
Characterization of Phytoestrogens in Medicago sativa L. and Grazing Beef Cattle

 ,
, 
 and
and
Abstract
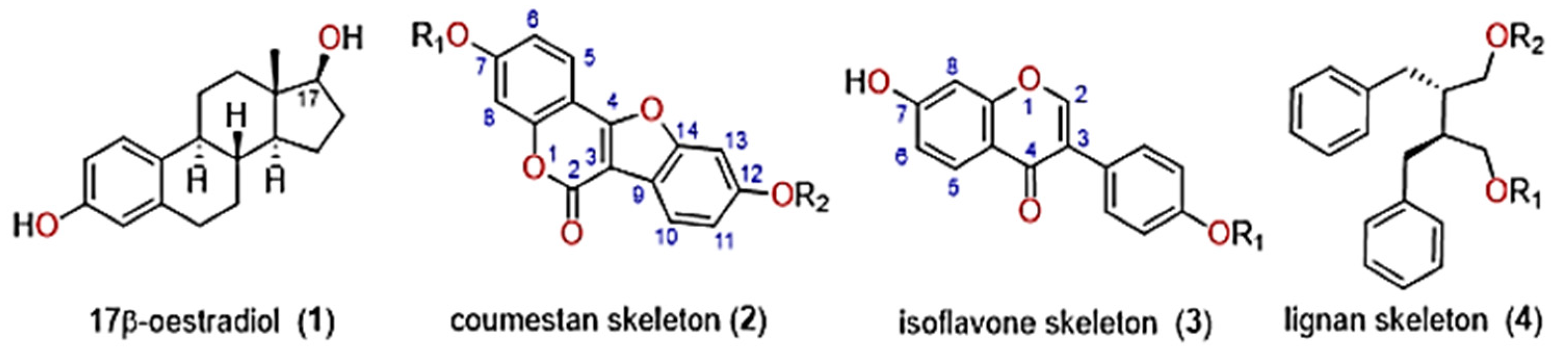
:1. Introduction
