
Sex-Specific Catabolic Metabolism Alterations in the Critically Ill following High Dose Vitamin D


 , and
, and 
Abstract
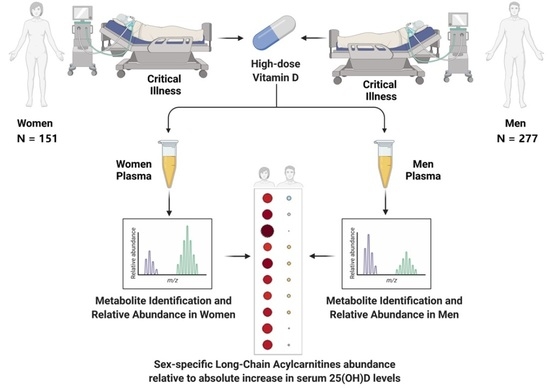
1. Introduction
2. Results
2.1. Demographics
2.2. Pharmacokinetics
2.3. Sex-Stratified Analyses
2.4. Responder Cohort Analysis
2.5. Mediation Analysis
3. Discussion
4. Materials and Methods
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gandhi, M.; Aweeka, F.; Greenblatt, R.M.; Blaschke, T.F. Sex differences in pharmacokinetics and pharmacodynamics. Annu. Rev. Pharm. Toxicol. 2004, 44, 499–523. [Google Scholar] [CrossRef] [PubMed]
- Kolz, M.; Johnson, T.; Sanna, S.; Teumer, A.; Vitart, V.; Perola, M.; Mangino, M.; Albrecht, E.; Wallace, C.; Farrall, M.; et al. Meta-analysis of 28,141 individuals identifies common variants within five new loci that influence uric acid concentrations. PLoS Genet 2009, 5, e1000504. [Google Scholar] [CrossRef] [PubMed]
- Mittelstrass, K.; Ried, J.S.; Yu, Z.; Krumsiek, J.; Gieger, C.; Prehn, C.; Roemisch-Margl, W.; Polonikov, A.; Peters, A.; Theis, F.J.; et al. Discovery of sexual dimorphisms in metabolic and genetic biomarkers. PLoS Genet 2011, 7, e1002215. [Google Scholar] [CrossRef] [PubMed]
- Dunn, W.B.; Lin, W.; Broadhurst, D.; Begley, P.; Brown, M.; Zelena, E.; Vaughan, A.A.; Halsall, A.; Harding, N.; Knowles, J.D.; et al. Molecular phenotyping of a UK population: Defining the human serum metabolome. Metabolomics 2015, 11, 9–26. [Google Scholar] [CrossRef]
- Franconi, F.; Campesi, I. Sex Impact on Biomarkers, Pharmacokinetics and Pharmacodynamics. Curr. Med. Chem. 2017, 24, 2561–2575. [Google Scholar] [CrossRef]
- Kantae, V.; Krekels, E.H.J.; Esdonk, M.J.V.; Lindenburg, P.; Harms, A.C.; Knibbe, C.A.J.; Van der Graaf, P.H.; Hankemeier, T. Integration of pharmacometabolomics with pharmacokinetics and pharmacodynamics: Towards personalized drug therapy. Metabolomics 2017, 13, 9. [Google Scholar] [CrossRef]
- Karlsson Lind, L.; von Euler, M.; Korkmaz, S.; Schenck-Gustafsson, K. Sex differences in drugs: The development of a comprehensive knowledge base to improve gender awareness prescribing. Biol. Sex Differ. 2017, 8, 32. [Google Scholar] [CrossRef]
- Kiehntopf, M.; Nin, N.; Bauer, M. Metabolism, metabolome, and metabolomics in intensive care: Is it time to move beyond monitoring of glucose and lactate? Am. J. Respir. Crit. Care Med. 2013, 187, 906–907. [Google Scholar] [CrossRef]
- Johansson, P.I.; Nakahira, K.; Rogers, A.J.; McGeachie, M.J.; Baron, R.M.; Fredenburgh, L.E.; Harrington, J.; Choi, A.M.K.; Christopher, K.B. Plasma mitochondrial DNA and metabolomic alterations in severe critical illness. Crit. Care 2018, 22, 360. [Google Scholar] [CrossRef]
- Lasky-Su, J.; Dahlin, A.; Litonjua, A.A.; Rogers, A.J.; McGeachie, M.J.; Baron, R.M.; Gazourian, L.; Barragan-Bradford, D.; Fredenburgh, L.E.; Choi, A.M.K.; et al. Metabolome alterations in severe critical illness and vitamin D status. Crit. Care 2017, 21, 193. [Google Scholar] [CrossRef]
- Dupuis, M.L.; Pagano, M.T.; Pierdominici, M.; Ortona, E. The role of vitamin D in autoimmune diseases: Could sex make the difference? Biol. Sex Differ. 2021, 12, 12. [Google Scholar] [CrossRef] [PubMed]
- Sanghera, D.K.; Sapkota, B.R.; Aston, C.E.; Blackett, P.R. Vitamin D Status, Gender Differences, and Cardiometabolic Health Disparities. Ann. Nutr. Metab. 2017, 70, 79–87. [Google Scholar] [CrossRef] [PubMed]
