
Spatially Mapping the Baseline and Bisphenol-A Exposed Daphnia magna Lipidome Using Desorption Electrospray Ionization—Mass Spectrometry


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Application of Desorption Electrospray Ionization—Mass Specrometry to Daphnia magna
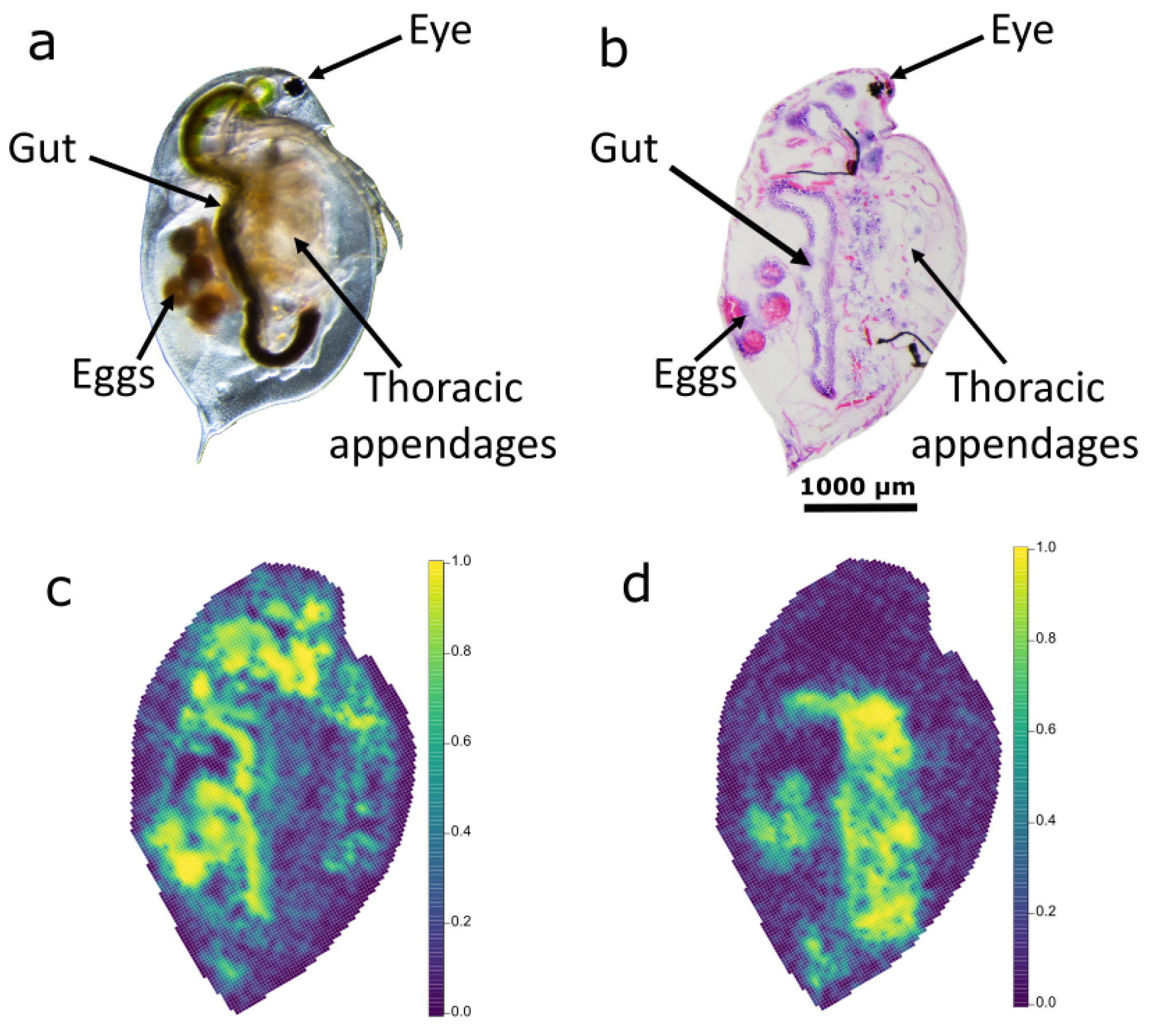
2.1.1. Preparation of Tissue Sections
2.1.2. Quality Control Strategies to Maximize DESI-MS Image Quality
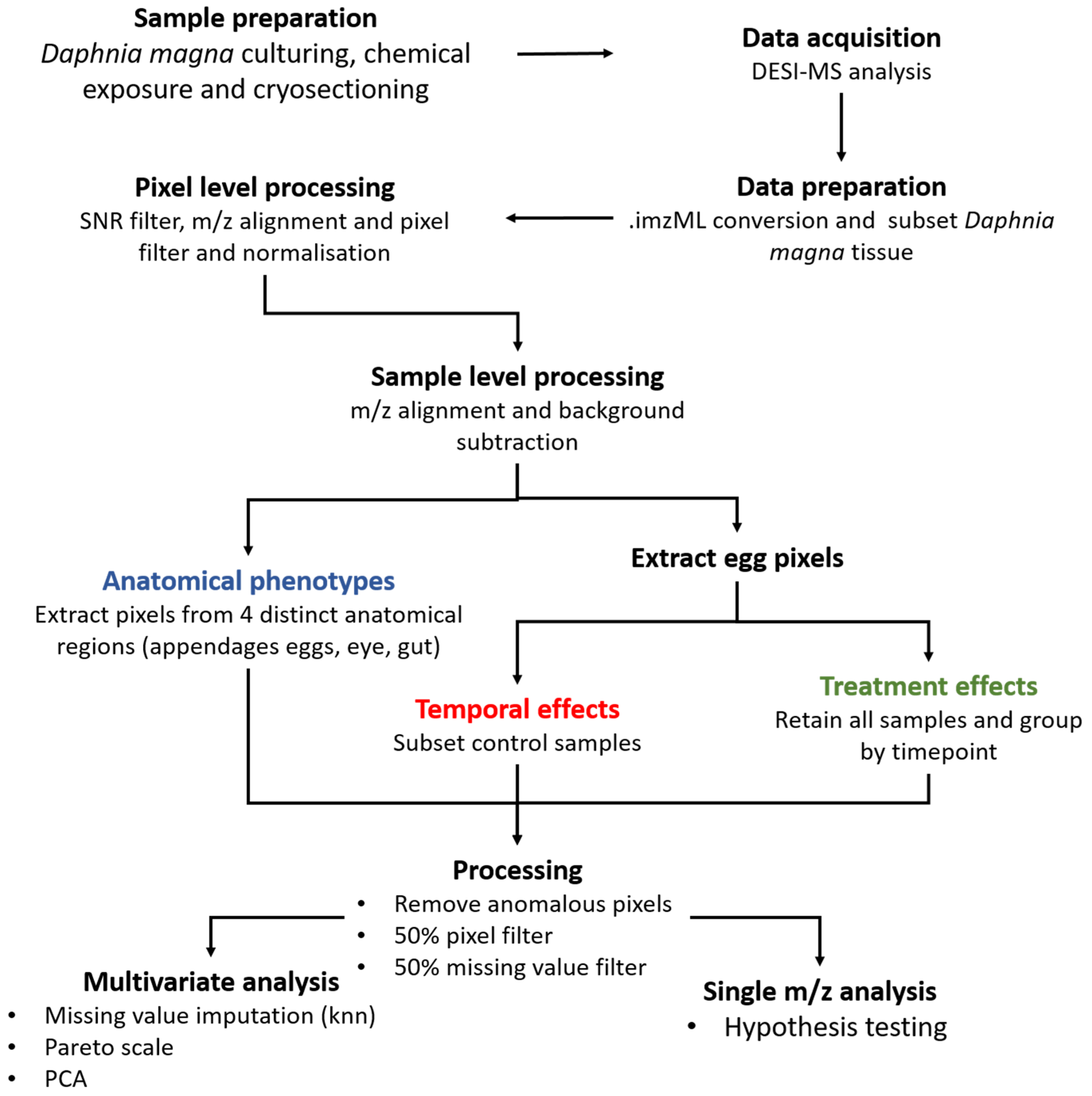
