
Melatonin Supplementation Alters Maternal and Fetal Amino Acid Concentrations and Placental Nutrient Transporters in a Nutrient Restriction Bovine Model

Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals and Experimental Design
2.2. Blood Parameters
2.3. Placentome Collection and Separation
2.4. Gene Expression
2.5. Statistical Analysis
3. Results
3.1. Plasma Amino Acids
3.2. Placenta Transcript Abundance
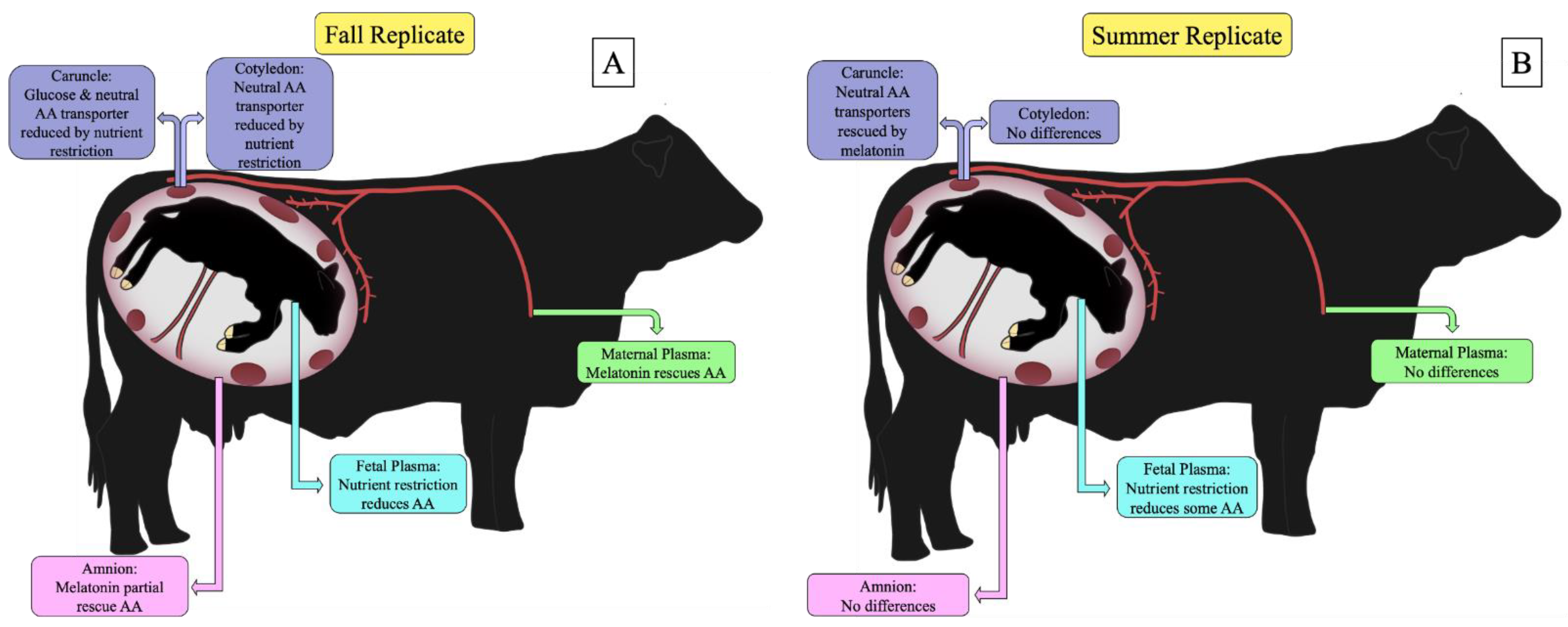
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Reynolds, L.P.; Redmer, D.A. Utero-placental vascular development and placental function. J. Anim. Sci. 1995, 73, 1839–1851. [Google Scholar] [CrossRef] [PubMed]
- Wu, G.; Bazer, F.W.; Wallace, J.M.; Spencer, T.E. Board-invited review: Intrauterine growth retardation: Implications for the animal sciences. J. Anim. Sci. 2006, 84, 2316–2337. [Google Scholar] [CrossRef] [PubMed]
- Lemley, C.O.; Meyer, A.M.; Camacho, L.E.; Neville, T.L.; Newman, D.J.; Caton, J.S.; Vonnahme, K.A. Melatonin supplementation alters uteroplacental hemodynamics and fetal development in an ovine model of intrauterine growth restriction. Am. J. Physiol. -Regul. Integr. Comp. Physiol. 2012, 302, R454–R467. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Contreras-Correa, Z.E.; Messman, R.D.; Sidelinger, D.R.; Heath King, E.; Sánchez-Rodríguez, H.L.; Burnett, D.D.; Lemley, C.O. Melatonin alters bovine uterine artery hemodynamics, vaginal temperatures, and fetal morphometrics during late gestational nutrient restriction in a season-dependent manner. J. Anim. Sci. 2021, 99, skab242. [Google Scholar] [CrossRef]
- Lemley, C.O.; Hart, C.G.; Lemire, R.L.; King, E.H.; Hopper, R.M.; Park, S.B.; Rude, B.J.; Burnett, D.D. Maternal nutrient restriction alters uterine artery hemodynamics and placentome vascular density in Bos indicus and Bos taurus. J. Anim. Sci. 2018, 96, 4823–4834. [Google Scholar] [CrossRef]
- Edwards, A.K.; McKnight, S.M.; Askelson, K.; McKnight, J.R.; Dunlap, K.A.; Satterfield, M.C. Adaptive responses to maternal nutrient restriction alter placental transport in ewes. Placenta 2020, 96, 1–9. [Google Scholar] [CrossRef]