2. Results
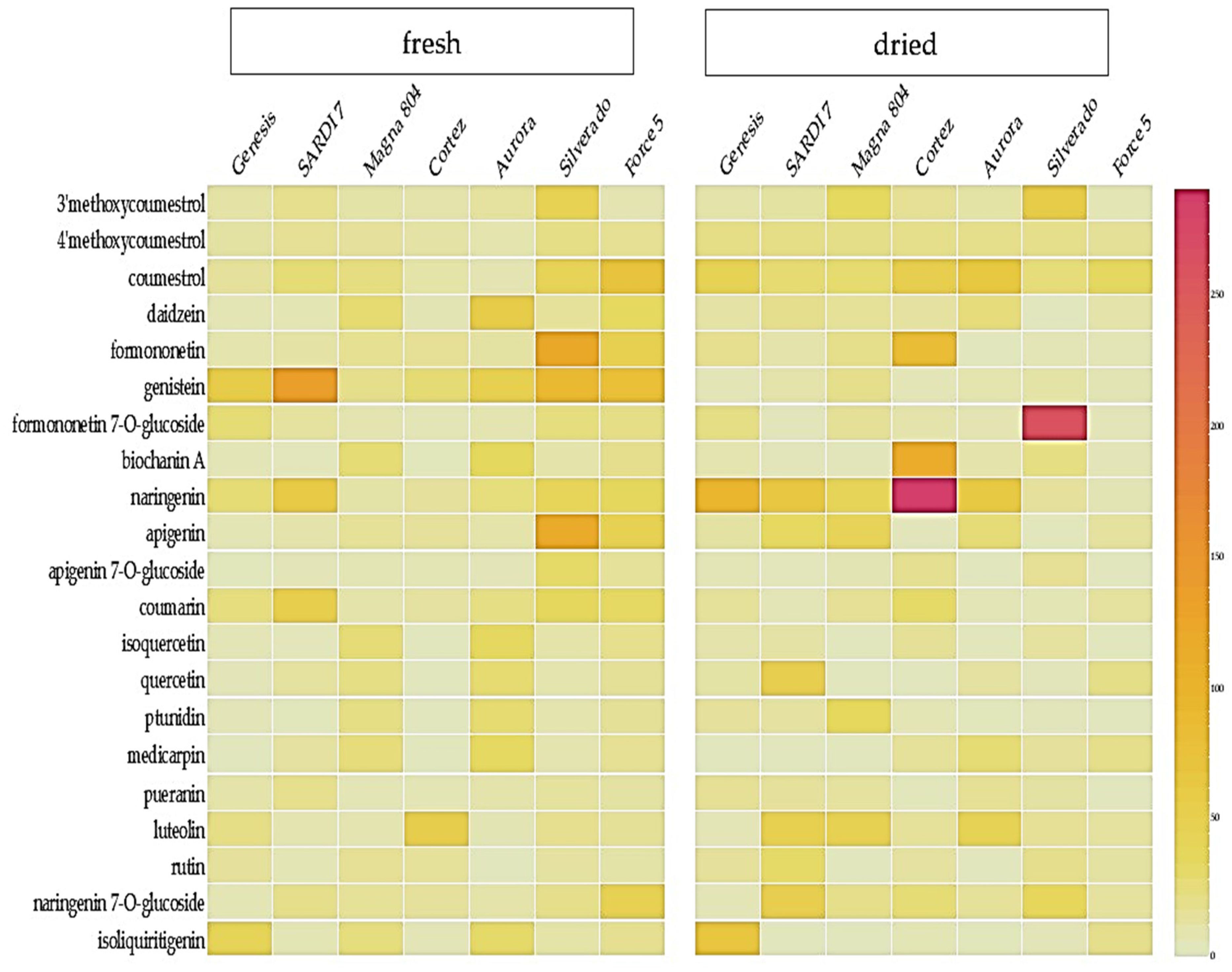
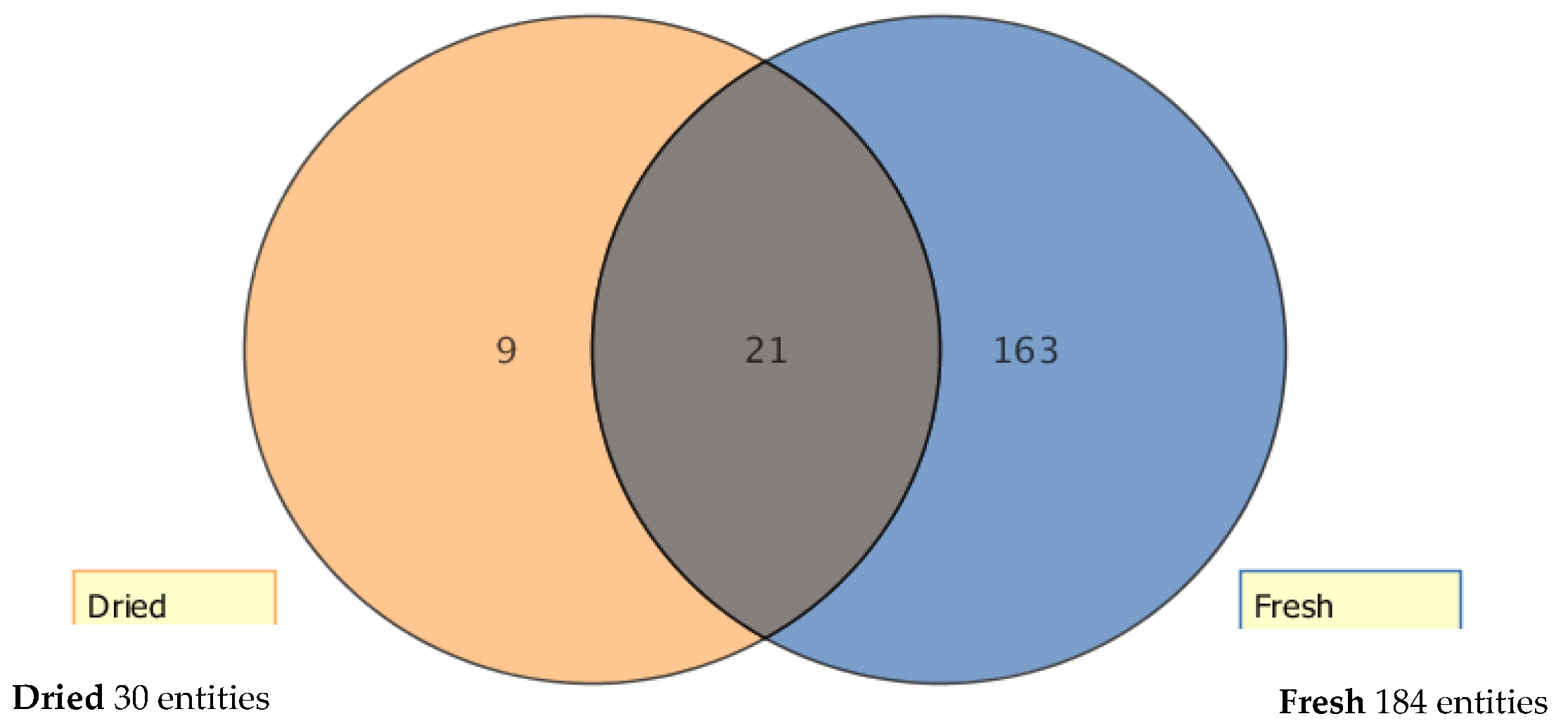