2.1.3. Signal Processing the Study Samples to Yield Reliable Spatial Information
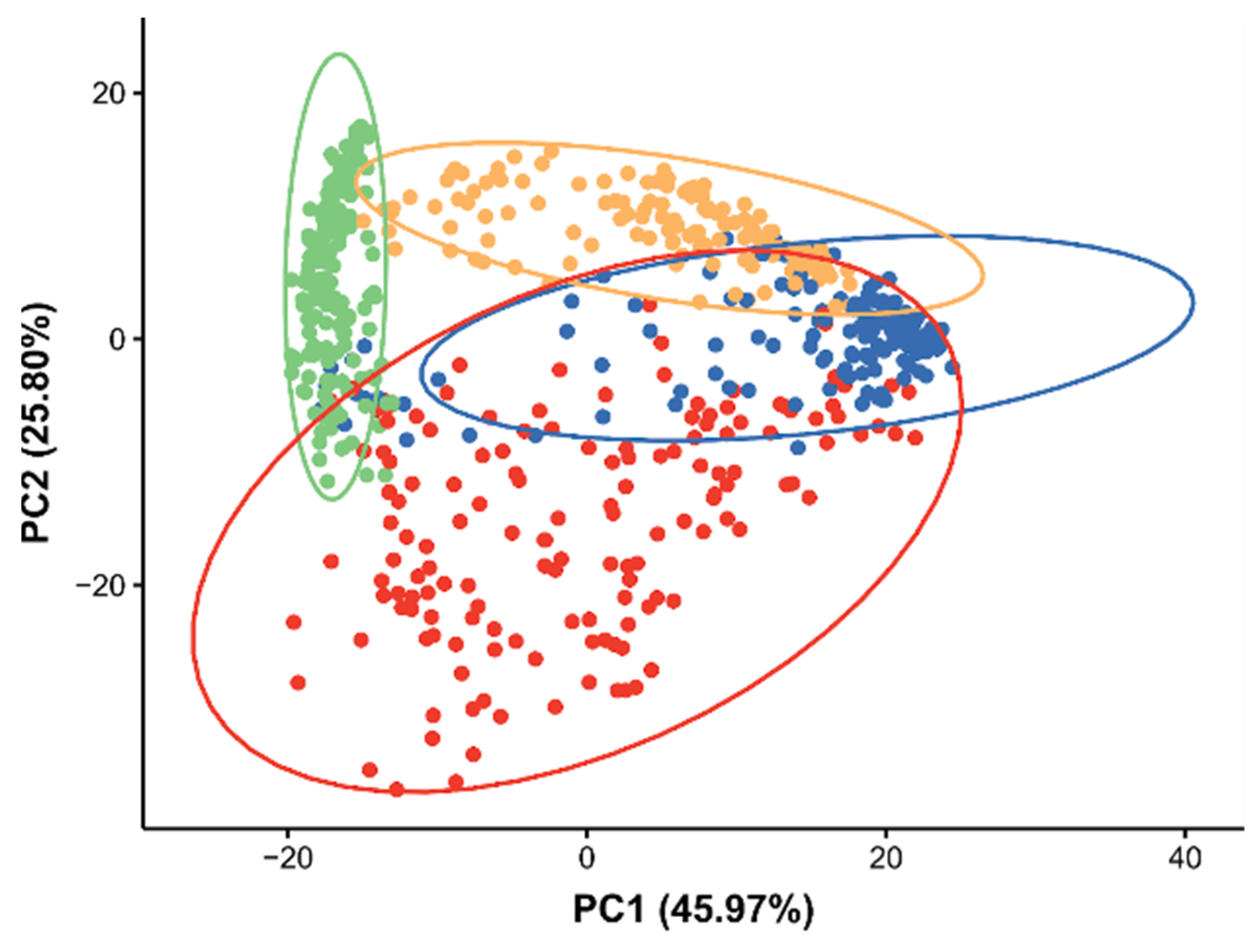
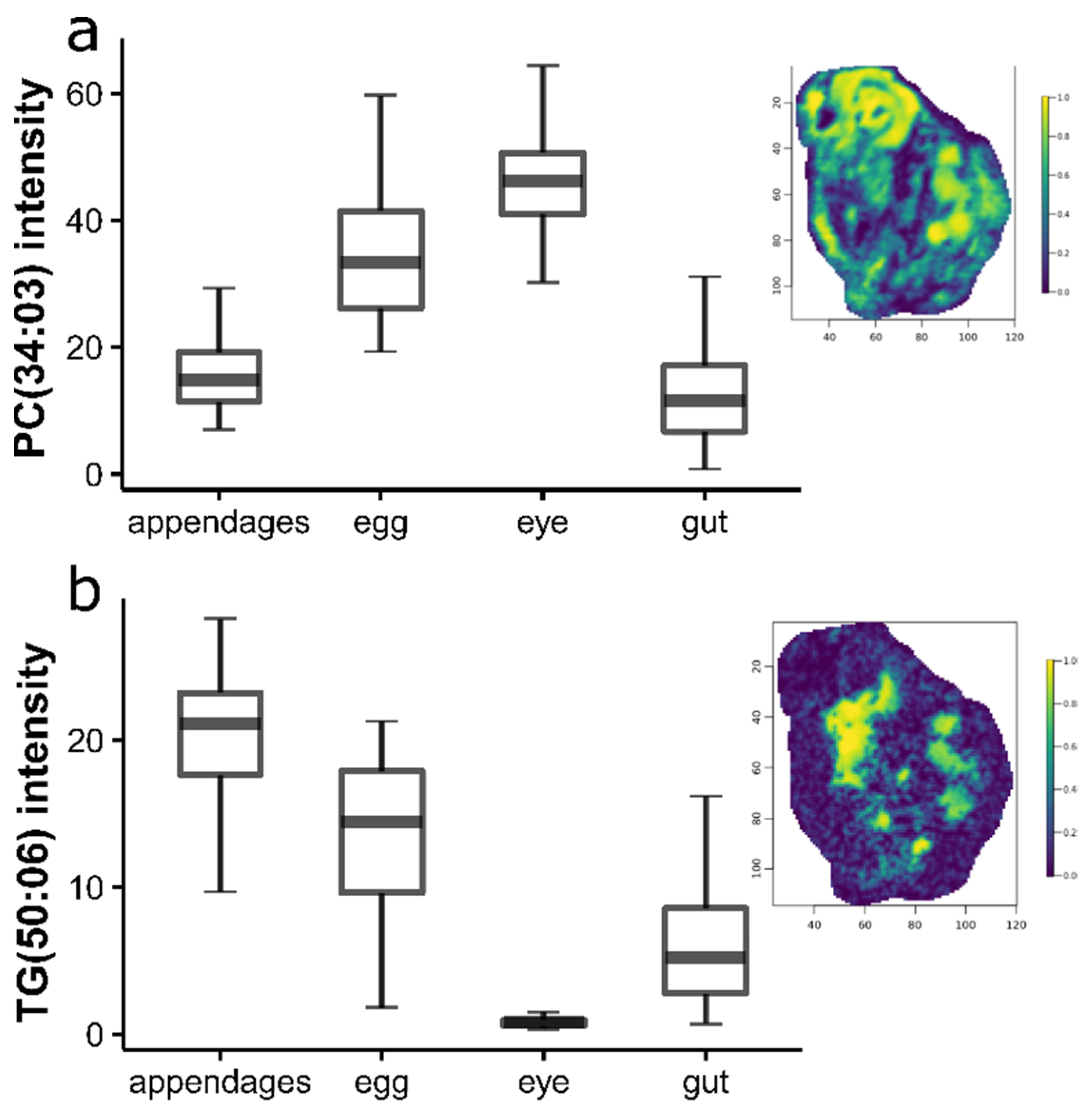
2.2. Lipidomic Differences between Daphnia magna Tissue Types
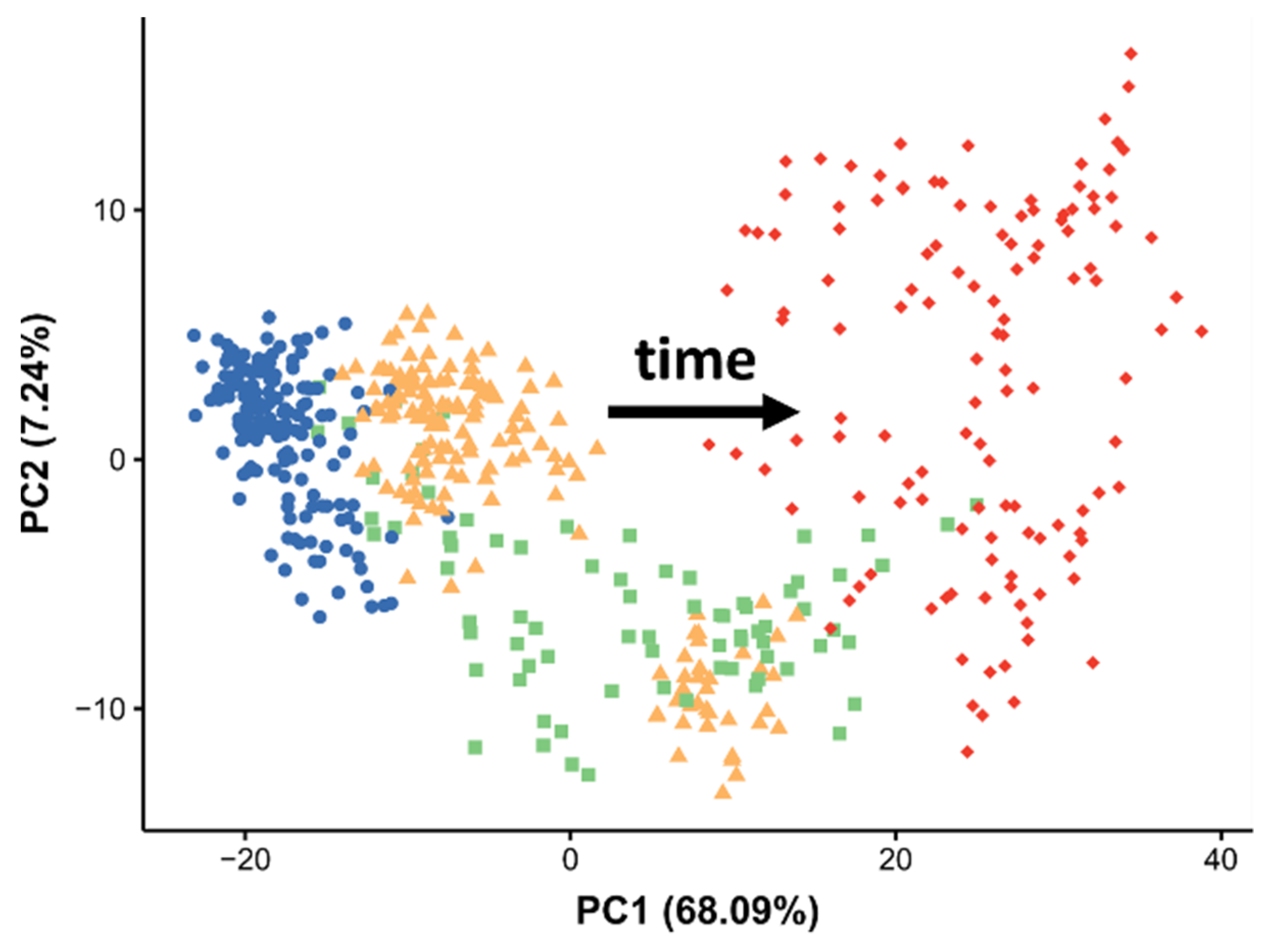
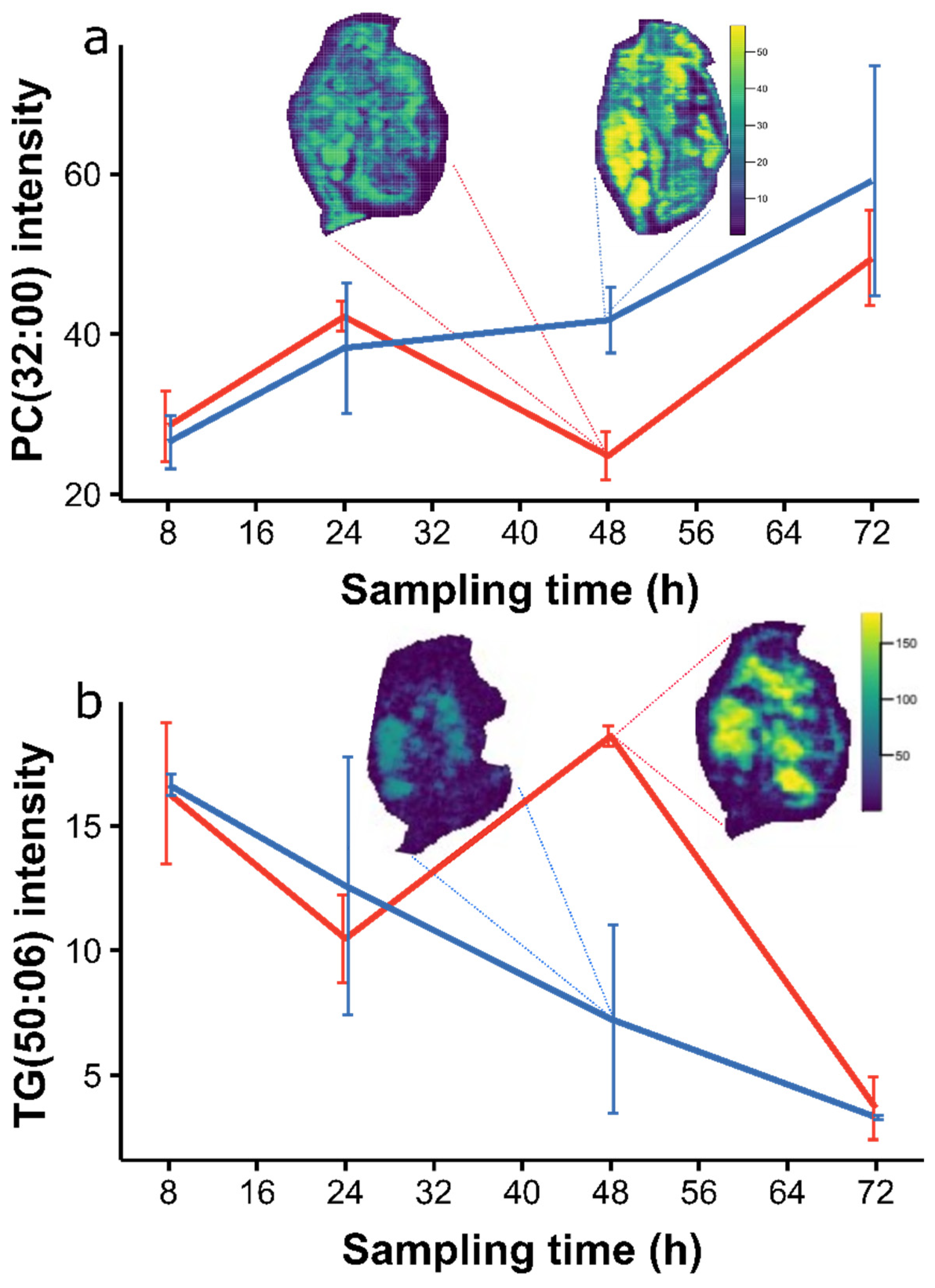
2.3. Temporal Effects in the Baseline Lipidome during Egg Development
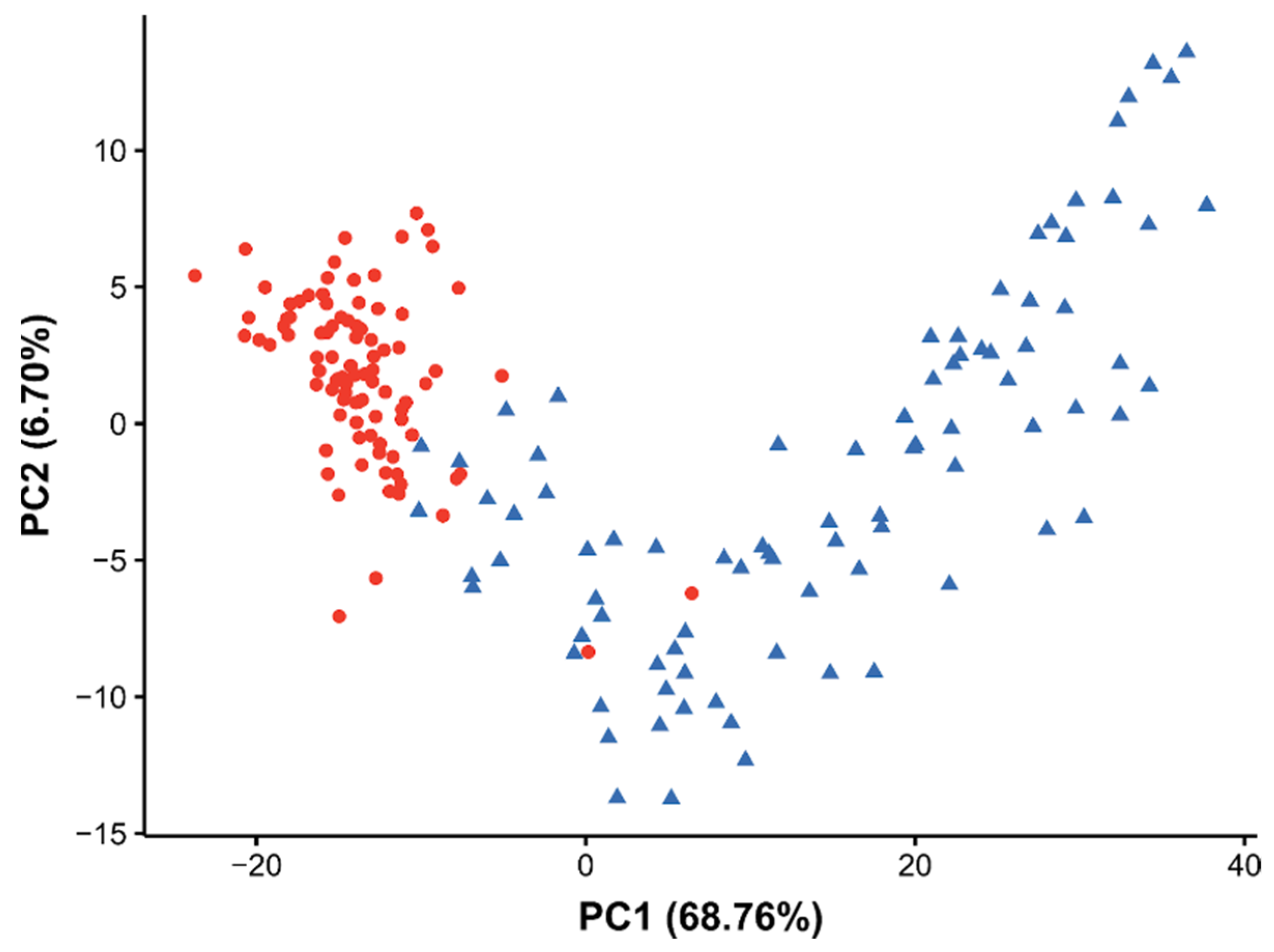
2.4. Perturbation of the Daphnia magna Egg Lipidome to Bisphenol-A
3. Discussion