- Crouse, M.S.; McLean, K.J.; Greseth, N.P.; Ward, A.K.; Reynolds, L.P.; Dahlen, C.R.; Neville, B.W.; Borowicz, P.P.; Caton, J.S. The effects of maternal nutrient restriction and day of early pregnancy on the location and abundance of neutral amino acid transporters in beef heifer utero-placental tissues. J. Anim. Sci. 2020, 98, skaa197. [Google Scholar] [CrossRef]
- Paulis, L.; Šimko, F. Blood pressure modulation and cardiovascular protection by melatonin: Potential mechanisms behind. Physiol. Res. 2007, 56, 671–684. [Google Scholar] [CrossRef]
- Brockus, K.E.; Hart, C.G.; Gilfeather, C.L.; Fleming, B.O.; Lemley, C.O. Dietary melatonin alters uterine artery hemodynamics in pregnant Holstein heifers. Domest. Anim. Endocrinol. 2016, 55, 1–10. [Google Scholar] [CrossRef]
- Simko, F.; Paulis, L. Melatonin as a potential antihypertensive treatment. J. Pineal Res. 2007, 42, 319–322. [Google Scholar] [CrossRef]
- Lin, G.; Wang, X.; Wu, G.; Feng, C.; Zhou, H.; Li, D.; Wang, J. Improving amino acid nutrition to prevent intrauterine growth restriction in mammals. Amino Acids 2014, 46, 1605–1623. [Google Scholar] [CrossRef] [PubMed]
- Wu, G.; Bazer, F.W.; Satterfield, M.C.; Li, X.; Wang, X.; Johnson, G.A.; Burghardt, R.C.; Dai, Z.; Wang, J.; Wu, Z. Impacts of arginine nutrition on embryonic and fetal development in mammals. Amino Acids 2013, 45, 241–256. [Google Scholar] [CrossRef] [PubMed]
- Wu, G.; Bazer, F.W.; Datta, S.; Johnson, G.A.; Li, P.; Satterfield, M.C.; Spencer, T.E. Proline metabolism in the conceptus: Implications for fetal growth and development. Amino Acids 2008, 35, 691–702. [Google Scholar] [CrossRef] [PubMed]
- Iruloh, C.G.; D′Souza, S.W.; Fergusson, W.D.; Baker, P.N.; Sibley, C.P.; Glazier, J.D. Amino Acid Transport Systems β and A in Fetal T Lymphocytes in Intrauterine Growth Restriction and With Tumor Necrosis Factor-α Treatment. Pediatr. Res. 2009, 65, 51–56. [Google Scholar] [CrossRef] [Green Version]
- Jansson, T. Amino Acid Transporters in the Human Placenta. Pediatr. Res. 2001, 49, 141–147. [Google Scholar] [CrossRef] [Green Version]
- Regnault, T.R.H.; Friedman, J.E.; Wilkening, R.B.; Anthony, R.V.; Hay, W.W., Jr. Fetoplacental transport and utilization of amino acids in IUGR—A review. Placenta 2005, 26, S52–S62. [Google Scholar] [CrossRef]
