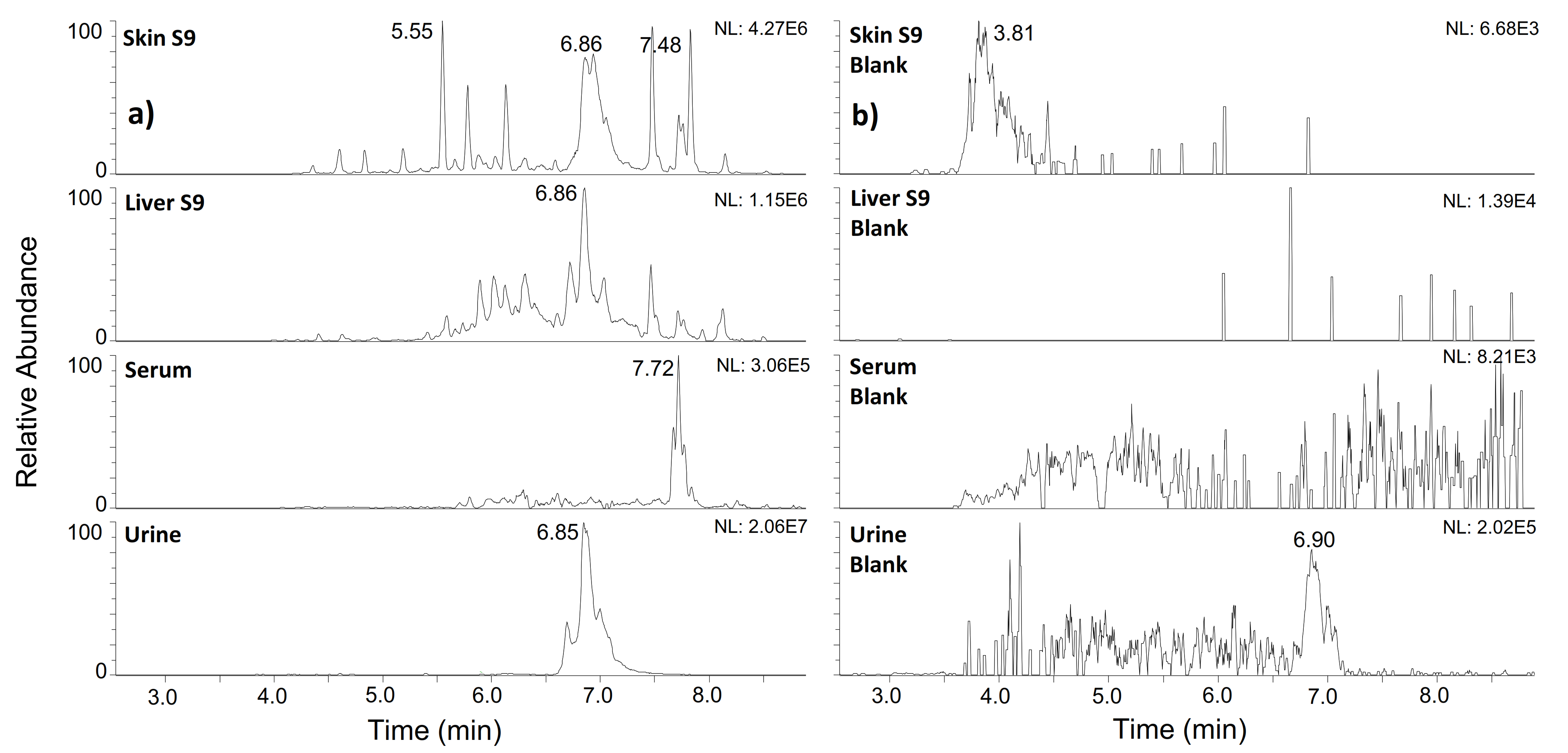
2.1. hGH
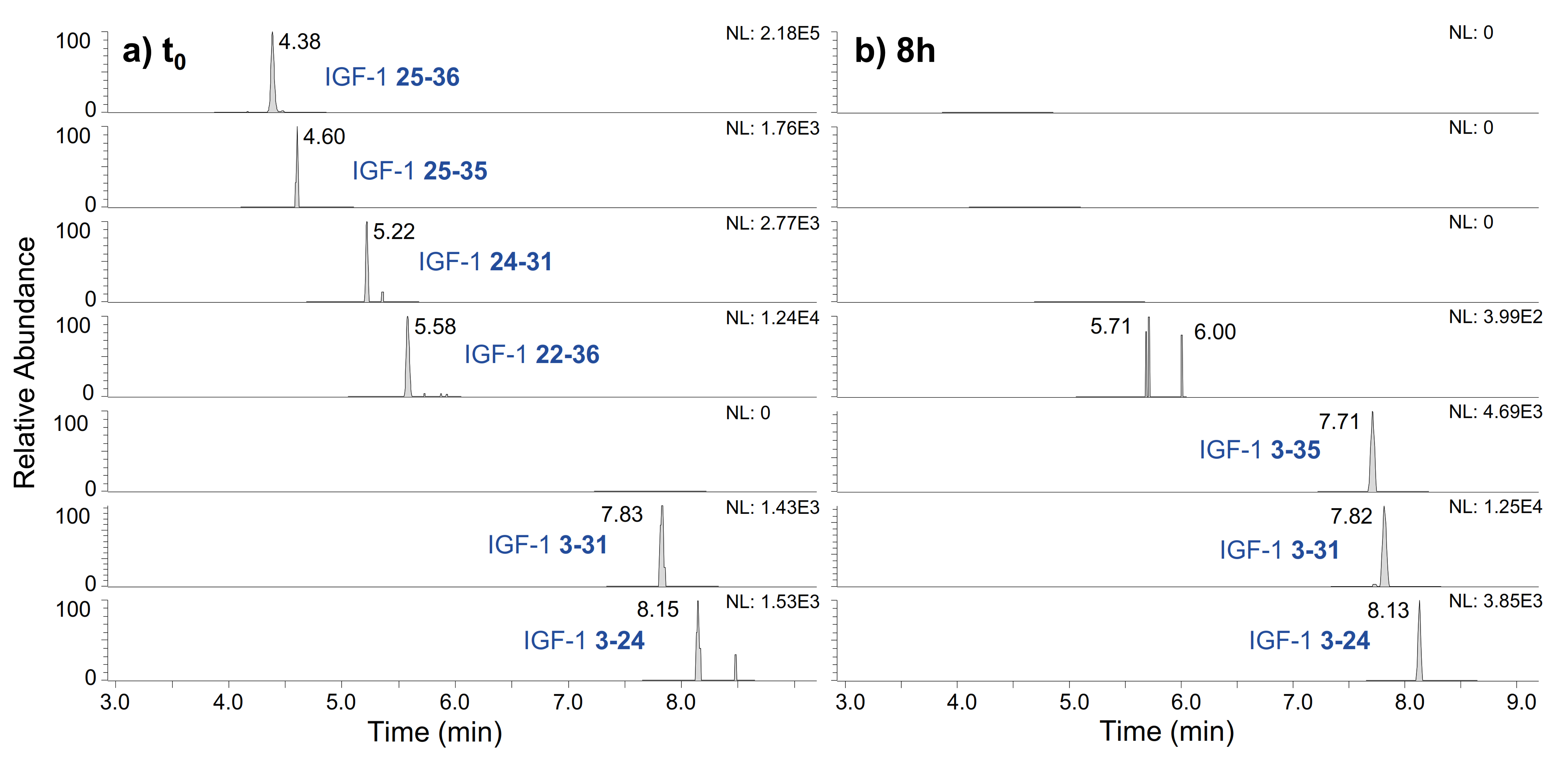
2.2. IGF-I
3. Materials and Methods
3.1. Chemicals and Reagents
3.2. In Vitro Experiments
3.3. Fragment Stability in Human Serum
3.4. Liquid Chromatography–High-Resolution Mass Spectrometry
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Thevis, M.; Kuuranne, T.; Geyer, H. Annual banned-substance review: Analytical approaches in human sports drug testing 2019/2020. Drug Test Anal. 2021, 13, 8–35. [Google Scholar] [CrossRef] [PubMed]
- Ayotte, C.; Miller, J.; Thevis, M. Challenges in modern anti-doping analytical science. Med. Sport Sci. 2017, 62, 68–76. [Google Scholar] [PubMed]
- Thevis, M.; Kuuranne, T.; Geyer, H. Annual banned-substance review - analytical approaches in human sports drug testing. Drug Test Anal. 2020, 12, 7–26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Butch, A.W.; Ahrens, B.D.; Avliyakulov, N.K. Urine reference intervals for human chorionic gonadotropin (hcg) isoforms by immunoextraction-tandem mass spectrometry to detect hcg use. Drug Test Anal. 2018, 10, 956–960. [Google Scholar] [CrossRef]
- Piper, T.; Geyer, H.; Nieschlag, E.; Bally, L.; Thevis, M. Carbon isotope ratios of endogenous steroids found in human serum-method development, validation, and reference population-derived thresholds. Anal. Bioanal. Chem. 2021, 413, 5655–5667. [Google Scholar] [CrossRef]
- Thevis, M.; Guddat, S.; Flenker, U.; Schanzer, W. Quantitative analysis of urinary glycerol levels for doping control purposes using gas chromatography-mass spectrometry. Eur. J. Mass Spectrom. 2008, 14, 117–125. [Google Scholar] [CrossRef]
- Barroso, O.; Handelsman, D.J.; Strasburger, C.; Thevis, M. Analytical challenges in the detection of peptide hormones for anti-doping purposes. Bioanalysis 2012, 4, 1577–1590. [Google Scholar] [CrossRef]
- van den Broek, I.; Blokland, M.; Nessen, M.A.; Sterk, S. Current trends in mass spectrometry of peptides and proteins: Application to veterinary and sports-doping control. Mass Spectrom. Rev. 2015, 34, 571–594. [Google Scholar] [CrossRef]