3.1. Lipidomic Differences between Daphnia magna Tissue Types
3.2. Spatial Lipidome of Daphnia magna
4. Materials and Methods
4.1. Daphnia magna Culturing and Bisphenol-A Exposures
4.2. Preparing Tissue Sections for DESI-MS
4.2.1. Preparing Embedding Media
4.2.2. Cryosectioning Daphnia magna
4.2.3. Preparing Quality Control Samples
4.3. Acquiring DESI-MS Data
4.3.1. Preparing the Desorption Electrospray Ionization—Mass Spectrometer
4.3.2. Analyzing Daphnia magna Study Samples
4.3.3. Quality Control Strategy
4.4. Data Processing and Statistical Analysis of Daphnia magna DESI-MS Data
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sonnhof, U.; Grafe, P.; Krumnikl, J.; Linder, M.; Schindler, L. Test No. 202: Daphnia sp. Acute Immobilisation Test; OECD Guidelines for the Testing of Chemicals, Section 2; OECD: Paris, France, 2004; ISBN 9789264069947. [Google Scholar]
- OECD. Test No. 211: Daphnia Magna Reproduction Test; OECD Guidelines for the Testing of Chemicals, Section 2; OECD: Paris, France, 2012; ISBN 9789264185203. [Google Scholar]
- European Commission. Communication from the Commission to the European Parliament, the Council, the European Economic and Social Committee and the Committee of the Regions; Chemicals Strategy for Sustainability Towards a Toxic-Free Environment: Brussels, Belgium, 2020. [Google Scholar]
- European Commission Chemicals Legislation—Revision of REACH Regulation to Help Achieve a Toxic-Free Environment; European Commission: Brussels, Belgium, 2021.