- Ilahi, M.; Armas, L.A.; Heaney, R.P. Pharmacokinetics of a single, large dose of cholecalciferol. Am. J. Clin. Nutr. 2008, 87, 688–691. [Google Scholar] [CrossRef] [PubMed]
- Romagnoli, E.; Mascia, M.L.; Cipriani, C.; Fassino, V.; Mazzei, F.; D’Erasmo, E.; Carnevale, V.; Scillitani, A.; Minisola, S. Short and long-term variations in serum calciotropic hormones after a single very large dose of ergocalciferol (vitamin D2) or cholecalciferol (vitamin D3) in the elderly. J. Clin. Endocrinol. Metab. 2008, 93, 3015–3020. [Google Scholar] [CrossRef]
- Cipriani, C.; Romagnoli, E.; Scillitani, A.; Chiodini, I.; Clerico, R.; Carnevale, V.; Mascia, M.L.; Battista, C.; Viti, R.; Pileri, M.; et al. Effect of a single oral dose of 600,000 IU of cholecalciferol on serum calciotropic hormones in young subjects with vitamin D deficiency: A prospective intervention study. J. Clin. Endocrinol. Metab. 2010, 95, 4771–4777. [Google Scholar] [CrossRef]
- Sanders, K.M.; Stuart, A.L.; Williamson, E.J.; Simpson, J.A.; Kotowicz, M.A.; Young, D.; Nicholson, G.C. Annual high-dose oral vitamin D and falls and fractures in older women: A randomized controlled trial. JAMA 2010, 303, 1815–1822. [Google Scholar] [CrossRef]
- Kaddurah-Daouk, R.; Kristal, B.S.; Weinshilboum, R.M. Metabolomics: A global biochemical approach to drug response and disease. Annu. Rev. Pharmacol. Toxicol. 2008, 48, 653–683. [Google Scholar] [CrossRef]
- Amrein, K.; Lasky-Su, J.A.; Dobnig, H.; Christopher, K.B. Metabolomic basis for response to high dose vitamin D in critical illness. Clin. Nutr. 2021, 40, 2053–2060. [Google Scholar] [CrossRef]
- Soldin, O.P.; Mattison, D.R. Sex differences in pharmacokinetics and pharmacodynamics. Clin. Pharm. 2009, 48, 143–157. [Google Scholar] [CrossRef]
- Amrein, K.; Schnedl, C.; Holl, A.; Riedl, R.; Christopher, K.B.; Pachler, C.; Urbanic Purkart, T.; Waltensdorfer, A.; Munch, A.; Warnkross, H.; et al. Effect of high-dose vitamin D3 on hospital length of stay in critically ill patients with vitamin D deficiency: The VITdAL-ICU randomized clinical trial. JAMA 2014, 312, 1520–1530. [Google Scholar] [CrossRef]
- Fong, Y.M.; Marano, M.A.; Moldawer, L.L.; Wei, H.; Calvano, S.E.; Kenney, J.S.; Allison, A.C.; Cerami, A.; Shires, G.T.; Lowry, S.F. The acute splanchnic and peripheral tissue metabolic response to endotoxin in humans. J. Clin. Investg. 1990, 85, 1896–1904. [Google Scholar] [CrossRef]
- Boden, G. Obesity and free fatty acids. Endocrinol. Metab. Clin. N. Am. 2008, 37, 635–646. [Google Scholar] [CrossRef]
- Murphy, M.O.; Loria, A.S. Sex-specific effects of stress on metabolic and cardiovascular disease: Are women at higher risk? Am. J. Physiol. Regul. Integr. Comp. Physiol. 2017, 313, R1–R9. [Google Scholar] [CrossRef] [PubMed]
- Mauvais-Jarvis, F. Sex differences in metabolic homeostasis, diabetes, and obesity. Biol. Sex Differ. 2015, 6, 14. [Google Scholar] [CrossRef] [PubMed]
- Roeters van Lennep, J.E.; Westerveld, H.T.; Erkelens, D.W.; van der Wall, E.E. Risk factors for coronary heart disease: Implications of gender. Cardiovasc. Res. 2002, 53, 538–549. [Google Scholar] [CrossRef]
- Sugiyama, M.G.; Agellon, L.B. Sex differences in lipid metabolism and metabolic disease risk. Biochem. Cell Biol. 2012, 90, 124–141. [Google Scholar] [CrossRef]