- Katsila, T.; Siskos, A.P.; Tamvakopoulos, C. Peptide and protein drugs: The study of their metabolism and catabolism by mass spectrometry. Mass Spectrom. Rev. 2012, 31, 110–133. [Google Scholar] [CrossRef]
- Esposito, S.; Deventer, K.; Geldof, L.; Van Eenoo, P. In vitro models for metabolic studies of small peptide hormones in sport drug testing. J. Pept. Sci. 2015, 21, 1–9. [Google Scholar] [CrossRef]
- Okano, M.; Sato, M.; Ikekita, A.; Kageyama, S. Determination of growth hormone secretagogue pralmorelin (ghrp-2) and its metabolite in human urine by liquid chromatography/electrospray ionization tandem mass spectrometry. Rapid Commun. Mass Spectrom. 2010, 24, 2046–2056. [Google Scholar] [CrossRef]
- Thomas, A.; Brinkkotter, P.; Schanzer, W.; Thevis, M. Metabolism of human insulin after subcutaneous administration: A possible means to uncover insulin misuse. Anal. Chim. Acta 2015, 897, 53–61. [Google Scholar] [CrossRef]
- Thomas, A.; Delahaut, P.; Krug, O.; Schanzer, W.; Thevis, M. Metabolism of growth hormone releasing peptides. Anal. Chem. 2012, 84, 10252–10259. [Google Scholar] [CrossRef]
- Thomas, A.; Knoop, A.; Schanzer, W.; Thevis, M. Characterization of in vitro generated metabolites of selected peptides <2 kda prohibited in sports. Drug Test Anal. 2017, 9, 1799–1803. [Google Scholar] [CrossRef] [Green Version]
- Thomas, A.; Thevis, M. Identification of metabolites of peptide-derived drugs using an isotope-labeled reporter ion screening strategy. Clin. Chem. Lab. Med. 2020, 58, 690–700. [Google Scholar] [CrossRef]
- Zvereva, I.; Semenistaya, E.; Krotov, G.; Rodchenkov, G. Comparison of various in vitro model systems of the metabolism of synthetic doping peptides: Proteolytic enzymes, human blood serum, liver and kidney microsomes and liver s9 fraction. J. Proteomics 2016, 149, 85–97. [Google Scholar] [CrossRef]
- World Anti-Doping Agency. The 2021 Prohibited List. Available online: https://www.wada-ama.org/sites/default/files/resources/files/2021list_en.pdf (accessed on 27 January 2022).
- Barroso, O.; Mazzoni, I.; Rabin, O. Hormone abuse in sports: The antidoping perspective. Asian J. Androl. 2008, 10, 391–402. [Google Scholar] [CrossRef]
- Brennan, B.P.; Kanayama, G.; Hudson, J.I.; Pope, H.G., Jr. Human growth hormone abuse in male weightlifters. Am. J. Addict. 2011, 20, 9–13. [Google Scholar] [CrossRef] [Green Version]