- Ankley, G.T.; Bennett, R.S.; Erickson, R.J.; Hoff, D.J.; Hornung, M.W.; Johnson, R.D.; Mount, D.R.; Nichols, J.W.; Russom, C.L.; Schmieder, P.K.; et al. Adverse outcome pathways: A conceptual framework to support ecotoxicology research and risk assessment. Environ. Toxicol. Chem. 2010, 29, 730–741. [Google Scholar] [CrossRef]
- Viant, M.R.; Ebbels, T.M.; Beger, R.D.; Ekman, D.R.; Epps, D.J.; Kamp, H.; Leonards, P.E.; Loizou, G.D.; MacRae, J.I.; van Ravenzwaay, B.; et al. Use cases, best practice and reporting standards for metabolomics in regulatory toxicology. Nat. Commun. 2019, 10, 3041. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramirez, T.; Daneshian, M.; Kamp, H.; Bois, F.; Clench, M.; Coen, M.; Donley, B.; Fischer, S.; Ekman, D.; Fabian, E. Metabolomics in toxicology and preclinical research. ALTEX 2013, 30, 209–225. [Google Scholar] [CrossRef] [Green Version]
- Taylor, N.; White, T.; Viant, M. Defining the Baseline and Oxidant Perturbed Lipidomic Profiles of Daphnia magna. Metabolites 2017, 7, 11. [Google Scholar] [CrossRef] [Green Version]
- Jordão, R.; Casas, J.; Fabrias, G.; Campos, B.; Piña, B.; Lemos, M.F.; Soares, A.M.; Tauler, R.; Barata, C. Obesogens beyond vertebrates: Lipid perturbation by tributyltin in the crustacean Daphnia magna. Environ. Health Perspect. 2015, 123, 813–819. [Google Scholar] [CrossRef] [Green Version]
- Fuertes, I.; Barata, C. Characterization of neurotransmitters and related metabolites in Daphnia magna juveniles deficient in serotonin and exposed to neuroactive chemicals that affect its behavior: A targeted LC-MS/MS method. Chemosphere 2021, 263, 127814. [Google Scholar] [CrossRef] [PubMed]
- Wagner, N.D.; Helm, P.A.; Simpson, A.J.; Simpson, M.J. Metabolomic responses to pre-chlorinated and final effluent wastewater with the addition of a sub-lethal persistent contaminant in Daphnia magna. Environ. Sci. Pollut. Res. 2019, 26, 9014–9026. [Google Scholar] [CrossRef]
- Oliveira Pereira, E.A.; Labine, L.M.; Kleywegt, S.; Jobst, K.J.; Simpson, A.J.; Simpson, M.J. Metabolomics Reveals That Bisphenol Pollutants Impair Protein Synthesis-Related Pathways in Daphnia magna. Metabolites 2021, 11, 666. [Google Scholar] [CrossRef] [PubMed]
- Jordão, R.; Garreta, E.; Campos, B.; Lemos, M.F.; Soares, A.M.; Tauler, R.; Barata, C. Compounds altering fat storage in Daphnia magna. Sci. Total Environ. 2016, 545–546, 127–136. [Google Scholar] [CrossRef]
- Chughtai, K.; Heeren, R.M. Mass Spectrometric Imaging for Biomedical Tissue Analysis. Chem. Rev. 2010, 110, 3237–3277. [Google Scholar] [CrossRef] [Green Version]
- Caprioli, R.M.; Farmer, T.B.; Gile, J. Molecular Imaging of Biological Samples: Localization of Peptides and Proteins Using MALDI-TOF MS. Anal. Chem. 1997, 69, 4751–4760. [Google Scholar] [CrossRef]
- Singhal, R.; Carrigan, J.B.; Wei, W.; Taniere, P.; Hejmadi, R.K.; Forde, C.; Ludwig, C.; Bunch, J.; Griffiths, R.L.; Johnson, P.J.; et al. MALDI profiles of proteins and lipids for the rapid characterisation of upper GI-tract cancers. J. Proteom. 2013, 80, 207–215. [Google Scholar] [CrossRef]
- Ščupáková, K.; Soons, Z.; Ertaylan, G.; Pierzchalski, K.A.; Eijkel, G.B.; Ellis, S.R.; Greve, J.W.; Driessen, A.; Verheij, J.; De Kok, T.M.; et al. Spatial Systems Lipidomics Reveals Nonalcoholic Fatty Liver Disease Heterogeneity. Anal. Chem. 2018, 90, 5130–5138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bruinen, A.L.; Fisher, G.L.; Balez, R.; van der Sar, A.M.; Ooi, L.; Heeren, R.M. Identification and High-Resolution Imaging of α-Tocopherol from Human Cells to Whole Animals by TOF-SIMS Tandem Mass Spectrometry. J. Am. Soc. Mass Spectrom. 2018, 29, 1571–1581. [Google Scholar] [CrossRef] [PubMed]
- Hall, Z.; Chu, Y.; Griffin, J.L. Liquid Extraction Surface Analysis Mass Spectrometry Method for Identifying the Presence and Severity of Nonalcoholic Fatty Liver Disease. Anal. Chem. 2017, 89, 5161–5170. [Google Scholar] [CrossRef]
- Hale, O.J.; Cooper, H.J. Native Mass Spectrometry Imaging and In Situ Top-Down Identification of Intact Proteins Directly from Tissue. J. Am. Soc. Mass Spectrom. 2020, 31, 2531–2537. [Google Scholar] [CrossRef] [PubMed]
- Takats, Z.; Wiseman, J.; Gologan, B.; Cooks, R. Mass Spectrometry Sampling under Ambient Conditions with Desorption Electrospray Ionization. Science 2004, 306, 471–473. [Google Scholar] [CrossRef] [Green Version]
- Shrestha, B.; Roman, G.; Hattan, S.; Leveille, W.; Murphy, J.; Michienzi, J.; Musacchio, J.; Jones, E.; Pringle, S.; Innocent, C.; et al. High Spatial Resolution Molecular Tissue Imaging by Desorption Electrospray Ionization using Novel Sprayer Assembly. J. Biomol. Tech. 2020, 31, S17–S18. [Google Scholar]