- Uranga, A.P.; Levine, J.; Jensen, M. Isotope tracer measures of meal fatty acid metabolism: Reproducibility and effects of the menstrual cycle. Am. J. Physiol. Endocrinol. Metab. 2005, 288, E547–E555. [Google Scholar] [CrossRef]
- Preiser, J.C.; Ichai, C.; Orban, J.C.; Groeneveld, A.B. Metabolic response to the stress of critical illness. Br. J. Anaesth. 2014, 113, 945–954. [Google Scholar] [CrossRef] [PubMed]
- Chary, S.; Amrein, K.; Lasky-Su, J.; Dobnig, H.; Christopher, K.B. The Sex-specific Metabolic Response to Critical Illness: A post-hoc metabolomics study of the VITdAL-ICU trial. Sci. Rep. 2021, 11, 3951. [Google Scholar] [CrossRef]
- Blot, S.I.; Pea, F.; Lipman, J. The effect of pathophysiology on pharmacokinetics in the critically ill patient--concepts appraised by the example of antimicrobial agents. Adv. Drug Deliv. Rev. 2014, 77, 3–11. [Google Scholar] [CrossRef]
- Stern, J.H.; Rutkowski, J.M.; Scherer, P.E. Adiponectin, Leptin, and Fatty Acids in the Maintenance of Metabolic Homeostasis through Adipose Tissue Crosstalk. Cell Metab. 2016, 23, 770–784. [Google Scholar] [CrossRef] [PubMed]
- Laughlin, G.A.; Barrett-Connor, E.; May, S. Sex-specific determinants of serum adiponectin in older adults: The role of endogenous sex hormones. Int. J. Obes. 2007, 31, 457–465. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Kennedy, A.; Gettys, T.W.; Watson, P.; Wallace, P.; Ganaway, E.; Pan, Q.; Garvey, W.T. The metabolic significance of leptin in humans: Gender-based differences in relationship to adiposity, insulin sensitivity, and energy expenditure. J. Clin. Endocrinol. Metab. 1997, 82, 1293–1300. [Google Scholar] [CrossRef] [PubMed]
- Mutt, S.J.; Hypponen, E.; Saarnio, J.; Jarvelin, M.R.; Herzig, K.H. Vitamin D and adipose tissue-more than storage. Front. Physiol. 2014, 5, 228. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Dong, Y.; Bhagatwala, J.; Raed, A.; Huang, Y.; Zhu, H. Vitamin D3 Supplementation Increases Long-Chain Ceramide Levels in Overweight/Obese African Americans: A Post-Hoc Analysis of a Randomized Controlled Trial. Nutrients 2020, 12, 981. [Google Scholar] [CrossRef]
- Koch, A.; Grammatikos, G.; Trautmann, S.; Schreiber, Y.; Thomas, D.; Bruns, F.; Pfeilschifter, J.; Badenhoop, K.; Penna-Martinez, M. Vitamin D Supplementation Enhances C18(dihydro)ceramide Levels in Type 2 Diabetes Patients. Int. J. Mol. Sci. 2017, 18, 1532. [Google Scholar] [CrossRef]
- Kobayashi, H.; Amrein, K.; Lasky-Su, J.; Christopher, K.B. Procalcitonin Metabolomics in the Critically Ill reveal relationships between inflammation intensity and energy utilization pathways. Sci. Rep. 2021, 11, 23194. [Google Scholar] [CrossRef]
- Dimitrov, V.; Barbier, C.; Ismailova, A.; Wang, Y.; Dmowski, K.; Salehi-Tabar, R.; Memari, B.; Groulx-Boivin, E.; White, J.H. Vitamin D-regulated Gene Expression Profiles: Species-specificity and Cell-specific Effects on Metabolism and Immunity. Endocrinology 2021, 162. [Google Scholar] [CrossRef]
- Wallner, S.; Schmitz, G. Plasmalogens the neglected regulatory and scavenging lipid species. Chem. Phys. Lipids 2011, 164, 573–589. [Google Scholar] [CrossRef]
- Nalos, M.; Parnell, G.; Robergs, R.; Booth, D.; McLean, A.S.; Tang, B.M. Transcriptional reprogramming of metabolic pathways in critically ill patients. Intensive Care Med. Exp. 2016, 4, 21. [Google Scholar] [CrossRef]