- Kaspar, H.; Dettmer, K.; Gronwald, W.; Oefner, P.J. Automated GC–MS analysis of free amino acids in biological fluids. J. Chromatogr. B 2008, 870, 222–232. [Google Scholar] [CrossRef]
- Cain, A.J.; Lemley, C.O.; Walters, F.K.; Christiansen, D.L.; King, E.H.; Hopper, R.M. Pre-breeding beef heifer management and season affect mid to late gestation uterine artery hemodynamics. Theriogenology 2017, 87, 9–15. [Google Scholar] [CrossRef]
- King, G.J.; Macleod, G.K. Reproductive function in beef cows calving in the spring or fall. Anim. Reprod. Sci. 1984, 6, 255–266. [Google Scholar] [CrossRef]
- Caldwell, J.D.; Coffey, K.P.; Jennings, J.A.; Philipp, D.; Young, A.N.; Tucker, J.D.; Hubbell III, D.S.; Hess, T.; Looper, M.L.; West, C.P. Performance by spring and fall-calving cows grazing with full, limited, or no access to toxic Neotyphodium coenophialum-infected tall fescue. J. Anim. Sci. 2013, 91, 465–476. [Google Scholar] [CrossRef]
- Lemley, C.O.; Camacho, L.E.; Meyer, A.M.; Kapphahn, M.; Caton, J.S.; Vonnahme, K.A. Dietary melatonin supplementation alters uteroplacental amino acid flux during intrauterine growth restriction in ewes. Animal 2013, 7, 1500–1507. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chung, M.; Teng, C.; Timmerman, M.; Meschia, G.; Battaglia, F.C. Production and utilization of amino acids by ovine placenta in vivo. Am. J. Physiol. Endocrinol. Metab. 1998, 274, E13–E22. [Google Scholar] [CrossRef] [PubMed]
- Brown, L.D.; Rozance, P.J.; Thorn, S.R.; Friedman, J.E.; Hay, W.W., Jr. Acute supplementation of amino acids increases net protein accretion in IUGR fetal sheep. Am. J. Physiol. Endocrinol. Metab. 2012, 303, E352–E364. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wai, S.G.; Rozance, P.J.; Wesolowski, S.R.; Hay, W.W., Jr.; Brown, L.D. Prolonged amino acid infusion into intrauterine growth-restricted fetal sheep increases leucine oxidation rates. Am. J. Physiol. Endocrinol. Metab. 2018, 315, E1143–E1153. [Google Scholar] [CrossRef] [PubMed]
- Lassala, A.; Bazer, F.W.; Cudd, T.A.; Datta, S.; Keisler, D.H.; Satterfield, M.C.; Spencer, T.E.; Wu, G. Parenteral administration of L-arginine prevents fetal growth restriction in undernourished ewes. J. Nutr. 2010, 140, 1242–1248. [Google Scholar] [CrossRef] [Green Version]