- Qin, L.; Zhang, Y.; Liu, Y.; He, H.; Han, M.; Li, Y.; Zeng, M.; Wang, X. Recent advances in matrix-assisted laser desorption/ionisation mass spectrometry imaging (MALDI-MSI) for in situ analysis of endogenous molecules in plants. Phytochem. Anal. 2018, 29, 351–364. [Google Scholar] [CrossRef]
- Perez, C.J.; Tata, A.; de Campos, M.L.; Peng, C.; Ifa, D.R. Monitoring Toxic Ionic Liquids in Zebrafish (Danio rerio) with Desorption Electrospray Ionization Mass Spectrometry Imaging (DESI-MSI). J. Am. Soc. Mass Spectrom. 2017, 28, 1136–1148. [Google Scholar] [CrossRef] [Green Version]
- Pirro, V.; Guffey, S.C.; Sepúlveda, M.S.; Mahapatra, C.T.; Ferreira, C.R.; Jarmusch, A.K.; Cooks, R.G. Lipid dynamics in zebrafish embryonic development observed by DESI-MS imaging and nanoelectrospray-MS. Mol. Biosyst. 2016, 12, 2069–2079. [Google Scholar] [CrossRef]
- Niehoff, A.C.; Kettling, H.; Pirkl, A.; Chiang, Y.N.; Dreisewerd, K.; Yew, J.Y. Analysis of Drosophila Lipids by Matrix-Assisted Laser Desorption/Ionization Mass Spectrometric Imaging. Anal. Chem. 2014, 86, 11086–11092. [Google Scholar] [CrossRef]
- Liu, W.; Nie, H.; Liang, D.; Bai, Y.; Liu, H. Phospholipid imaging of zebrafish exposed to fipronil using atmospheric pressure matrix-assisted laser desorption ionization mass spectrometry. Talanta 2020, 209, 120357. [Google Scholar] [CrossRef] [PubMed]
- Böhme, S.; Stärk, H.J.; Kühnel, D.; Reemtsma, T. Exploring LA-ICP-MS as a quantitative imaging technique to study nanoparticle uptake in Daphnia magna and zebrafish (Danio rerio) embryos. Anal. Bioanal. Chem. 2015, 407, 5477–5485. [Google Scholar] [CrossRef] [Green Version]
- Ishida, Y.; Nakanishi, O.; Hirao, S.; Tsuge, S.; Urabe, J.; Sekino, T.; Nakanishi, M.; Kimoto, T.; Ohtani, H. Direct Analysis of Lipids in Single Zooplankter Individuals by Matrix-Assisted Laser Desorption/Ionization Mass Spectrometry. Anal. Chem. 2003, 75, 4514–4518. [Google Scholar] [CrossRef]
- Mu, X.; Rider, C.V.; Gap, S.H.; Hoy, H.; LeBlanc, G.A. Covert signal disruption: Anti-ecdysteroidal activity of bisphenol A involves cross talk between signaling pathways. Environ. Toxicol. Chem. 2005, 24, 146–152. [Google Scholar] [CrossRef] [PubMed]
- Fuertes, I.; Jordão, R.; Casas, F.; Barata, C. Allocation of glycerolipids and glycerophospholipids from adults to eggs in Daphnia magna: Perturbations by compounds that enhance lipid droplet accumulation. Environ. Pollut. 2018, 242, 1702–1710. [Google Scholar] [CrossRef]
- Schwartz, S.A.; Reyzer, M.L.; Caprioli, R.M. Direct tissue analysis using matrix-assisted laser desorption/ionization mass spectrometry: Practical aspects of sample preparation. J. Mass Spectrom. 2003, 38, 699–708. [Google Scholar] [CrossRef]
- Nelson, K.A.; Daniels, G.J.; Fournie, J.W.; Hemmer, M.J. Optimization of Whole-Body Zebrafish Sectioning Methods for Mass Spectrometry Imaging. J. Biomol. Tech. 2013, 24, 119–127. [Google Scholar] [CrossRef]
- Chramow, A.; Hamid, T.S.; Eberlin, L.S.; Girod, M.; Ifa, D.R. Imaging of whole zebra fish (Danio rerio) by desorption electrospray ionization mass spectrometry. Rapid Commun. Mass Spectrom. 2014, 28, 2084–2088. [Google Scholar] [CrossRef] [PubMed]
- Khalil, S.M.; Römpp, A.; Pretzel, J.; Becker, K.; Spengler, B. Phospholipid Topography of Whole-Body Sections of the Anopheles stephensi Mosquito, Characterized by High-Resolution Atmospheric-Pressure Scanning Microprobe Matrix-Assisted Laser Desorption/Ionization Mass Spectrometry Imaging. Anal. Chem. 2015, 87, 11309–11316. [Google Scholar] [CrossRef] [Green Version]
- Tanasawa, I. Things We Do Not Know about Cryopreservation of Biological Organs. Ann. N. Y. Acad. Sci. 1998, 858, 227–234. [Google Scholar] [CrossRef]
- Claude, E.; Jones, E.A.; Pringle, S.D. DESI Mass Spectrometry Imaging (MSI). In Imaging Mass Spectrometry Methods and Protocols; Springer Protocols: Hertfordshire, UK, 2017; Volume 26, pp. 65–75. ISBN 9781493970506. [Google Scholar]
- Bemis, K.D.; Harry, A.; Eberlin, L.S.; Ferreira, C.; van de Ven, S.M.; Mallick, P.; Stolowitz, M.; Vitek, O. Cardinal: An R package for statistical analysis of mass spectrometry-based imaging experiments. Bioinformatics 2015, 31, 2418–2420. [Google Scholar] [CrossRef] [Green Version]
- Currie, L.A. Limits for qualitative detection and quantitative determination. Application to radiochemistry. Anal. Chem. 1968, 40, 586–593. [Google Scholar] [CrossRef]
- Committee for Risk Assessment RAC-ECHA. Background Document to the Opinion Proposing Harmonised Classification and Labelling at Community Level of4,4’-isopropylidenediphenol; bisphenol A. Dortmund, Germany. 2020. Available online: https://echa.europa.eu/documents/10162/b8a9b144-33c0-064f-bedc-39032a59e0dc (accessed on 18 December 2021).