- Jain, S.K.; Micinski, D. Vitamin D upregulates glutamate cysteine ligase and glutathione reductase, and GSH formation, and decreases ROS and MCP-1 and IL-8 secretion in high-glucose exposed U937 monocytes. Biochem. Biophys. Res. Commun. 2013, 437, 7–11. [Google Scholar] [CrossRef] [PubMed]
- Langley, R.J.; Tsalik, E.L.; van Velkinburgh, J.C.; Glickman, S.W.; Rice, B.J.; Wang, C.; Chen, B.; Carin, L.; Suarez, A.; Mohney, R.P.; et al. An integrated clinico-metabolomic model improves prediction of death in sepsis. Sci. Transl. Med. 2013, 5, 195ra195. [Google Scholar] [CrossRef] [PubMed]
- Koves, T.R.; Ussher, J.R.; Noland, R.C.; Slentz, D.; Mosedale, M.; Ilkayeva, O.; Bain, J.; Stevens, R.; Dyck, J.R.; Newgard, C.B.; et al. Mitochondrial overload and incomplete fatty acid oxidation contribute to skeletal muscle insulin resistance. Cell Metab. 2008, 7, 45–56. [Google Scholar] [CrossRef] [PubMed]
- Makrecka-Kuka, M.; Sevostjanovs, E.; Vilks, K.; Volska, K.; Antone, U.; Kuka, J.; Makarova, E.; Pugovics, O.; Dambrova, M.; Liepinsh, E. Plasma acylcarnitine concentrations reflect the acylcarnitine profile in cardiac tissues. Sci. Rep. 2017, 7, 17528. [Google Scholar] [CrossRef]
- Violante, S.; Ijlst, L.; Te Brinke, H.; Koster, J.; Tavares de Almeida, I.; Wanders, R.J.; Ventura, F.V.; Houten, S.M. Peroxisomes contribute to the acylcarnitine production when the carnitine shuttle is deficient. Biochim. Biophys. Acta 2013, 1831, 1467–1474. [Google Scholar] [CrossRef]
- Yin, Y.; Yu, Z.; Xia, M.; Luo, X.; Lu, X.; Ling, W. Vitamin D attenuates high fat diet-induced hepatic steatosis in rats by modulating lipid metabolism. Eur. J. Clin. Investig. 2012, 42, 1189–1196. [Google Scholar] [CrossRef]
- Marcotorchino, J.; Tourniaire, F.; Astier, J.; Karkeni, E.; Canault, M.; Amiot, M.J.; Bendahan, D.; Bernard, M.; Martin, J.C.; Giannesini, B.; et al. Vitamin D protects against diet-induced obesity by enhancing fatty acid oxidation. J. Nutr. Biochem. 2014, 25, 1077–1083. [Google Scholar] [CrossRef]
- Sergeev, I.N.; Song, Q. High vitamin D and calcium intakes reduce diet-induced obesity in mice by increasing adipose tissue apoptosis. Mol. Nutr. Food Res. 2014, 58, 1342–1348. [Google Scholar] [CrossRef]
- Latham, C.M.; Brightwell, C.R.; Keeble, A.R.; Munson, B.D.; Thomas, N.T.; Zagzoog, A.M.; Fry, C.S.; Fry, J.L. Vitamin D Promotes Skeletal Muscle Regeneration and Mitochondrial Health. Front. Physiol. 2021, 12, 660498. [Google Scholar] [CrossRef]
- Sinha, A.; Hollingsworth, K.G.; Ball, S.; Cheetham, T. Improving the vitamin D status of vitamin D deficient adults is associated with improved mitochondrial oxidative function in skeletal muscle. J. Clin. Endocrinol. Metab. 2013, 98, E509–E513. [Google Scholar] [CrossRef]
- Ventura-Clapier, R.; Moulin, M.; Piquereau, J.; Lemaire, C.; Mericskay, M.; Veksler, V.; Garnier, A. Mitochondria: A central target for sex differences in pathologies. Clin. Sci. 2017, 131, 803–822. [Google Scholar] [CrossRef] [PubMed]
- Malorni, W.; Campesi, I.; Straface, E.; Vella, S.; Franconi, F. Redox features of the cell: A gender perspective. Antioxid. Redox Signal. 2007, 9, 1779–1801. [Google Scholar] [CrossRef] [PubMed]
- Vina, J.; Borras, C. Women live longer than men: Understanding molecular mechanisms offers opportunities to intervene by using estrogenic compounds. Antioxid. Redox Signal. 2010, 13, 269–278. [Google Scholar] [CrossRef] [PubMed]