- Kwon, H.; Ford, S.P.; Bazer, F.W.; Spencer, T.E.; Nathanielsz, P.W.; Nijland, M.J.; Hess, B.W.; Wu, G. Maternal Nutrient Restriction Reduces Concentrations of Amino Acids and Polyamines in Ovine Maternal and Fetal Plasma and Fetal Fluids1. Biol. Reprod. 2004, 71, 901–908. [Google Scholar] [CrossRef] [Green Version]
- Orczyk-Pawilowicz, M.; Jawien, E.; Deja, S.; Hirnle, L.; Zabek, A.; Mlynarz, P. Metabolomics of human amniotic fluid and maternal plasma during normal pregnancy. PLoS ONE 2016, 11, e0152740. [Google Scholar] [CrossRef] [Green Version]
- Brown, L.D.; Green, A.S.; Limesand, S.W.; Rozance, P.J. Maternal amino acid supplementation for intrauterine growth restriction. Front. Biosci. Sch. Ed. 2011, 3, 428. [Google Scholar] [CrossRef] [Green Version]
- Reid, D.S.; Burnett, D.D.; Contreras-Correa, Z.E.; Lemley, C.O. Differences in bovine placentome blood vessel density and transcriptomics in a mid to late-gestating maternal nutrient restriction model. Placenta 2022, 117, 122–130. [Google Scholar] [CrossRef]
- Forde, N.; Simintiras, C.A.; Sturmey, R.; Mamo, S.; Kelly, A.K.; Spencer, T.E.; Bazer, F.W.; Lonergan, P. Amino acids in the uterine luminal fluid reflects the temporal changes in transporter expression in the endometrium and conceptus during early pregnancy in cattle. PLoS ONE 2014, 9, e100010. [Google Scholar] [CrossRef]


{kind=link}
| Gene | Function | Assay ID | Accession | Amplicon | |
|---|---|---|---|---|---|
| SLC2A8 | Glucose and fructose transport | Bt03217733_m1 | NP_963286 | 80 | |
| SLC43A2 | Sodium independent amino acid transport | Bt03246452_m1 | NP_001069014.1 | 94 | |
| SLC7A5 | Large neutral amino acid transport | Bt03215392_m1 | NP_777038.1 | 64 | |
| SLC7A8 | Large neutral amino acid transport | Bt04312788_m1 | NP_001179818.1 | 66 | |
| SLC38A10 | Sodium dependent amino acid transport | Bt03267849_m1 | BC153282 | 56 | |
| SLC38A2 | System A amino acid transport | Bt03255119_m1 | NP_001075893.1 | 67 | |
| RPL19 | Reference gene | Bt03229687_g1 | NP_001035606.1 | 82 | |
| SF3A1 | Reference gene | Bt03254301_m1 | NP_001074979.1 | 60 | |
| Fall Trial | p-Value | ||||||
|---|---|---|---|---|---|---|---|
| Maternal Plasma | ADQ-CON | RES-CON | ADQ-MEL | RES-MEL | NUT | TRT | NUT*TRT |
| System A | 512.03 ± 28.96 c | 1040.38 ± 166.26 a | 685.55 ± 122.33 b | 514.80 ± 68.89 b | - | - | <0.01 |
