
A Comparison of Mother’s Milk and the Neonatal Urine Metabolome: A Unique Fingerprinting for Different Nutritional Phenotypes


 ,
,  , ,
, , 
Abstract
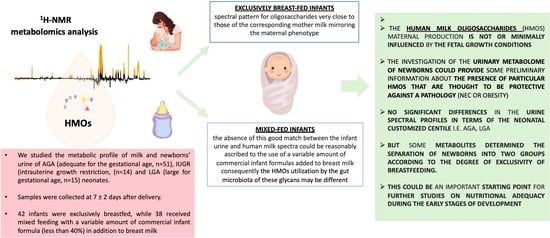
:
1. Introduction
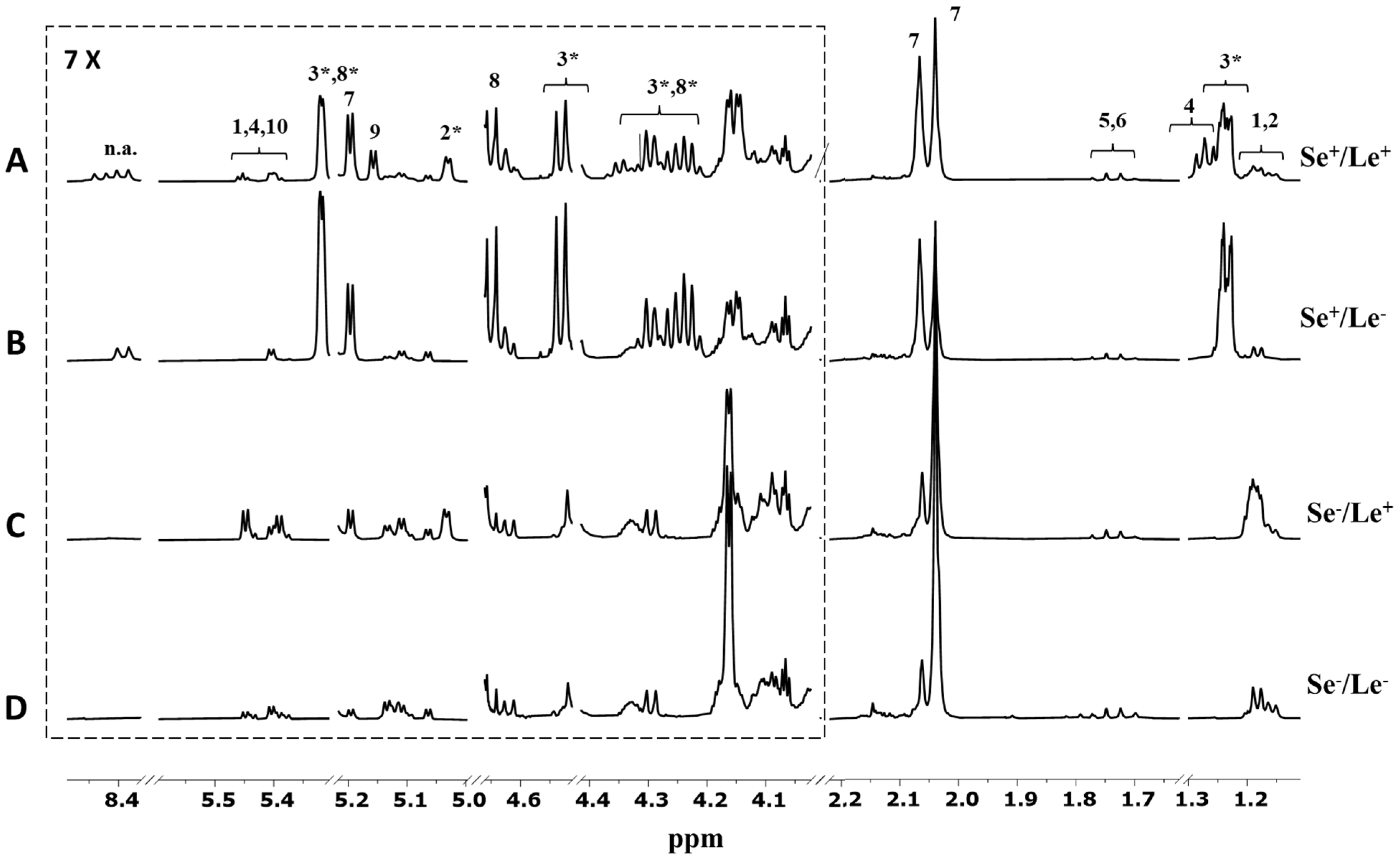
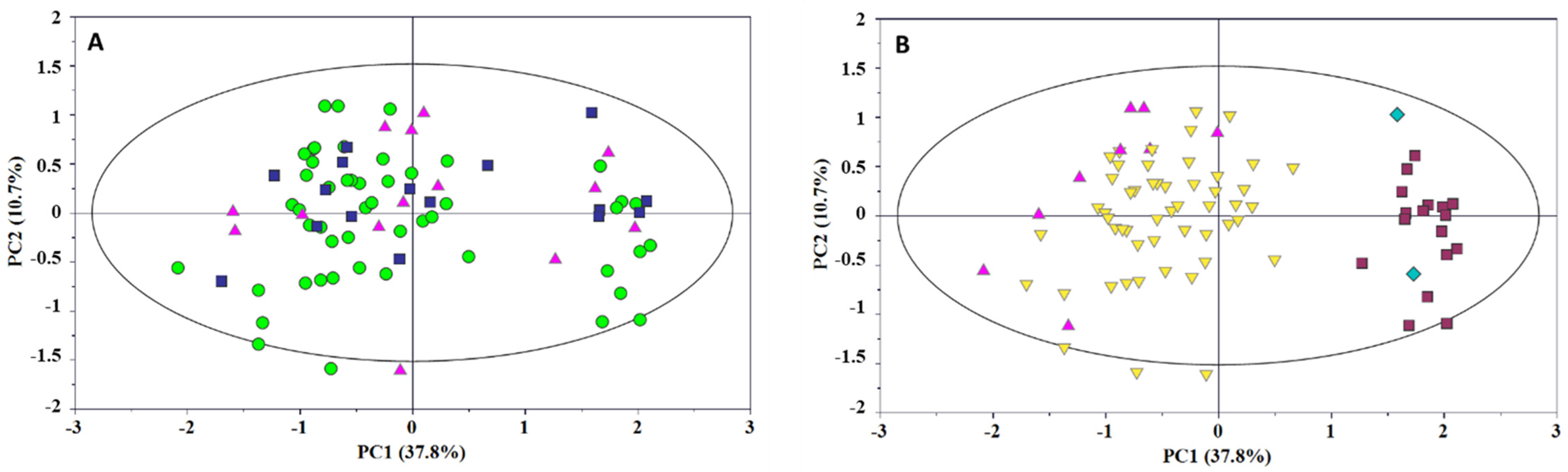
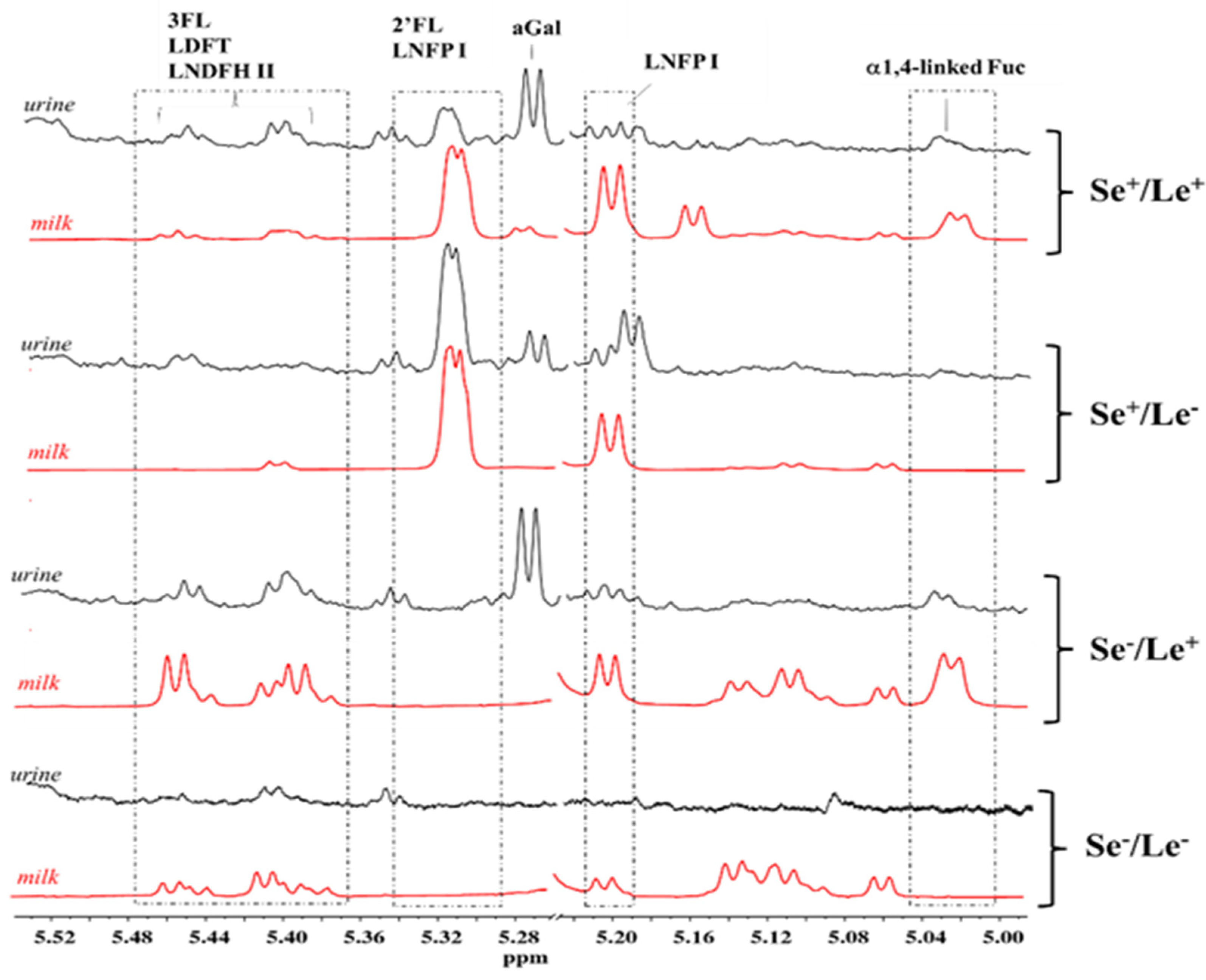
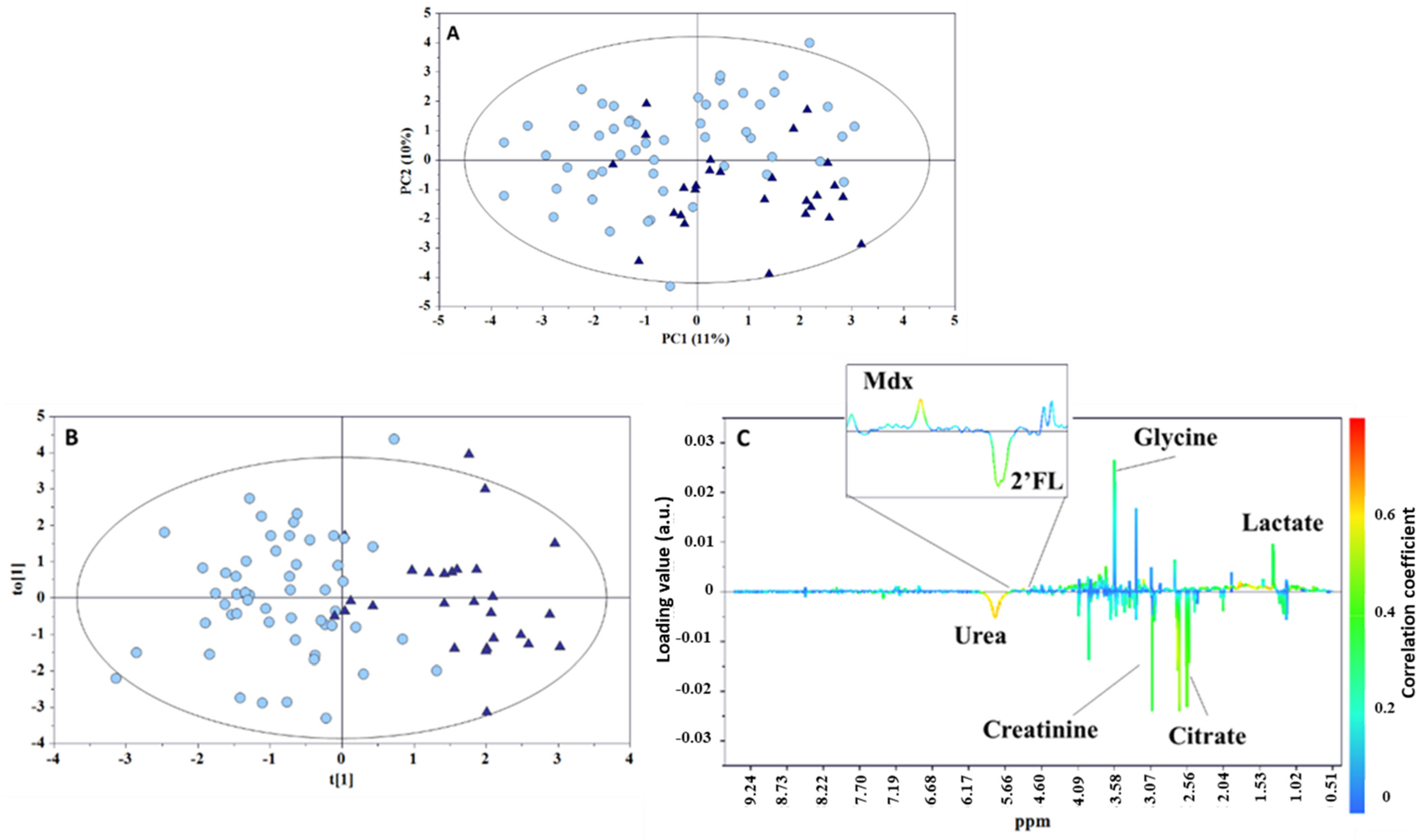
2. Results and Discussion

3. Materials and Methods
3.1. Subject and Sample Collection
3.2. Sample Preparation
3.3. NMR Spectroscopy
3.4. Spectral Processing and Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ballard, O.; Morrow, A.L. Human milk composition: Nutrients and bioactive factors. Pediatr. Clin. N. Am. 2013, 60, 49–74. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Corona, L.; Lussu, A.; Bosco, A.; Pintus, R.; Cesare Marincola, F.; Fanos, V.; Dessì, A. Human Milk Oligosaccharides: A Comprehensive Review towards Metabolomics. Children 2021, 8, 804. [Google Scholar] [CrossRef] [PubMed]
- Seferovic, M.D.; Mohammad, M.; Pace, R.M.; Engevik, M.; Versalovic, J.; Bode, L.; Haymond, M.; Aagaard, K.M. Maternal diet alters human milk oligosaccharide composition with implications for the milk metagenome. Sci. Rep. 2020, 10, 22092. [Google Scholar] [CrossRef] [PubMed]
- Biddulph, C.; Holmes, M.; Kuballa, A.; Davies, P.S.W.; Koorts, P.; Carter, R.J.; Maher, J. Human Milk Oligosaccharide Profiles and Associations with Maternal Nutritional Factors: A Scoping Review. Nutrients 2021, 13, 965. [Google Scholar] [CrossRef] [PubMed]
- Saben, J.L.; Sims, C.R.; Abraham, A.; Bode, L.; Andres, A. Human Milk Oligosaccharide Concentrations and Infant Intakes Are Associated with Maternal Overweight and Obesity and Predict Infant Growth. Nutrients 2021, 13, 446. [Google Scholar] [CrossRef] [PubMed]