- Demarest, T.G.; McCarthy, M.M. Sex differences in mitochondrial (dys)function: Implications for neuroprotection. J. Bioenerg. Biomembr. 2015, 47, 173–188. [Google Scholar] [CrossRef]
- Austad, S.N.; Fischer, K.E. Sex Differences in Lifespan. Cell Metab. 2016, 23, 1022–1033. [Google Scholar] [CrossRef]
- Bruls, Y.M.; de Ligt, M.; Lindeboom, L.; Phielix, E.; Havekes, B.; Schaart, G.; Kornips, E.; Wildberger, J.E.; Hesselink, M.K.; Muoio, D.; et al. Carnitine supplementation improves metabolic flexibility and skeletal muscle acetylcarnitine formation in volunteers with impaired glucose tolerance: A randomised controlled trial. EBioMedicine 2019, 49, 318–330. [Google Scholar] [CrossRef]
- Zhang, J.; Light, A.R.; Hoppel, C.L.; Campbell, C.; Chandler, C.J.; Burnett, D.J.; Souza, E.C.; Casazza, G.A.; Hughen, R.W.; Keim, N.L.; et al. Acylcarnitines as markers of exercise-associated fuel partitioning, xenometabolism, and potential signals to muscle afferent neurons. Exp. Physiol. 2017, 102, 48–69. [Google Scholar] [CrossRef]
- Holloway, G.P.; Bezaire, V.; Heigenhauser, G.J.; Tandon, N.N.; Glatz, J.F.; Luiken, J.J.; Bonen, A.; Spriet, L.L. Mitochondrial long chain fatty acid oxidation, fatty acid translocase/CD36 content and carnitine palmitoyltransferase I activity in human skeletal muscle during aerobic exercise. J. Physiol. 2006, 571, 201–210. [Google Scholar] [CrossRef]
- Bezaire, V.; Bruce, C.R.; Heigenhauser, G.J.; Tandon, N.N.; Glatz, J.F.; Luiken, J.J.; Bonen, A.; Spriet, L.L. Identification of fatty acid translocase on human skeletal muscle mitochondrial membranes: Essential role in fatty acid oxidation. Am. J. Physiol. Endocrinol. Metab. 2006, 290, E509–E515. [Google Scholar] [CrossRef]
- Kiens, B.; Roepstorff, C.; Glatz, J.F.; Bonen, A.; Schjerling, P.; Knudsen, J.; Nielsen, J.N. Lipid-binding proteins and lipoprotein lipase activity in human skeletal muscle: Influence of physical activity and gender. J. Appl. Physiol. 2004, 97, 1209–1218. [Google Scholar] [CrossRef]
- Maher, A.C.; Akhtar, M.; Vockley, J.; Tarnopolsky, M.A. Women have higher protein content of beta-oxidation enzymes in skeletal muscle than men. PLoS ONE 2010, 5, e12025. [Google Scholar] [CrossRef] [PubMed]
- Newgard, C.B. Interplay between lipids and branched-chain amino acids in development of insulin resistance. Cell Metab. 2012, 15, 606–614. [Google Scholar] [CrossRef] [PubMed]
- Vickers, A.J. How many repeated measures in repeated measures designs? Statistical issues for comparative trials. BMC Med. Res. Methodol. 2003, 3, 22. [Google Scholar] [CrossRef] [PubMed]
- Mei, Y.; Kim, B.S.; Tsui, K. Linear mixed effects models for feature selection in high dimensional NMR spectra. Exprt Syst. Applic. 2009, 36, 4703–4708. [Google Scholar] [CrossRef]
- Ernest, B.; Gooding, J.R.; Campagna, S.R.; Saxton, A.M.; Voy, B.H. MetabR: An R script for linear model analysis of quantitative metabolomic data. BMC Res. Notes 2012, 5, 596. [Google Scholar] [CrossRef]
- Benjamini, Y.; Yekutieli, D. The control of the false discovery rate in multiple testing under dependency. Ann. Stat. 2001, 29, 1165–1188. [Google Scholar] [CrossRef]
- Cheng, J.B.; Motola, D.L.; Mangelsdorf, D.J.; Russell, D.W. De-orphanization of cytochrome P450 2R1: A microsomal vitamin D 25-hydroxilase. J. Biol. Chem. 2003, 278, 38084–38093. [Google Scholar] [CrossRef]
- Yamasaki, T.; Izumi, S.; Ide, H.; Ohyama, Y. Identification of a novel rat microsomal vitamin D3 25-hydroxylase. J. Biol. Chem. 2004, 279, 22848–22856. [Google Scholar] [CrossRef]