- Holt, R.I.; Sonksen, P.H. Growth hormone, igf-i and insulin and their abuse in sport. Br. J. Pharmacol. 2008, 154, 542–556. [Google Scholar] [CrossRef] [Green Version]
- Holt, R.I.G.; Ho, K.K.Y. The use and abuse of growth hormone in sports. Endocr. Rev. 2019, 40, 1163–1185. [Google Scholar] [CrossRef]
- Hartman, M.L.; Faria, A.C.; Vance, M.L.; Johnson, M.L.; Thorner, M.O.; Veldhuis, J.D. Temporal structure of in vivo growth hormone secretory events in humans. Am. J. Physiol. 1991, 260, E101–E110. [Google Scholar] [CrossRef] [PubMed]
- Le Roith, D.; Bondy, C.; Yakar, S.; Liu, J.L.; Butler, A. The somatomedin hypothesis: 2001. Endocr. Rev. 2001, 22, 53–74. [Google Scholar] [CrossRef]
- Tanner, J.M. Human growth hormone. Nature 1972, 237, 433–439. [Google Scholar] [CrossRef] [PubMed]
- Anderson, L.J.; Tamayose, J.M.; Garcia, J.M. Use of growth hormone, igf-i, and insulin for anabolic purpose: Pharmacological basis, methods of detection, and adverse effects. Mol. Cell Endocrinol. 2018, 464, 65–74. [Google Scholar] [CrossRef]
- Bidlingmaier, M.; Suhr, J.; Ernst, A.; Wu, Z.; Keller, A.; Strasburger, C.J.; Bergmann, A. High-sensitivity chemiluminescence immunoassays for detection of growth hormone doping in sports. Clin. Chem. 2009, 55, 445–453. [Google Scholar] [CrossRef] [PubMed]
- Holt, R.I. Detecting growth hormone misuse in athletes. Indian J. Endocrinol. Metab. 2013, 17, S18–S22. [Google Scholar] [CrossRef] [PubMed]
- Powrie, J.K.; Bassett, E.E.; Rosen, T.; Jorgensen, J.O.; Napoli, R.; Sacca, L.; Christiansen, J.S.; Bengtsson, B.A.; Sonksen, P.H.; Group, G.H.P.S. Detection of growth hormone abuse in sport. Growth Horm. IGF Res. 2007, 17, 220–226. [Google Scholar] [CrossRef]
- Equey, T.; Pastor, A.; de la Torre Fornell, R.; Thomas, A.; Giraud, S.; Thevis, M.; Kuuranne, T.; Baume, N.; Barroso, O.; Aikin, R. Application of the athlete biological passport approach to the detection of growth hormone doping. J. Clin. Endocrinol. Metab. 2021. [Google Scholar] [CrossRef]
- The Mammalian Degradome Database. Available online: http://degradome.uniovi.es/dindex.html (accessed on 27 January 2022).