- Gómez-Gallego, C.; Morales, J.; Monleón, D.; du Toit, E.; Kumar, H.; Linderborg, K.; Zhang, Y.; Yang, B.; Isolauri, E.; Salminen, S.; et al. Human Breast Milk NMR Metabolomic Profile across Specific Geographical Locations and Its Association with the Milk Microbiota. Nutrients 2018, 10, 1355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Azad, M.B.; Robertson, B.; Atakora, F.; Becker, A.B.; Subbarao, P.; Moraes, T.J.; Mandhane, P.J.; Turvey, S.E.; Lefebvre, D.L.; Sears, M.R.; et al. Human Milk Oligosaccharide Concentrations Are Associated with Multiple Fixed and Modifiable Maternal Characteristics, Environmental Factors, and Feeding Practices. J. Nutr. 2018, 148, 1733–1742. [Google Scholar] [CrossRef] [PubMed]
- Fanos, V.; Pintus, R.; Dessì, A. Clinical Metabolomics in Neonatology: From Metabolites to Diseases. Neonatology 2018, 113, 406–413. [Google Scholar] [CrossRef]
- Dessì, A.; Murgia, A.; Agostino, R.; Pattumelli, M.G.; Schirru, A.; Scano, P.; Fanos, V.; Caboni, P. Exploring the Role of Different Neonatal Nutrition Regimens during the First Week of Life by Urinary GC-MS Metabolomics. Int. J. Mol. Sci. 2016, 17, 265. [Google Scholar] [CrossRef] [Green Version]
- Dessì, A.; Puddu, M.; Ottonello, G.; Fanos, V. Metabolomics and fetal-neonatal nutrition: Between "not enough" and "too much". Molecules 2013, 18, 11724–11732. [Google Scholar] [CrossRef] [Green Version]
- Van Leeuwen, S.S.; Schoemaker, R.J.W.; Gerwig, G.J.; van Leusen-van Kan, E.J.M.; Dijkhuizen, L.; Kamerling, J.P. Rapid milk group classification by 1H NMR analysis of Le and H epitopes in human milk oligosaccharide donor samples. Glicobyology 2014, 24, 728–739. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Praticò, G.; Capuani, G.; Tommassini, A.; Baldassarre, M.E.; Delfini, M.; Miccheli, A. Exploring human breast milk composition by NMR-based metabolomics. Nat. Prod. Res. 2014, 28, 95–101. [Google Scholar] [CrossRef] [PubMed]