- Zemaitis, K.J.; Izydorczak, A.M.; Thompson, A.C.; Wood, T.D. Streamlined Multimodal DESI and MALDI Mass Spectrometry Imaging on a Singular Dual-Source FT-ICR Mass Spectrometer. Metabolites 2021, 11, 253. [Google Scholar] [CrossRef] [PubMed]
- Palmer, A.; Phapale, P.; Chernyavsky, I.; Lavigne, R.; Fay, D.; Tarasov, A.; Kovalev, V.; Fuchser, J.; Nikolenko, S.; Pineau, C.; et al. FDR-controlled metabolite annotation for high-resolution imaging mass spectrometry. Nat. Methods 2017, 14, 57–60. [Google Scholar] [CrossRef]
- Shrestha, B.; Towers, M.; Olivos, H.; Midey, A.; Claude, E. Lipidomic Imaging of Isobaric Lipids Using High-Resolution Ion Mobility Mass Spectrometry with DESI XS. Waters. 2021. Available online: https://www.waters.com/waters/library.htm?locale=en_US&lid=135085365 (accessed on 18 December 2021).
- Claes, B.S.; Bowman, A.P.; Poad, B.L.; Young, R.S.; Heeren, R.M.; Blanksby, S.J.; Ellis, S.R. Mass Spectrometry Imaging of Lipids with Isomer Resolution Using High-Pressure Ozone-Induced Dissociation. Anal. Chem. 2021, 93, 9826–9834. [Google Scholar] [CrossRef] [PubMed]
- Sumner, L.W.; Amberg, A.; Barrett, D.; Beale, M.H.; Beger, R.; Daykin, C.A.; Fan, T.W.; Fiehn, O.; Goodacre, R.; Griffin, J.L.; et al. Proposed minimum reporting standards for chemical analysis Chemical Analysis Working Group (CAWG) Metabolomics Standards Initiative (MSI) NIH Public Access. Metabolomics 2007, 3, 211–221. [Google Scholar] [CrossRef] [Green Version]
- Viant, M.R.; Kurland, I.J.; Jones, M.R.; Dunn, W.B. How close are we to complete annotation of metabolomes? Curr. Opin. Chem. Biol. 2017, 36, 64–69. [Google Scholar] [CrossRef]
- Brennan, S.J.; Brougham, C.A.; Roche, J.J.; Fogarty, A.M. Multi-generational effects of four selected environmental oestrogens on Daphnia magna. Chemosphere 2006, 64, 49–55. [Google Scholar] [CrossRef] [PubMed]
- Taylor, N.S.; Weber, R.J.; Southam, A.D.; Payne, T.G.; Hrydziuszko, O.; Arvanitis, T.N.; Viant, M.R. A new approach to toxicity testing in Daphnia magna: Application of high throughput FT-ICR mass spectrometry metabolomics. Metabolomics 2009, 5, 44–58. [Google Scholar] [CrossRef]
- Schramm, T.; Hester, A.; Klinkert, I.; Both, J.P.; Heeren, R.M.; Brunelle, A.; Laprévote, O.; Desbenoit, N.; Robbe, M.F.; Stoeckli, M.; et al. imzML—A common data format for the flexible exchange and processing of mass spectrometry imaging data. J. Proteom. 2012, 75, 5106–5110. [Google Scholar] [CrossRef] [PubMed]
- Benjamini, Y.; Hochberg, Y. Controlling the False Discovery Rate: A Practical and Powerful Approach to Multiple Testing. J. R. Stat. Soc. 1995, 57, 289–300. [Google Scholar] [CrossRef]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Smith, M.J.; Weber, R.J.M.; Viant, M.R. Spatially Mapping the Baseline and Bisphenol-A Exposed Daphnia magna Lipidome Using Desorption Electrospray Ionization—Mass Spectrometry. Metabolites 2022, 12, 33. https://doi.org/10.3390/metabo12010033
Smith MJ, Weber RJM, Viant MR. Spatially Mapping the Baseline and Bisphenol-A Exposed Daphnia magna Lipidome Using Desorption Electrospray Ionization—Mass Spectrometry. Metabolites. 2022; 12(1):33. https://doi.org/10.3390/metabo12010033
Chicago/Turabian StyleSmith, Matthew J., Ralf J. M. Weber, and Mark R. Viant. 2022. "Spatially Mapping the Baseline and Bisphenol-A Exposed Daphnia magna Lipidome Using Desorption Electrospray Ionization—Mass Spectrometry" Metabolites 12, no. 1: 33. https://doi.org/10.3390/metabo12010033
APA StyleSmith, M. J., Weber, R. J. M., & Viant, M. R. (2022). Spatially Mapping the Baseline and Bisphenol-A Exposed Daphnia magna Lipidome Using Desorption Electrospray Ionization—Mass Spectrometry. Metabolites, 12(1), 33. https://doi.org/10.3390/metabo12010033