| System Br | 431.58 ± 32.82 | 383.23 ± 37.62 | 461.19 ± 34.67 | 461.59 ± 30.56 | 0.62 | 0.07 | 0.38 |
| System Bo | 540.01 ± 35.26 | 44.88 ± 41.08 | 580.98 ± 37.86 | 592.64 ± 32.98 | 0.87 | 0.16 | 0.91 |
| System N | 342.97 ± 37.54 b | 710.26 ± 115.26 a | 468.48 ± 81.27 ab | 338.95 ± 66.97 b | - | - | <0.01 |
| Cationic | 107.41 ± 9.66 ab | 132.37 ± 9.82 ab | 133.72 ± 9.93 a | 106.32 ± 8.97 b | - | - | 0.01 |
| Anion | 76.72 ± 2.58 c | 244.38 ± 63.22 a | 107.68 ± 17.07 ab | 90.01 ± 32.37 bc | - | - | <0.01 |
| Total | 1883.94 ± 49.14 b | 2676.84 ± 271.37 a | 2165.16 ± 189.28 ab | 1897.11 ± 134.93 b | - | - | 0.01 |
| Essential | 649.61 ± 32.86 | 620.72 ± 32.86 | 715.21 ± 32.86 | 661.74 ± 30.66 | 0.21 | 0.11 | 0.71 |
| Fetal Plasma | |||||||
| System A | 1229.16 ± 137.88 | 1206.54 ± 137.88 | 1348.55 ± 137.88 | 1235.97 ± 128.66 | 0.62 | 0.59 | 0.74 |
| System Br | 363.87 ± 29.54 | 352.72 ± 30.34 | 344.12 ± 31.57 | 348.76 ± 27.94 | 0.92 | 0.69 | 0.79 |
| System Bo | 43.90 ± 54.90 | 510.92 ± 63.97 | 536.58 ± 58.95 | 518.16 ± 51.36 | 0.01 | 0.60 | 0.88 |
| System N | 770.52 ± 127.05 | 752.38 ± 127.05 | 883.46 ± 127.05 | 792.39 ± 118.55 | 0.67 | 0.55 | 0.77 |
| Cationic | 135.04 ± 13.56 | 130.53 ± 13.39 | 148.40 ± 13.39 | 129.37 ± 12.67 | 0.38 | 0.66 | 0.59 |
| Anion | 334.13 ± 43.75 | 256.46 ± 43.75 | 426.94 ± 43.75 | 385.70 ± 40.83 | 0.18 | 0.02 | 0.68 |
| Total | 3058.75 ± 207.49 | 2835.00 ± 207.49 | 3293.79 ± 207.49 | 3016.63 ± 193.61 | 0.23 | 0.32 | 0.90 |
| Essential | 634.49 ± 41.05 | 599.99 ± 42.17 | 615.92 ± 43.88 | 590.72 ± 38.83 | 0.50 | 0.74 | 0.91 |
| Amnion | |||||||
| System A | 145.84 ± 26.37 b | 364.57 ± 67.50 a | 481.58 ± 180.53 a | 258.89 ± 108.28 ab | - | - | 0.01 |
| System Br | 34.43 ± 6.89 b | 256.31 ± 104.84 a | 215.38 ± 86.18 a | 38.25 ± 8.06 b | - | - | <0.01 |
| System Bo | 67.16 ± 9.66 b | 338.88 ± 125.65 a | 301.57 ± 109.36 ab | 74.85 ± 11.96 b | - | - | 0.01 |
| System N | 92.84 ± 12.88 b | 222.94 ± 49.27 ab | 290.22 ± 104.09 a | 145.94 ± 48.72 ab | - | - | 0.02 |
| Cationic | 42.51 ± 3.70 | 89.64 ± 20.56 | 89.97 ± 21.96 | 48.66 ± 7.65 | - | - | 0.03 |
| Anion | 42.64 ± 6.03 | 63.34 ± 10.56 | 54.22 ± 7.59 | 46.60 ± 7.08 | 0.12 | 0.38 | 0.10 |