- Wang, A.; Koleva, P.; du Toit, E.; Geddes, D.T.; Munblit, D.; Prescott, S.L.; Eggesbø, M.; Johnson, C.C.; Wegienka, G.; Shimojo, N.; et al. The Milk Metabolome of Non-secretor and Lewis Negative Mothers. Front. Nutr. 2021, 7, 380. [Google Scholar] [CrossRef] [PubMed]
- Conta, G.; Del Chierico, F.; Reddel, S.; Marini, F.; Sciubba, F.; Capuani, G.; Tomassini, A.; Di Cocco, M.E.; Laforgia, N.; Baldassarre, M.E.; et al. Longitudinal Multi-Omics Study of a Mother-Infant Dyad from Breastfeeding to Weaning: An Individualized Approach to Understand the Interactions Among Diet, Fecal Metabolome and Microbiota Composition. Front. Mol. Biosci. 2021, 8, 688440. [Google Scholar] [CrossRef] [PubMed]
- Smilowitz, J.T.; O’Sullivan, A.; Barile, D.; German, J.B.; Lönnerdal, B.; Slupski, C.M. The human milk metabolome reveals diverse oligosaccharides profiles. J. Nutr. 2013, 143, 1709–1718. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spevacek, A.R.; Smilowitz, J.T.; Chin, E.L.; Underwood, M.A.; German, J.B.; Slupsky, C.M. Infant Maturity at Birth Reveals Minor Differences in the Maternal Milk Metabolome in the First Month of Lactation. J. Nutr. 2015, 145, 1698–1708. [Google Scholar] [CrossRef] [Green Version]
- Sundekilde, U.K.; Downey, E.; O’Mahony, J.A.; O’Shea, C.A.; Ryan, C.A.; Kelly, A.L.; Bertram, H.C. The Effect of Gestational and Lactational Age on the Human Milk Metabolome. Nutrients 2016, 8, 304. [Google Scholar] [CrossRef] [Green Version]
- Dessì, A.; Briana, D.; Corbu, S.; Gavrili, S.; Cesare Marincola, F.; Georgantzi, S.; Pintus, R.; Fanos, V.; Malamitsi-Puchner, A. Metabolomics of breast milk: The importance of phenotypes. Metabolites 2018, 8, 79. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cesare Marincola, F.; Corbu, S.; Lussu, M.; Noto, A.; Dessì, A.; Longo, S.; Civardi, E.; Garofoli, F.; Grenci, B.; Mongini, E.; et al. Impact of early postnatal nutrition on the NMR urinary metabolic profile of infant. J. Proteome Res. 2016, 15, 3712–3723. [Google Scholar] [CrossRef]