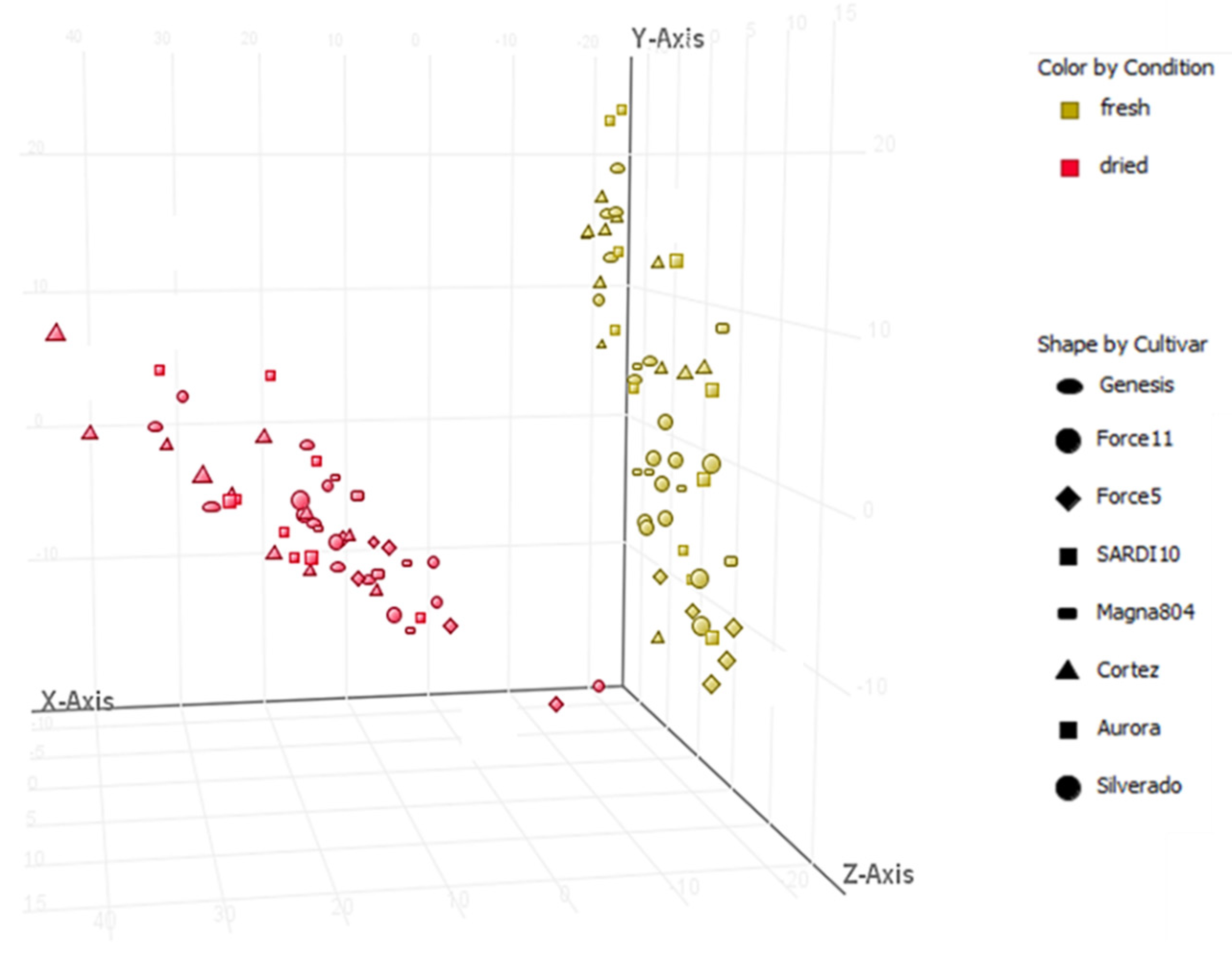
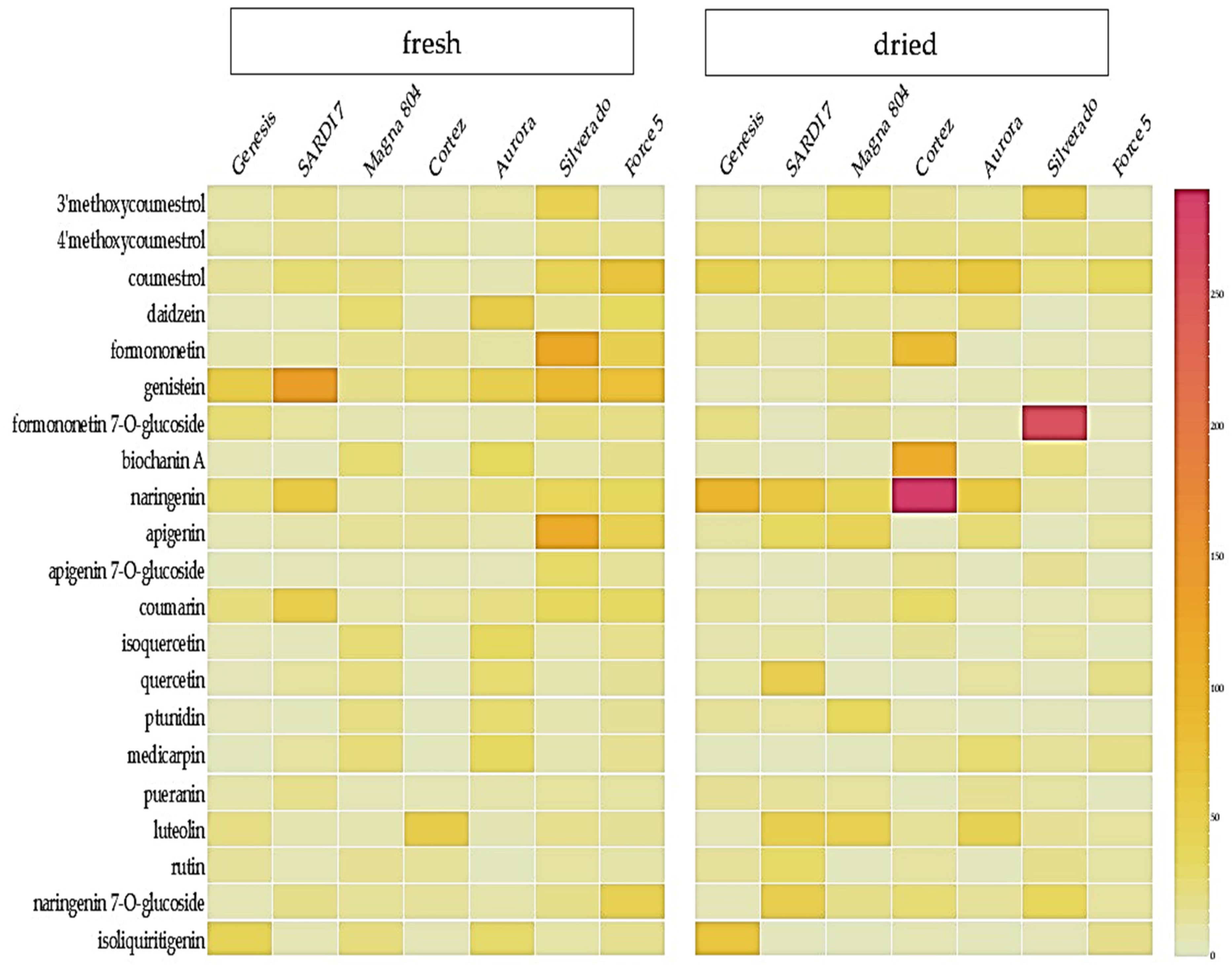