- Seamans, K.M.; Cashman, K.D. Existing and potentially novel functional markers of vitamin D status: A systematic review. Am. J. Clin. Nutr. 2009, 89, 1997S–2008S. [Google Scholar] [CrossRef]
- Chong, J.; Xia, J. Using MetaboAnalyst 4.0 for Metabolomics Data Analysis, Interpretation, and Integration with Other Omics Data. Methods Mol. Biol. 2020, 2104, 337–360. [Google Scholar] [CrossRef]
- Henglin, M.; Niiranen, T.; Watrous, J.D.; Lagerborg, K.A.; Antonelli, J.; Claggett, B.L.; Demosthenes, E.J.; von Jeinsen, B.; Demler, O.; Vasan, R.S.; et al. A Single Visualization Technique for Displaying Multiple Metabolite-Phenotype Associations. Metabolites 2019, 9, 128. [Google Scholar] [CrossRef] [PubMed]
- Dustin, T.; Yamamoto, T.; Hirose, K.; Keele, L.; Imai, K. mediation: R package for causal mediation analysis. J. Stat. Softw. 2014, 59, 1–38. [Google Scholar]
- Pietzner, M.; Budde, K.; Homuth, G.; Kastenmuller, G.; Henning, A.K.; Artati, A.; Krumsiek, J.; Volzke, H.; Adamski, J.; Lerch, M.M.; et al. Hepatic Steatosis Is Associated with Adverse Molecular Signatures in Subjects Without Diabetes. J. Clin. Endocrinol. Metab. 2018, 103, 3856–3868. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
| Characteristic | Absolute Increase in 25(OH)D Level between Day 0 and Day 3 | Total | p-Value | |||
|---|---|---|---|---|---|---|
| 25(OH)D ≥ 7.5 ng/mL | 25(OH)D < 7.5 ng/mL | |||||
| Female | Male | Female | Male | |||
| No. | 59 | 97 | 92 | 180 | 428 | |
| Age (Years) Mean (SD) | 66.5 (15.6) | 63.3 (16.4) | 69.3 (11.5) | 61.2 (14.7) | 64.2 (14.9) | <0.001 * |
| SAPS II Mean (SD) | 34.0 (16.2) | 33.0 (17.0) | 34.9 (13.8) | 32.5 (15.1) | 33.4 (15.4) | 0.67 * |
| CRP μg/mL Day 0 Mean (SD) | 125.9 (101.8) | 125.2 (93.6) | 116.0 (93.2) | 128.9 (81.9) | 124.9 (89.8) | 0.74 * |
| Day 0 25(OH)D ng/mL Mean (SD) | 12.5 (4.5) | 15.6 (15.7) | 13.6 (6.4) | 13.7 (4.9) | 14 (8.8) | 0.15 * |
| Vitamin D3 Intervention No. (%) | 57 (97) | 96 (99) | 21 (23) | 38 (21) | 212 (50) | <0.001 |
| Change in 25(OH)D ng/mL Median (IQR) | 22.3 [14.7, 37.1] | 22.2 [14.7, 38.2] | 0.5 [−1.6, 2.5] | 0.6 [−1.2, 2.9] | 3.3 [0, 16.7] | <0.001 ‡ |
| BMI Mean (SD) | 26.7 (4.8) | 27.2 (4.6) | 27.5 (5.8) | 26.9 (5.4) | 27.1 (5.2) | 0.75 * |
| Charlson Comorbidity Index Mean (SD) | 2.2 (1.6) | 3.1 (2.4) | 3.2 (2.1) | 3.0 (2.2) | 3.0 (2.2) | <0.001 * |
| Diabetes History No. (%) | 15 (25) | 19 (20) | 24 (26) | 43 (24) | 101 (24) | 0.73 |
| Glucose Day 0 Mean (SD) | 167.7 (64.5) | 147.5 (46.1) | 153.5 (50.7) | 144.2 (48.5) | 150.3 (51.4) | 0.025 * |
| ICU | 0.13 | |||||
| Anesthesia ICU No. (%) | 10 (17) | 15 (16) | 14 (15) | 42 (24) | 81 (19) | |
| Cardiac Surgery ICU No. (%) | 13 (22) | 24 (25) | 27 (30) | 59 (33) | 123 (29) | |
| Medicine ICU No. (%) | 12 (21) | 19 (20) | 19 (21) | 39 (22) | 89 (21) | |
| Neurological ICU No. (%) | 21 (36) | 30 (32) | 26 (29) | 29 (16) | 106 (25) | |
| Surgical No. (%) | 2 (3) | 7 (7) | 5 (5) | 8 (5) | 22 (5) | |
| 28-day Mortality No. (%) | 11 (19) | 11 (11) | 23 (25) | 50 (28) | 95 (22) | 0.013 |
| Metabolite | Female Stratum | Male Stratum | Sub-Pathway | ||||
|---|---|---|---|---|---|---|---|
| β Coefficient | p-Value | q-Value | β Coefficient | p-Value | q-Value | ||
| Palmitoylcarnitine (C16) | 0.004593 | 2.65 × 10−3 | 5.83 × 10−2 | −0.00156 | 2.76 × 10−1 | 4.96 × 10−1 | Long-chain Acylcarnitine |