- Chiu, C.Y.; Yeh, K.W.; Lin, G.; Chiang, M.H.; Yang, S.C.; Chao, W.J.; Yao, T.C.; Tsai, M.H.; Hua, M.C.; Liao, S.L.; et al. Metabolomics Reveals Dynamic Metabolic Changes Associated with Age in Early Childhood. PLoS ONE 2016, 11, e0149823. [Google Scholar] [CrossRef] [Green Version]
- Pottel, H.; Vrydags, N.; Mahieu, B.; Vandewynckele, E.; Croes, K.; Martens, F. Establishing age/sex related serum creatinine reference intervals from hospital laboratory data based on different statistical methods. Clin. Chim. Acta 2008, 396, 49–55. [Google Scholar] [CrossRef] [PubMed]
- Boer, D.P.; de Rijke, Y.B.; Hop, W.C.; Cransberg, K.; Dorresteijn, E.M. Reference valuesfor serum creatinine in children younger than 1 year of age. Pediatr. Nephrol. 2010, 25, 2107–2113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- López-Hernández, Y.; Oropeza-Valdez, J.J.; Blanco-Sandate, J.O.; Herrera-Van Oostdam, A.S.; Zheng, J.; Chi Guo, A.; Lima-Rogel, V.; Rajabzadeh, R.; Salgado-Bustamante, M.; Adrian-Lopez, J.; et al. The Urinary Metabolome of Healthy Newborns. Metabolites 2020, 10, 165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marincola, F.C.; Dessì, A.; Pattumelli, M.G.; Corbu, S.; Ossicini, C.; Ciccarelli, S.; Agostino, R.; Mussap, M.; Fanos, V. (1)H NMR-based urine metabolic profile of IUGR, LGA, and AGA newborns in the first week of life. Clin. Chim. Acta 2015, 451 Pt A, 28–34. [Google Scholar] [CrossRef]
- Gale, C.; Logan, K.M.; Santhakumaran, S.; Parkinson, J.R.; Hyde, M.J.; Modi, N. Effect of breastfeeding compared with formula feeding on infant body composition: A systematic review and meta-analysis. Am. J. Clin. Nutr. 2012, 95, 656–669. [Google Scholar] [CrossRef] [Green Version]