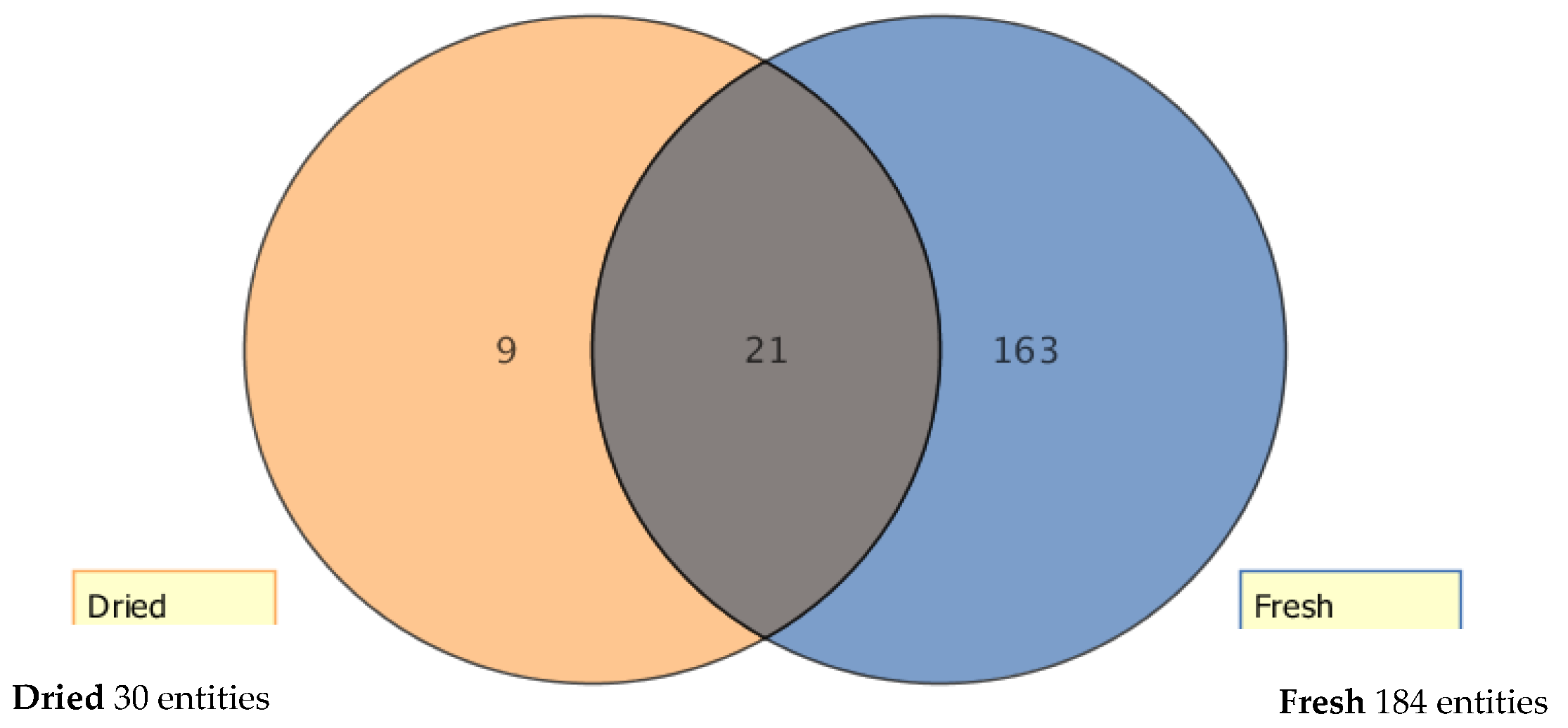
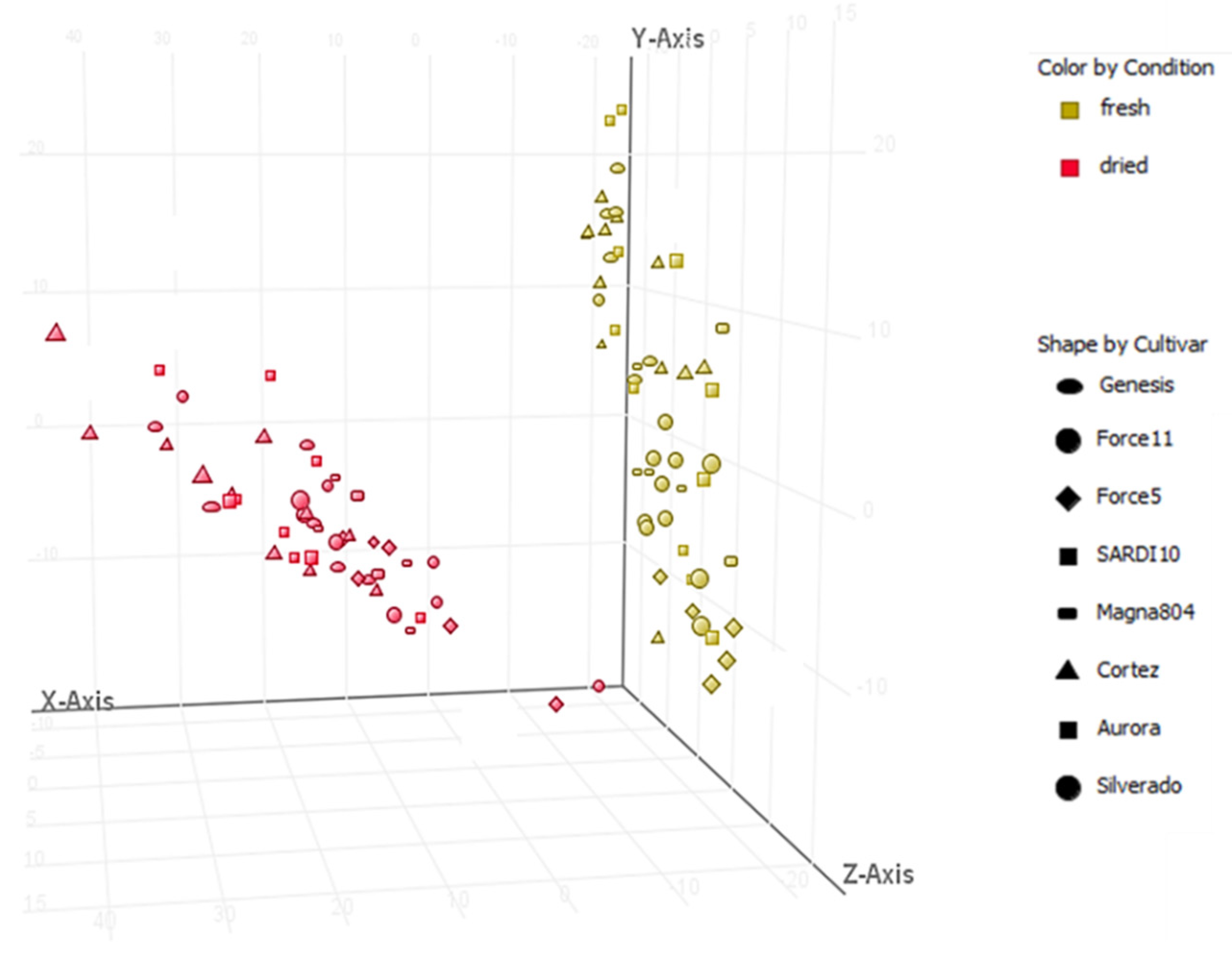