- de Veer, S.J.; Furio, L.; Harris, J.M.; Hovnanian, A. Proteases and proteomics: Cutting to the core of human skin pathologies. Proteomics Clin Appl 2014, 8, 389–402. [Google Scholar] [CrossRef]
- Falick, A.M.; Hines, W.M.; Medzihradszky, K.F.; Baldwin, M.A.; Gibson, B.W. Low-mass ions produced from peptides by high-energy collision-induced dissociation in tandem mass spectrometry. J. Am. Soc. Mass Spectrom. 1993, 4, 882–893. [Google Scholar] [CrossRef] [Green Version]
- Busby, W.H., Jr.; Quackenbush, G.E.; Humm, J.; Youngblood, W.W.; Kizer, J.S. An enzyme(s) that converts glutaminyl-peptides into pyroglutamyl-peptides. Presence in pituitary, brain, adrenal medulla, and lymphocytes. J. Biol. Chem. 1987, 262, 8532–8536. [Google Scholar] [CrossRef]
- Tritsch, G.L.; Moore, G.E. Spontaneous decomposition of glutamine in cell culture media. Exp. Cell Res. 1962, 28, 360–364. [Google Scholar] [CrossRef]
- Pfizer Pharma GmbH. Genotropin 5mg/ml Full Prescribing Information. Available online: https://www.medicines.org.uk/emc/product/32/smpc#gref (accessed on 2 February 2022).
- Ferring GmbH. Zomacton 10 mg/ml Full Prescribing Information. Available online: https://www.hpra.ie/img/uploaded/swedocuments/Licence_PA1009-008-003_13102021160350.pdf (accessed on 2 February 2022).
- Merck Serono GmbH. Saizen 5,83 mg/ml Full Prescribing Information. Available online: https://medinfo.emdserono.ca/content/dam/web/health-care/biopharma/web/MI/en_CA/docs/Saizen-PP-ENG-Aug-7.pdf (accessed on 2 February 2022).
- Allard, J.; Duan, C. Igf-binding proteins: Why do they exist and why are there so many? Front. Endocinol. 2018, 9, 117. [Google Scholar] [CrossRef] [Green Version]
- Clemmons, D.R. Role of insulin-like growth factor binding proteins in controlling igf actions. Mol. Cell Endocrinol. 1998, 140, 19–24. [Google Scholar] [CrossRef]
- Clemmons, D.R. Igf-i assays: Current assay methodologies and their limitations. Pituitary 2007, 10, 121–128. [Google Scholar] [CrossRef]
- Thomas, A.; Kohler, M.; Schanzer, W.; Delahaut, P.; Thevis, M. Determination of igf-1 and igf-2, their degradation products and synthetic analogues in urine by lc-ms/ms. Analyst 2011, 136, 1003–1012. [Google Scholar] [CrossRef]
- Yamamoto, H.; Murphy, L.J. Enzymatic conversion of igf-i to des(1-3)igf-i in rat serum and tissues: A further potential site of growth hormone regulation of igf-i action. J. Endocrinol. 1995, 146, 141–148. [Google Scholar] [CrossRef]
- Ipsen Biopharmaceuticals, I. Increlex Full Prescribing Information. Available online: https://www.ipsen.com/websites/Ipsen_Online/wp-content/uploads/sites/9/2020/04/10140519/Increlex-Full-Prescribing-Information (accessed on 2 February 2022).