- Breij, L.M.; Abrahamse-Berkeveld, M.; Acton, D.; De Lucia Rolfe, E.; Ong, K.K.; Hokken-Koelega, A.C.S. Impact of Early Infant Growth, Duration of Breastfeeding and Maternal Factors on Total Body Fat Mass and Visceral Fat at 3 and 6 Months of Age. Ann. Nutr. Metab. 2017, 71, 203–210. [Google Scholar] [CrossRef]
- Koletzko, B.; von Kries, R.; Closa, R.; Escribano, J.; Scaglioni, S.; Giovannini, M.; Beyer, J.; Demmelmair, H.; Anton, B.; Gruszfeld, D.; et al. Can infant feeding choices modulate later obesity risk? Am. J. Clin. Nutr. 2009, 89, 1502S–1508S. [Google Scholar] [CrossRef] [Green Version]
- Shoji, H.; Taka, H.; Kaga, N.; Ikeda, N.; Kitamura, T.; Miura, Y.; Shimizu, T. A pilot study of the effect of human breast milk on urinary metabolome analysis in infants. J. Pediatr. Endocrinol. Metab. 2017, 30, 939–946. [Google Scholar] [CrossRef]
- Lucas, A.; Boyes, S.; Bloom, S.R.; Aynsley-Green, A. Metabolic and endocrine responses to a milk feed in six-day-old term infants: Differences between breast and cow’s milk formula feeding. Acta Paediatr. Scand. 1981, 70, 195–200. [Google Scholar] [CrossRef]
- Guay, C.; Madiraju, S.R.; Aumais, A.; Joly, E.; Prentki, M. A role for ATP-citrate lyase, malic enzyme, and pyruvate/citrate cycling in glucose-induced insulin secretion. J. Biol. Chem. 2007, 282, 35657–35665. [Google Scholar] [CrossRef] [Green Version]
- Gu, H.; Pan, Z.; Xi, B.; Hainline, B.E.; Shanaiah, N.; Asiago, V.; Gowda, G.A.; Raftery, D. 1H NMR metabolomics study of age profiling in children. NMR Biomed. 2009, 22, 826–833. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guneral, F.; Bachmann, C. Age-related reference values for urinary organic acids in a healthy Turkish pediatric population. Clin. Chem. 1994, 40, 862–866. [Google Scholar] [CrossRef] [PubMed]