| Total | 525.96 ± 74.69 b | 1296.53 ± 279.26 ab | 1409.06 ± 386.44 a | 744.10 ± 230.51 ab | - | - | 0.02 |
| Essential | 104.61 ± 13.72 b | 416.74 ± 144.12 a | 378.91 ± 126.68 a | 117.58 ± 23.33 b | - | - | 0.01 |
| Summer Trial | p Value | ||||||
|---|---|---|---|---|---|---|---|
| Maternal Plasma | ADQ-CON | RES-CON | ADQ-MEL | RES-MEL | NUT | TRT | NUT*TRT |
| System A | 1020.97 ± 229.93 | 1076.96 ± 229.93 | 962.94 ± 229.93 | 859.07 ± 213.48 | 0.92 | 0.55 | 0.83 |
| System Br | 436.57 ± 29.94 ab | 376.50 ± 26.88 b | 412.14 ± 33.13 ab | 496.68 ± 27.46 a | - | - | 0.02 |
| System Bo | 596.56 ± 28.70 ab | 526.00 ± 28.62 b | 578.95 ± 34.49 ab | 645.74 ± 29.14 a | - | - | 0.03 |
| System N | 781.29 ± 148.09 | 724.37 ± 148.12 | 693.30 ± 148.04 | 637.47 ± 135.69 | 0.70 | 0.55 | 0.99 |
| Cationic | 107.23 ± 7.75 | 110.77 ± 9.80 | 133.75 ± 10.02 | 117.85 ± 10.39 | 0.59 | 0.09 | 0.45 |
| Anion | 116.99 ± 27.26 | 146.28 ± 44.54 | 179.18 ± 52.34 | 96.39 ± 25.46 | 0.77 | 0.89 | 0.37 |
| Total | 2481.94 ± 321.02 | 2525.55 ± 333.38 | 2753.47 ± 381.67 | 2146.38 ± 109.50 | 0.70 | 0.86 | 0.66 |
| Essential | 780.96 ± 38.02 | 625.66 ± 36.86 | 767.78 ± 48.44 | 729.11 ± 47.84 | 0.76 | 0.11 | 0.07 |
| Fetal Plasma | |||||||
| System A | 1418.83 ± 111.75 | 1387.76 ± 264.42 | 1440.70 ± 113.34 | 1627.40 ± 55.52 | 0.21 | 0.56 | 0.53 |
| System Br | 397.96 ± 25.89 | 296.81 ± 51.25 | 424.59 ± 26.73 | 367.26 ± 14.05 | 0.01 | 0.19 | 0.90 |
| System Bo | 591.95 ± 31.54 | 473.28 ± 78.81 | 634.72 ± 37.87 | 544.50 ± 20.38 | 0.15 | 0.77 | 0.59 |
| System N | 992.98 ± 123.99 | 957.46 ± 123.96 | 885.32 ± 128.04 | 1160.96 ± 117.25 | 0.14 | 0.47 | 0.06 |
| Cationic | 77.54 ± 10.43 | 61.12 ± 6.96 | 81.90 ± 9.43 | 63.99 ± 7.37 | 0.64 | 0.71 | 0.75 |
| Anion | 142.34 ± 19.36 | 129.18 ± 20.48 | 214.14 ± 20.46 | 196.44 ± 19.13 | 0.58 | 0.03 | 0.67 |
| Total | 2298.56 ± 121.57 | 2122.10 ± 354.06 | 2397.82 ± 133.17 | 2479.26 ± 78.33 | 0.23 | 0.03 | 0.20 |
| Essential | 687.99 ± 38.02 | 552.43 ± 87.25 | 712.19 ± 37.09 | 616.02 ± 12.43 | 0.02 | 0.25 | 0.83 |
| Amnion | |||||||
| System A | 524.93 ± 207.20 | 405.86 ± 60.36 | 355.05 ± 70.99 | 346.49 ± 89.86 | 0.80 | 0.22 | 0.84 |
| System Br | 331.21 ± 117.82 | 324.11 ± 95.51 | 196.19 ± 69.17 | 184.90 ± 91.63 | 0.50 | 0.09 | 0.72 |