| Margaroylcarnitine (C17) * | 0.005591 | 1.19 × 10−3 | 5.27 × 10−2 | −0.0007 | 6.47 × 10−1 | 7.81 × 10−1 | Long-chain Acylcarnitine |
| Stearoylcarnitine (C18) | 0.006447 | 1.08 × 10−5 | 7.89 × 10−3 | 0.0000581 | 9.67 × 10−1 | 9.81 × 10−1 | Long-chain Acylcarnitine |
| Linoleoylcarnitine (C18:2) * | 0.004122 | 1.11 × 10−2 | 9.81 × 10−2 | 0.001059 | 4.35 × 10−1 | 6.29 × 10−1 | Long-chain Acylcarnitine |
| Dihomo-linolenoylcarnitine (C20:3n3 or 6) * | 0.005715 | 7.64 × 10−4 | 4.15 × 10−2 | 0.001106 | 4.73 × 10−1 | 6.55 × 10−1 | Long-chain Acylcarnitine |
| Arachidonoylcarnitine (C20:4) | 0.004813 | 6.21 × 10−3 | 7.59 × 10−2 | 0.001038 | 5.10 × 10−1 | 6.78 × 10−1 | Long-chain Acylcarnitine |
| Arachidoylcarnitine (C20) * | 0.004313 | 1.21 × 10−2 | 9.89 × 10−2 | 0.000811 | 5.71 × 10−1 | 7.29 × 10−1 | Long-chain Acylcarnitine |
| Adrenoylcarnitine (C22:4) * | 0.00495 | 5.79 × 10−3 | 7.59 × 10−2 | 0.001161 | 4.98 × 10−1 | 6.68 × 10−1 | Long-chain Acylcarnitine |
| Lignoceroylcarnitine (C24) * | 0.004725 | 1.17 × 10−3 | 5.27 × 10−2 | −0.00017 | 9.02 × 10−1 | 9.43 × 10−1 | Long-chain Acylcarnitine |
| Cerotoylcarnitine (C26) * | 0.004831 | 1.86 × 10−3 | 5.50 × 10−2 | 0.000254 | 8.68 × 10−1 | 9.21 × 10−1 | Long-chain Acylcarnitine |
| Metabolite | Female Stratum | Male Stratum | Sub-Pathway | ||||
|---|---|---|---|---|---|---|---|
| β Coefficient | p-Value | q-Value | β Coefficient | p-Value | q-Value | ||
| Ceramide (d18:1/17:0, d17:1/18:0) * | −0.00018 | 9.15 × 10−1 | 9.60 × 10−1 | −0.00487 | 1.84 × 10−3 | 4.49 × 10−2 | Ceramide |
| N-stearoyl-sphingosine (d18:1/18:0) * | −0.00159 | 3.62 × 10−1 | 5.96 × 10−1 | −0.00494 | 1.59 × 10−3 | 4.08 × 10−2 | Ceramide |
| N-palmitoyl-sphingosine (d18:1/16:0) | −0.00208 | 1.87 × 10−1 | 4.10 × 10−1 | −0.00496 | 9.80 × 10−4 | 2.95 × 10−2 | Ceramide |
| N-stearoyl-sphingadienine (d18:2/18:0) * | −0.00199 | 2.73 × 10−1 | 5.08 × 10−1 | −0.00525 | 6.44 × 10−4 | 2.28 × 10−2 | Ceramide |
| Suberate (C8-DC) | −0.00435 | 1.79 × 10−2 | 1.23 × 10−1 | −0.00513 | 6.47 × 10−3 | 7.18 × 10−2 | Fatty Acid, Dicarboxylate |
| Heptenedioate (C7:1-DC) * | −0.00161 | 5.08 × 10−1 | 7.24 × 10−1 | −0.00535 | 1.27 × 10−2 | 9.53 × 10−2 | Fatty Acid, Dicarboxylate |
| 3-methyladipate | −0.00469 | 3.39 × 10−2 | 1.62 × 10−1 | −0.00648 | 5.36 × 10−3 | 6.71 × 10−2 | Fatty Acid, Dicarboxylate |
| 2-hydroxyadipate | −0.00377 | 8.30 × 10−2 | 2.65 × 10−1 | −0.00652 | 5.18 × 10−3 | 6.65 × 10−2 | Fatty Acid, Dicarboxylate |
| 3-hydroxyadipate * | −0.00247 | 3.31 × 10−1 | 5.68 × 10−1 | −0.00837 | 3.84 × 10−4 | 1.72 × 10−2 | Fatty Acid, Dicarboxylate |
| Arachidate (20:0) | −0.00073 | 5.61 × 10−1 | 7.56 × 10−1 | −0.00287 | 7.31 × 10−3 | 7.28 × 10−2 | Long Chain Fatty Acid |
| Oleate/vaccenate (18:1) | −0.00063 | 6.90 × 10−1 | 8.42 × 10−1 | −0.00357 | 6.20 × 10−3 | 7.18 × 10−2 | Long Chain Fatty Acid |
| Eicosenoate (20:1n9 or 1n11) | −0.00109 | 5.16 × 10−1 | 7.28 × 10−1 | −0.00383 | 6.73 × 10−3 | 7.28 × 10−2 | Long Chain Fatty Acid |
| Erucate (22:1n9) | −0.00221 | 1.72 × 10−1 | 3.90 × 10−1 | −0.00553 | 5.62 × 10−5 | 7.48 × 10−3 | Long Chain Fatty Acid |
| Docosapentaenoate (22:5n3) | −0.00108 | 5.60 × 10−1 | 7.55 × 10−1 | −0.00391 | 9.29 × 10−3 | 8.36 × 10−2 | Polyunsaturated Fatty Acid |
| Eicosapentaenoate (20:5n3) | −0.00402 | 5.46 × 10−2 | 2.10 × 10−1 | −0.00503 | 1.07 × 10−2 | 8.94 × 10−2 | Polyunsaturated Fatty Acid |