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peptide | Amino Acid Sequence | Monoisotopic Mass (Da) | Dominant Charge State | Retention Time (min) | Product of Proteolytic System | |||
|---|---|---|---|---|---|---|---|---|
| Skin S9 | Liver S9 | Serum | Urine | |||||
| Intact | - | 1106.5593 | 20+ | 9.21 | + | + | + | + |
| 135–142 | TGQIFKQT | 922.4993 | 1+ | 4.96 | + | |||
| 135–143 | TGQIFKQTY | 1085.5626 | 1+ | 5.48 | + | + | ||
| 34–44 | AYIPKEQKYSF | 687.3586 | 2+ | 5.52 | + | + | ||
| 125–139 | MGRLEDGSPRTGQIF | 832.4147 | 2+ | 5.86 | ||||
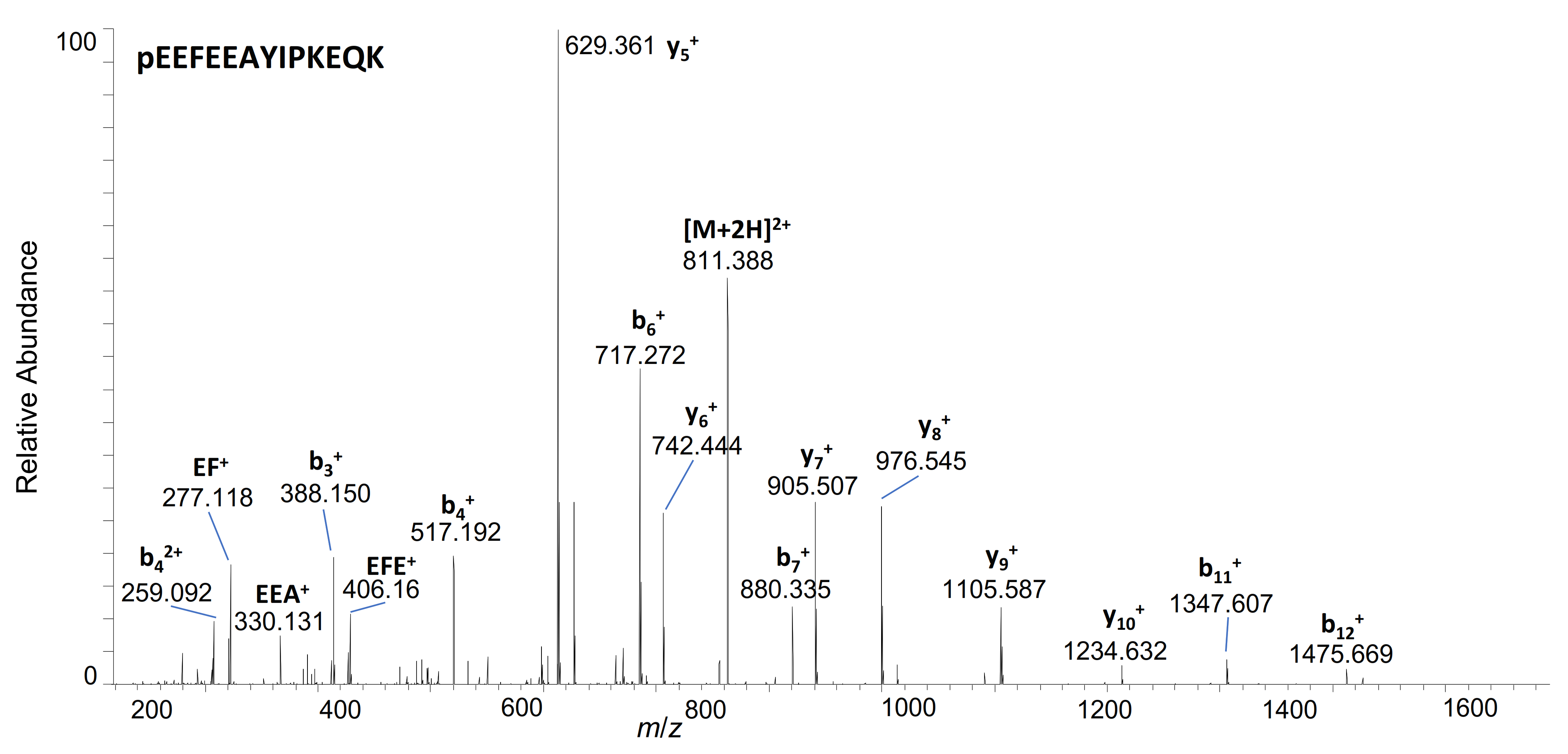
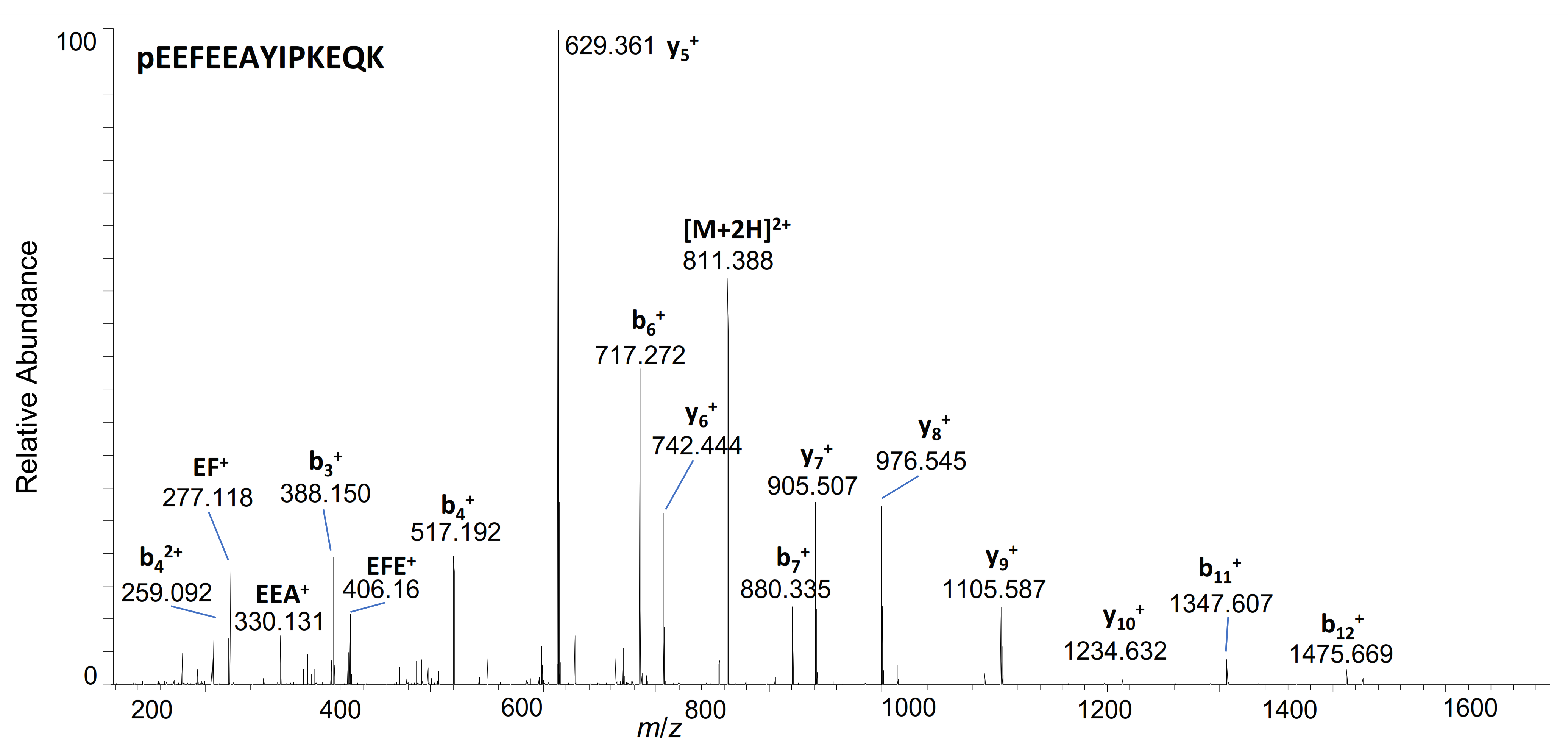
| 29–41 | pE * EFEEAYIPKEQK | 811.3909 | 2+ | 5.89 | + | |||
| 9–16 | LFDNAMLR | 979.5030 | 1+ | 6.21 | + | |||
| 86–91 | WLEPVQ | 771.4036 | 1+ | 6.25 | + | + | + | |
| 102–113 | VYGASDSNVYDL | 1302.5848 | 1+ | 5.32 | + | |||