| System Bo | 442.86 ± 146.00 | 420.16 ± 114.10 | 265.26 ± 83.17 | 247.35 ± 110.75 | 0.88 | 0.12 | 0.91 |
| System N | 294.15 ± 62.41 | 271.20 ± 52.98 | 218.59 ± 58.60 | 192.71 ± 55.30 | 0.67 | 0.19 | 0.98 |
| Cationic | 88.94 ± 14.20 | 85.73 ± 14.82 | 69.12 ± 10.52 | 93.47 ± 37.58 | 0.69 | 0.44 | 0.90 |
| Anion | 75.42 ± 15.42 | 49.62 ± 6.06 | 46.43 ± 5.07 | 45.01 ± 5.28 | 0.02 | 0.34 | 0.79 |
| Total | 1663.68 ± 341.91 | 1464.54 ± 290.30 | 1241.54 ± 314.54 | 1105.49 ± 299.78 | 0.60 | 0.22 | 0.92 |
| Essential | 517.51 ± 155.41 | 499.11 ± 127.43 | 340.53 ± 91.07 | 328.29 ± 116.51 | 0.38 | 0.18 | 0.74 |
| Fall Trial | p Value | ||||||
|---|---|---|---|---|---|---|---|
| Caruncle | ADQ-CON | RES-CON | ADQ-MEL | RES-MEL | NUT | TRT | NUT*TRT |
| SLC2A8 | 0.6893 ± 0.0617 | 1.8826 ± 0.5294 | 0.8714 ± 0.2051 | 1.0438 ± 0.3087 | 0.66 | 0.31 | 0.47 |
| SLC43A2 | 0.7092 ± 0.1151 | 0.9292 ± 0.2260 | 0.8478 ± 0.1176 | 1.8979 ± 0.3620 | 0.20 | 0.11 | 0.25 |
| SLC7A5 | 0.7013 ± 0.0774 | 1.7454 ± 0.5591 | 0.9267 ± 0.1778 | 1.2461 ± 0.2370 | 0.05 | 0.45 | 0.36 |
| SLC7A8 | 0.5305 ± 0.0709 | 0.5901 ± 0.1676 | 0.4050 ± 0.0725 | 0.5180 ± 0.0835 | 0.93 | 0.30 | 0.43 |
| SLC38A10 | 0.8010 ± 0.0760 | 1.0314 ± 0.1567 | 0.7262 ± 0.1105 | 1.1865 ± 0.1796 | 0.03 | 0.83 | 0.33 |
| SLC38A2 | 0.4988 ± 0.0956 | 0.8955 ± 0.2930 | 0.5765 ± 0.1260 | 0.6828 ± 0.1806 | 0.27 | 0.48 | 0.57 |
| Cotyledon | |||||||
| SLC2A8 | 1.8630 ± 0.6801 | 2.3565 ± 0.6405 | 1.2701 ± 0.2903 | 2.3602 ± 0.4184 | 0.03 | 0.95 | 0.66 |
| SLC43A2 | 0.9689 ± 0.2186 | 0.8167 ± 0.1563 | 0.7372 ± 0.1087 | 0.7485 ± 0.1581 | 0.63 | 0.78 | 0.74 |
| SLC7A5 | 1.3560 ± 0.1220 | 2.0552 ± 0.5263 | 2.0696 ± 0.6891 | 2.2455 ± 0.3947 | 0.50 | 0.35 | 0.82 |
| SLC7A8 | 1.0845 ± 0.2258 | 1.1113 ± 0.3165 | 1.7406 ± 0.3958 | 1.4869 ± 0.2127 | 0.57 | 0.02 | 0.35 |
| SLC38A10 | 1.2280 ± 0.1337 | 1.4432 ± 0.1348 | 1.3228 ± 0.1407 | 1.6269 ± 0.1280 | 0.09 | 0.31 | 0.74 |
| SLC38A2 | 0.7596 ± 0.1300 | 1.2885 ± 0.4477 | 1.0832 ± 0.3403 | 1.1360 ± 0.1771 | 0.14 | 0.45 | 0.96 |
| Summer Trial | p Value | ||||||
|---|---|---|---|---|---|---|---|
| Caruncle | ADQ-CON | RES-CON | ADQ-MEL | RES-MEL | NUT | TRT | NUT*TRT |
| SLC2A8 | 0.8645 ± 0.2889 | 0.9337 ± 0.2017 | 0.9708 ± 0.1910 | 0.7601 ± 0.1198 | 0.35 | 0.46 | 0.07 |