| Stearidonate (18:4n3) | −0.00428 | 8.68 × 10−2 | 2.70 × 10−1 | −0.00595 | 7.82 × 10−3 | 7.63 × 10−2 | Polyunsaturated Fatty Acid |
| Docosatrienoate (22:3n6) * | −0.00118 | 5.86 × 10−1 | 7.67 × 10−1 | −0.00759 | 6.13 × 10−5 | 7.48 × 10−3 | Polyunsaturated Fatty Acid |
| Metabolite | Female Stratum | Male Stratum | Sub-Pathway | ||||
|---|---|---|---|---|---|---|---|
| β Coefficient | p-Value | q-Value | β Coefficient | p-Value | q-Value | ||
| 3-methyl-2-oxovalerate | 0.00454 | 4.82 × 10−4 | 3.05 × 10−2 | 0.00372 | 2.47 × 10−3 | 5.24 × 10−2 | BCAA Metabolism |
| 4-methyl-2-oxopentanoate | 0.00357 | 1.02 × 10−2 | 9.48 × 10−2 | 0.00370 | 3.78 × 10−3 | 5.98 × 10−2 | BCAA Metabolism |
| Leucine | 0.00293 | 4.96 × 10−3 | 7.24 × 10−2 | 0.00372 | 2.41 × 10−4 | 1.34 × 10−2 | BCAA Metabolism |
| Isoleucine | 0.00293 | 5.01 × 10−3 | 7.24 × 10−2 | 0.00313 | 3.32 × 10−4 | 1.62 × 10−2 | BCAA Metabolism |
| 1-(1-enyl-stearoyl)-2-arachidonoyl-GPE (P-18:0/20:4) * | 0.00708 | 1.61 × 10−5 | 7.89 × 10−3 | 0.00563 | 1.17 × 10−4 | 1.16 × 10−2 | Plasmalogen |
| 1-(1-enyl-stearoyl)-2-linoleoyl-GPE (P-18:0/18:2) * | 0.00559 | 2.93 × 10−4 | 2.38 × 10−2 | 0.00514 | 1.66 × 10−5 | 5.39 × 10−3 | Plasmalogen |
| 1-(1-enyl-palmitoyl)-2-linoleoyl-GPE (P-16:0/18:2) * | 0.00551 | 2.03 × 10−3 | 5.50 × 10−2 | 0.00693 | 3.95 × 10−6 | 2.14 × 10−3 | Plasmalogen |
| 1-(1-enyl-palmitoyl)-2-arachidonoyl-GPE (P-16:0/20:4) * | 0.00484 | 5.14 × 10−4 | 3.05 × 10−2 | 0.00438 | 2.48 × 10−4 | 1.34 × 10−2 | Plasmalogen |
| 1-(1-enyl-palmitoyl)-2-arachidonoyl-GPC (P-16:0/20:4) * | 0.00465 | 2.45 × 10−3 | 5.76 × 10−2 | 0.00522 | 1.20 × 10−4 | 1.16 × 10−2 | Plasmalogen |
| 1-(1-enyl-palmitoyl)-2-linoleoyl-GPC (P-16:0/18:2) * | 0.00448 | 4.58 × 10−3 | 7.01 × 10−2 | 0.00434 | 2.47 × 10−3 | 5.24 × 10−2 | Plasmalogen |
| 1-(1-enyl-palmitoyl)-2-palmitoyl-GPC (P-16:0/16:0) * | 0.00414 | 2.43 × 10−4 | 2.22 × 10−2 | 0.00414 | 2.43 × 10−4 | 1.34 × 10−2 | Plasmalogen |
| Ribitol | −0.00464 | 1.01 × 10−2 | 9.48 × 10−2 | −0.00395 | 6.71 × 10−3 | 7.28 × 10−2 | Pentose Metabolism |
| Gluconate | −0.00940 | 5.22 × 10−3 | 7.40 × 10−2 | −0.01015 | 5.30 × 10−4 | 2.15 × 10−2 | Pentose Metabolism |
| Arabitol/xylitol | −0.00628 | 2.01 × 10−3 | 5.50 × 10−2 | −0.00463 | 6.97 × 10−3 | 7.28 × 10−2 | Pentose Metabolism |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chary, S.; Amrein, K.; Mahmoud, S.H.; Lasky-Su, J.A.; Christopher, K.B. Sex-Specific Catabolic Metabolism Alterations in the Critically Ill following High Dose Vitamin D. Metabolites 2022, 12, 207. https://doi.org/10.3390/metabo12030207
Chary S, Amrein K, Mahmoud SH, Lasky-Su JA, Christopher KB. Sex-Specific Catabolic Metabolism Alterations in the Critically Ill following High Dose Vitamin D. Metabolites. 2022; 12(3):207. https://doi.org/10.3390/metabo12030207
Chicago/Turabian StyleChary, Sowmya, Karin Amrein, Sherif H. Mahmoud, Jessica A. Lasky-Su, and Kenneth B. Christopher. 2022. "Sex-Specific Catabolic Metabolism Alterations in the Critically Ill following High Dose Vitamin D" Metabolites 12, no. 3: 207. https://doi.org/10.3390/metabo12030207
APA StyleChary, S., Amrein, K., Mahmoud, S. H., Lasky-Su, J. A., & Christopher, K. B. (2022). Sex-Specific Catabolic Metabolism Alterations in the Critically Ill following High Dose Vitamin D. Metabolites, 12(3), 207. https://doi.org/10.3390/metabo12030207