| 114–124 | LKDLEEGIQTL | 629.8481 | 2+ | 6.36 | + | |||
| 29–38 | pE*EFEEAYIPK | 1236.5783 | 1+ | 6.41 | + | + | ||
| 86–92 | WLEPVQF | 918.4720 | 1+ | 7.45 | + | + | ||
| 1–13 | FPTIPLSRLFDNA | 745.9037 | 2+ | 7.80 | + | + | ||
| 1–16 | FPTIPLSRLFDNAMLR | 631.0135 | 3+ | 8.20 | + | + | ||
| Peptide | Amino Acid Sequence | Monoisotopic Mass (Da) | Dominant Charge State | Retention Time (min) | Product of Proteolytic System | |||
|---|---|---|---|---|---|---|---|---|
| Skin S9 | Liver S9 | Serum | Urine | |||||
| Intact | 956.9592 | 8+ | 6.89 | + | + | + | ||
| 25–36 | FNKPTGYGSSSR | 650.8177 | 2+ | 4.36 | + | + | + | |
| 25–35 | FNKPTGYGSSS | 572.7671 | 2+ | 4.60 | + | + | + | |
| 24–31 | YFNKPTGY | 989.4727 | 1+ | 5.18 | + | |||
| 22–36 | GFYFNKPTGYGSSSR | 834.3943 | 2+ | 5.55 | + | + | + | |
| 4–70 | TLCGAELVDALQFVCGDRGFYFNKPTGYGSSSRRAPQTGIVDECCFRSCDLRRLEMYCAPLKPAKSA | 921.0660 | 8+ | 6.85 | + | + | + | + |