- Kirejczyk, J.K.; Porowski, T.; Konstantynowicz, J.; Kozerska, A.; Nazarkiewicz, A.; Hoppe, B.; Wasilewska, A. Urinary citrate excretion in healthy children depends on age and gender. Pediatr. Nephrol. 2014, 29, 1575–1582. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bahado-Singh, R.O.; Yilmaz, A.; Bisgin, H.; Turkoglu, O.; Kumar, P.; Sherman, E.; Mrazik, A.; Odibo, A.; Graham, S.F. Artificial intelligence and the analysis of multi-platform metabolomics data for the detection of intrauterine growth restriction. PLoS ONE 2019, 14, e0214121. [Google Scholar] [CrossRef]
- Gregersen, N.; Ingerslev, J.; Rasmussen, K. Low molecular weight organic acids in the urine of the newborn. Acta Paediatr. Scand. 1977, 66, 85–89. [Google Scholar] [CrossRef]
- Neuberger, A. The metabolism of glycine and serine. Comp. Biochem. 1981, 19, 257–303. [Google Scholar]
- Persaud, C.; Forrester, T.; Jackson, A.A. Urinary excretion of 5-L-oxoproline (pyroglutamic acid) is increased during recovery from severe childhood malnutrition and responds to supplemental glycine. J. Nutr. 1996, 126, 2823–2830. [Google Scholar]
- Mabrouk, G.M.; Jois, M.; Brosnan, J.T. Cell signalling and the hormonal stimulation of the hepatic glycine cleavage enzyme system by glucagon. Biochem. J. 1998, 330, 759–763. [Google Scholar] [CrossRef] [Green Version]
- He, X.; Parenti, M.; Grip, T.; Lönnerdal, B.; Timby, N.; Domellöf, M.; Hernell, O.; Slupsky, C.M. Fecal microbiome and metabolome of infants fed bovine MFGM supplemented formula or standard formula with breast-fed infants as reference: A randomized controlled trial. Sci. Rep. 2019, 9, 11589. [Google Scholar] [CrossRef] [Green Version]