| SLC43A2 | 0.8030 ± 0.1411 | 0.8314 ± 0.1520 | 1.2378 ± 0.4489 | 0.7084 ± 0.0592 | 0.78 | 0.55 | 0.98 |
| SLC7A5 | 2.1421 ± 1.2201 | 1.7401 ± 0.4498 | 1.6200 ± 0.2052 | 1.1358 ± 0.2489 | 0.77 | 0.80 | 0.13 |
| SLC7A8 | 0.8551 ± 0.2062 | 1.2899 ± 0.2353 | 1.0087 ± 0.2291 | 0.8653 ± 0.1672 | 0.54 | 0.52 | 0.18 |
| SLC38A10 | 0.8291 ± 0.1746 b | 1.3427 ± 0.1732 a | 2.3497 ± 1.2254 ab | 0.9615 ± 0.1369 ab | - | - | 0.04 |
| SLC38A2 | 0.3514 ± 0.0560 b | 0.7786 ± 0.132 a | 0.5043 ± 0.0877 a | 0.4226 ± 0.0812 b | - | - | <0.01 |
| Cotyledon | |||||||
| SLC2A8 | 1.8430 ± 0.1905 | 1.8302 ± 0.2693 | 1.6650 ± 0.1620 | 1.3944 ± 0.2035 | 0.29 | 0.22 | 0.49 |
| SLC43A2 | 3.3572 ± 0.9071 | 5.4449 ± 1.2810 | 3.5628 ± 1.2810 | 2.4225 ± 1.1894 | 0.71 | 0.21 | 0.16 |
| SLC7A5 | 1.9922 ± 0.9210 | 1.4450 ± 0.2738 | 0.7286 ± 0.1116 | 0.9518 ± 0.2310 | 0.27 | 0.09 | 0.63 |
| SLC7A8 | 0.6057 ± 0.0706 | 0.5407 ± 0.0849 | 1.1313 ± 0.3202 | 0.8001 ± 0.2779 | 0.67 | 0.71 | 0.60 |
| SLC38A10 | 0.8802 ± 0.2273 | 0.9702 ± 0.1828 | 1.7643 ± 0.4429 | 0.9625 ± 0.1304 | 0.55 | 0.17 | 0.12 |
| SLC38A2 | 1.2958 ± 0.3566 | 1.3249 ± 0.4019 | 1.5967 ± 0.6684 | 1.9284 ± 1.2245 | 0.85 | 0.52 | 0.90 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Swanson, R.; Contreras-Correa, Z.; Dinh, T.; King, H.; Sidelinger, D.; Burnett, D.; Lemley, C. Melatonin Supplementation Alters Maternal and Fetal Amino Acid Concentrations and Placental Nutrient Transporters in a Nutrient Restriction Bovine Model. Metabolites 2022, 12, 1208. https://doi.org/10.3390/metabo12121208
Swanson R, Contreras-Correa Z, Dinh T, King H, Sidelinger D, Burnett D, Lemley C. Melatonin Supplementation Alters Maternal and Fetal Amino Acid Concentrations and Placental Nutrient Transporters in a Nutrient Restriction Bovine Model. Metabolites. 2022; 12(12):1208. https://doi.org/10.3390/metabo12121208
Chicago/Turabian StyleSwanson, Rebecca, Zully Contreras-Correa, Thu Dinh, Heath King, Darcie Sidelinger, Derris Burnett, and Caleb Lemley. 2022. "Melatonin Supplementation Alters Maternal and Fetal Amino Acid Concentrations and Placental Nutrient Transporters in a Nutrient Restriction Bovine Model" Metabolites 12, no. 12: 1208. https://doi.org/10.3390/metabo12121208