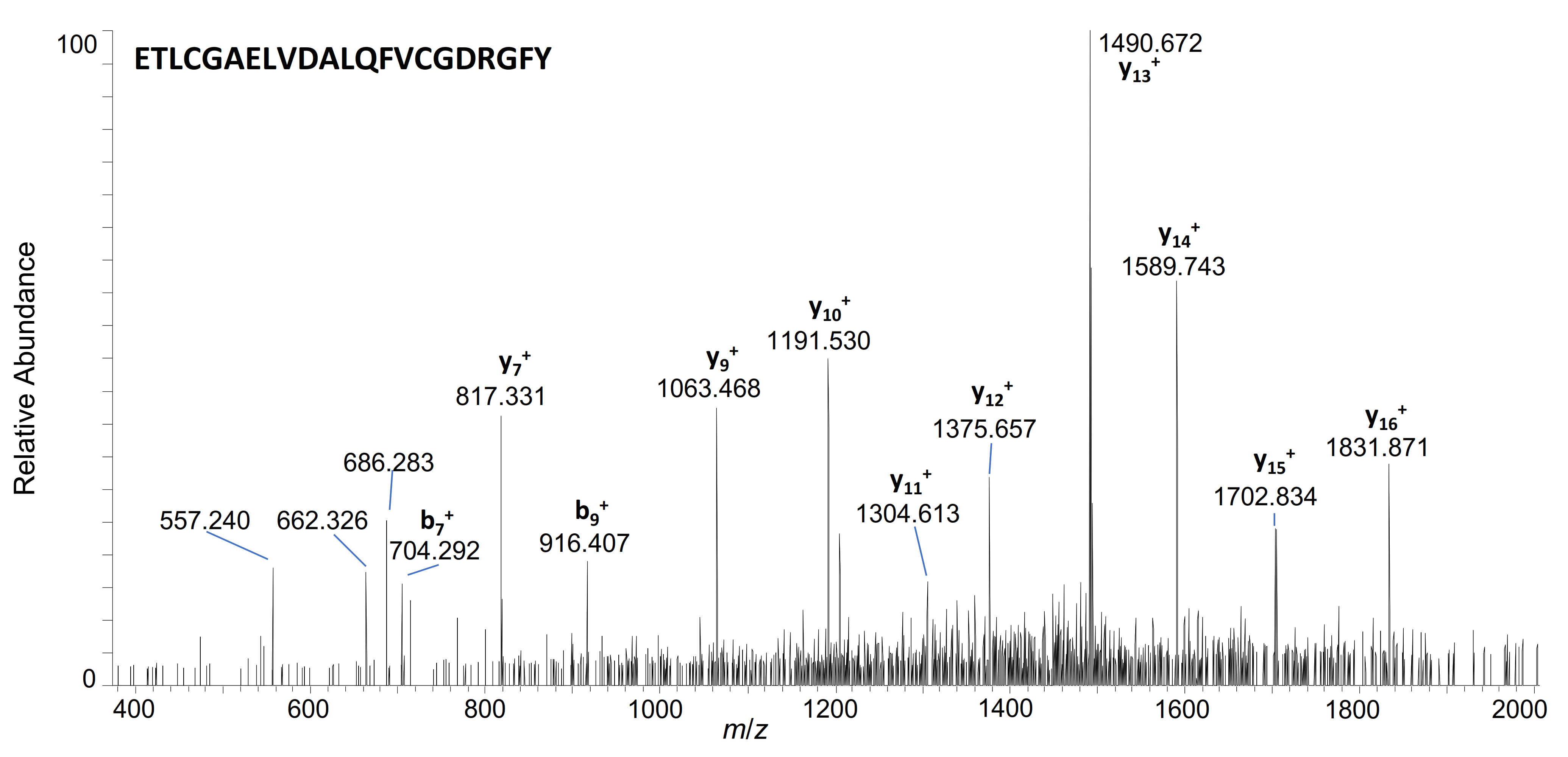
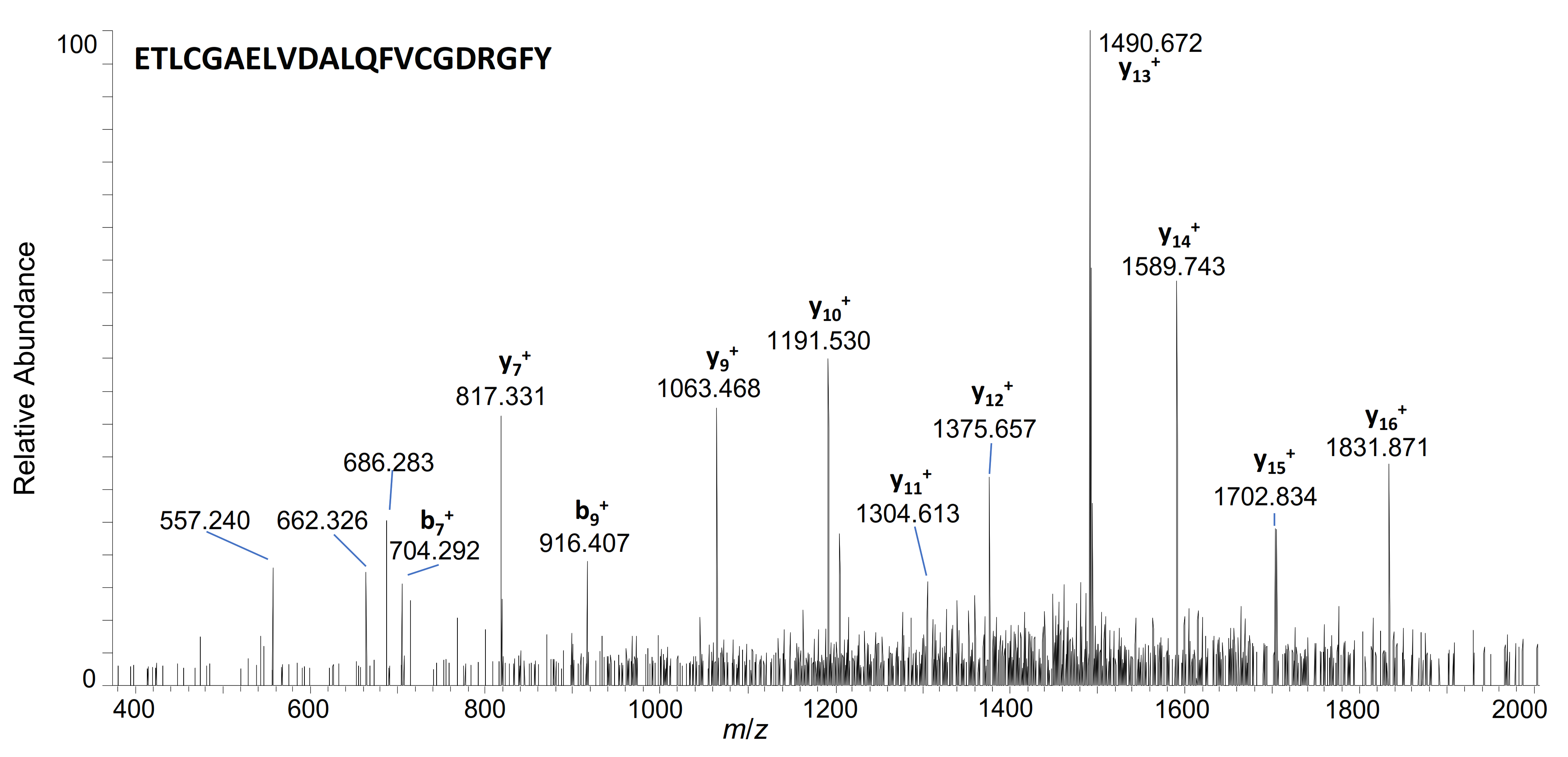
| 3–36 | ETLCGAELVDALQFVCGDRGFYFNKPTGYGSSSR | 922.1817 | 4+ | 7.48 | + | + | ||
| 3–35 | ETLCGAELVDALQFVCGDRGFYFNKPTGYGSSS | 1177.2061 | 3+ | 7.72 | + | + | + | |
| 3–31 | ETLCGAELVDALQFVCGDRGFYFNKPTGY | 1071.167 | 3+ | 7.83 | + | + | + | |
| 3–24 | ETLCGAELVDALQFVCGDRGFY | 1202.5510 | 2+ | 8.15 | + | + | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Krombholz, S.; Thomas, A.; Thevis, M. Investigations into the In Vitro Metabolism of hGH and IGF-I Employing Stable-Isotope-Labelled Drugs and Monitoring Diagnostic Immonium Ions by High-Resolution/High-Accuracy Mass Spectrometry. Metabolites 2022, 12, 146. https://doi.org/10.3390/metabo12020146
Krombholz S, Thomas A, Thevis M. Investigations into the In Vitro Metabolism of hGH and IGF-I Employing Stable-Isotope-Labelled Drugs and Monitoring Diagnostic Immonium Ions by High-Resolution/High-Accuracy Mass Spectrometry. Metabolites. 2022; 12(2):146. https://doi.org/10.3390/metabo12020146
Chicago/Turabian StyleKrombholz, Sophia, Andreas Thomas, and Mario Thevis. 2022. "Investigations into the In Vitro Metabolism of hGH and IGF-I Employing Stable-Isotope-Labelled Drugs and Monitoring Diagnostic Immonium Ions by High-Resolution/High-Accuracy Mass Spectrometry" Metabolites 12, no. 2: 146. https://doi.org/10.3390/metabo12020146
APA StyleKrombholz, S., Thomas, A., & Thevis, M. (2022). Investigations into the In Vitro Metabolism of hGH and IGF-I Employing Stable-Isotope-Labelled Drugs and Monitoring Diagnostic Immonium Ions by High-Resolution/High-Accuracy Mass Spectrometry. Metabolites, 12(2), 146. https://doi.org/10.3390/metabo12020146