- Forbes, J.D.; Azad, M.B.; Vehling, L.; Tun, H.M.; Konya, T.B.; Guttman, D.S.; Field, C.J.; Lefebvre, D.; Sears, M.R.; Canadian Healthy Infant Longitudinal Development (CHILD) Study Investigators; et al. Association of Exposure to Formula in the Hospital and Subsequent Infant Feeding Practices With Gut Microbiota and Risk of Overweight in the First Year of Life. JAMA Pediatr. 2018, 172, e181161. [Google Scholar] [CrossRef]
- Sodhi, C.P.; Wipf, P.; Yamaguchi, Y.; Fulton, W.B.; Kovler, M.; Niño, D.F.; Zhou, Q.; Banfield, E.; Werts, A.D.; Ladd, M.R.; et al. The human milk oligosaccharides 2’-fucosyllactose and 6’-sialyllactose protect against the development of necrotizing enterocolitis by inhibiting toll-like receptor 4 signaling. Pediatr Res. 2021, 89, 91–101. [Google Scholar] [CrossRef] [PubMed]
- Alderete, T.L.; Autran, C.; Brekke, B.E.; Knight, R.; Bode, L.; Goran, M.I.; Fields, D.A. Associations between human milk oligosaccharides and infant body composition in the first 6 mo of life. Am. J. Clin. Nutr. 2015, 102, 1381–1388. [Google Scholar] [CrossRef] [PubMed]
- Dessì, A.; Marincola, F.C.; Masili, A.; Gazzolo, D.; Fanos, V. Clinical Metabolomics and Nutrition: The New Frontier in Neonatology and Pediatrics. Biomed. Res. Int. 2014, 2014, 981219. [Google Scholar] [CrossRef] [PubMed]
- Eriksson, L.; Trygg, J.; Wold, S. CV-ANOVA for significance testing of PLS and OPLS® models. J. Chemom. Soc. 2008, 22, 594–600. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| IUGR (n = 14) | AGA (n = 51) | LGA (n = 15) | |
|---|---|---|---|
| Maternal Characteristics | |||
| Age. years: (min; max) | 34.71 (22.00; 44.00) | 35.25 (24.00; 43.00) | 34.13 (26.00; 44.00) |
| Type of delivery (vaginal delivery/cesarean section) | 6/8 | 26/25 | 5/10 |
| Amniocentesis | 3 | 14 | 6 |
| Chorionic villus sampling | 2 | 1 | 0 |
| Infants characteristics at birth | |||
| Gender (male/female) | 8/6 | 28/23 | 8/7 |
| Gestational age, weeks (mean ± SD) | 38 ± 4 | 39 ± 3 | 39 ± 2 |
| Birth weight, g (min; max) | 2412 (1450; 2790) | 3236 (2.500; 4110) | 3.975 (3220; 4490) |
| Length at birth, cm (min; max) | 46.5 (30; 51) | 50.1 (34.; 55) | 50.3 (36; 57) |
| Head circumference, cm (min; max) | 33.4 (32; 41) | 35.7 (33; 51) | 38.6 (34; 55) |
| Percentile | 3.57° | 46° | 94.87° |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dessì, A.; Marzullo, A.; Corbu, S.; Bosco, A.; Cesare Marincola, F.; Pattumelli, M.G.; Mussap, M.; Pintus, R.; Fanos, V.; Agostino, R. A Comparison of Mother’s Milk and the Neonatal Urine Metabolome: A Unique Fingerprinting for Different Nutritional Phenotypes. Metabolites 2022, 12, 113. https://doi.org/10.3390/metabo12020113
Dessì A, Marzullo A, Corbu S, Bosco A, Cesare Marincola F, Pattumelli MG, Mussap M, Pintus R, Fanos V, Agostino R. A Comparison of Mother’s Milk and the Neonatal Urine Metabolome: A Unique Fingerprinting for Different Nutritional Phenotypes. Metabolites. 2022; 12(2):113. https://doi.org/10.3390/metabo12020113
Chicago/Turabian StyleDessì, Angelica, Alessandra Marzullo, Sara Corbu, Alice Bosco, Flaminia Cesare Marincola, Maria Grazia Pattumelli, Michele Mussap, Roberta Pintus, Vassilios Fanos, and Rocco Agostino. 2022. "A Comparison of Mother’s Milk and the Neonatal Urine Metabolome: A Unique Fingerprinting for Different Nutritional Phenotypes" Metabolites 12, no. 2: 113. https://doi.org/10.3390/metabo12020113
APA StyleDessì, A., Marzullo, A., Corbu, S., Bosco, A., Cesare Marincola, F., Pattumelli, M. G., Mussap, M., Pintus, R., Fanos, V., & Agostino, R. (2022). A Comparison of Mother’s Milk and the Neonatal Urine Metabolome: A Unique Fingerprinting for Different Nutritional Phenotypes. Metabolites, 12(2), 113. https://doi.org/10.3390/metabo12020113