2.1. Metabolic Profiling of Selected Lucerne Cultivars
2.2. Quantification of Phytoestrogens in Lucerne Cultivars
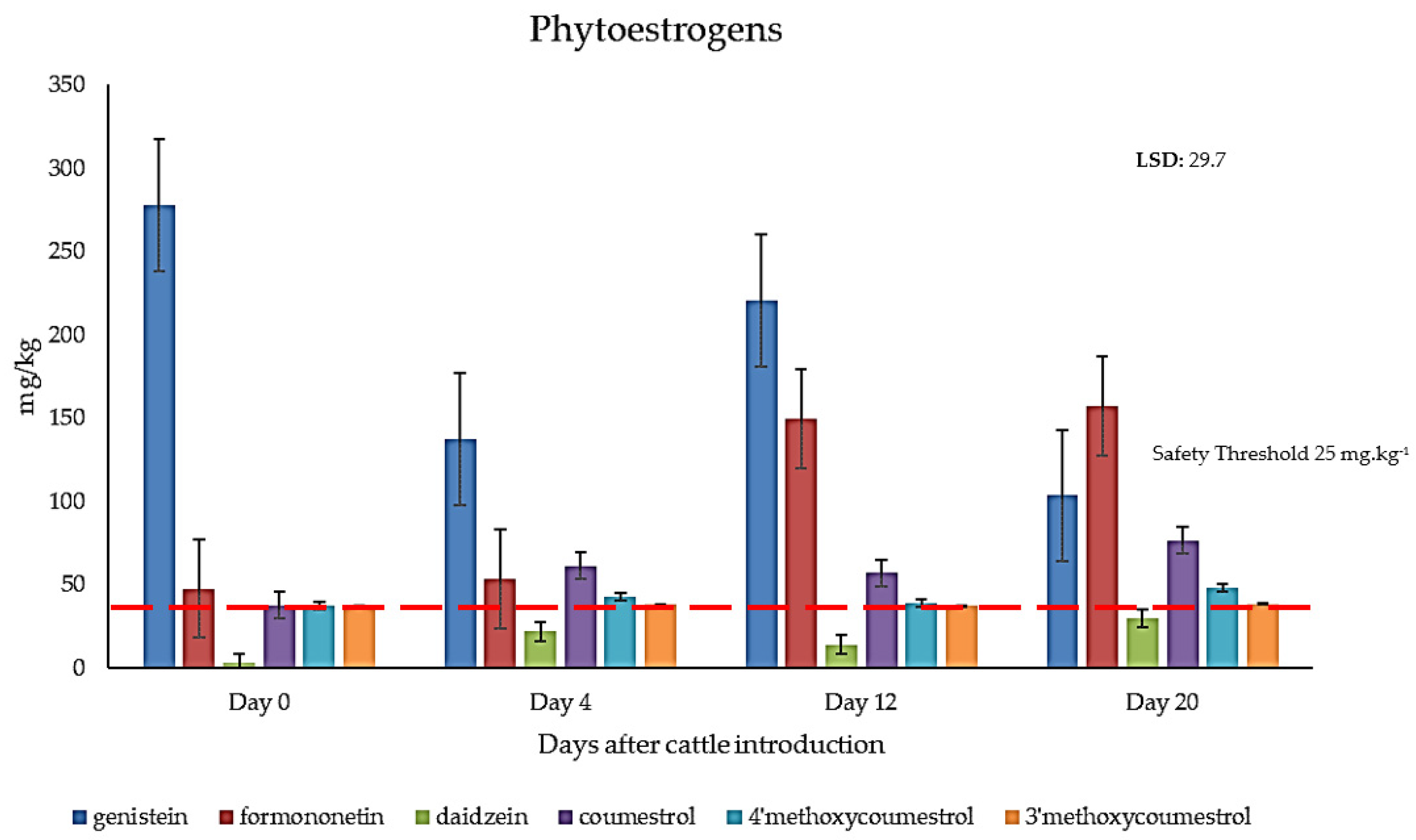
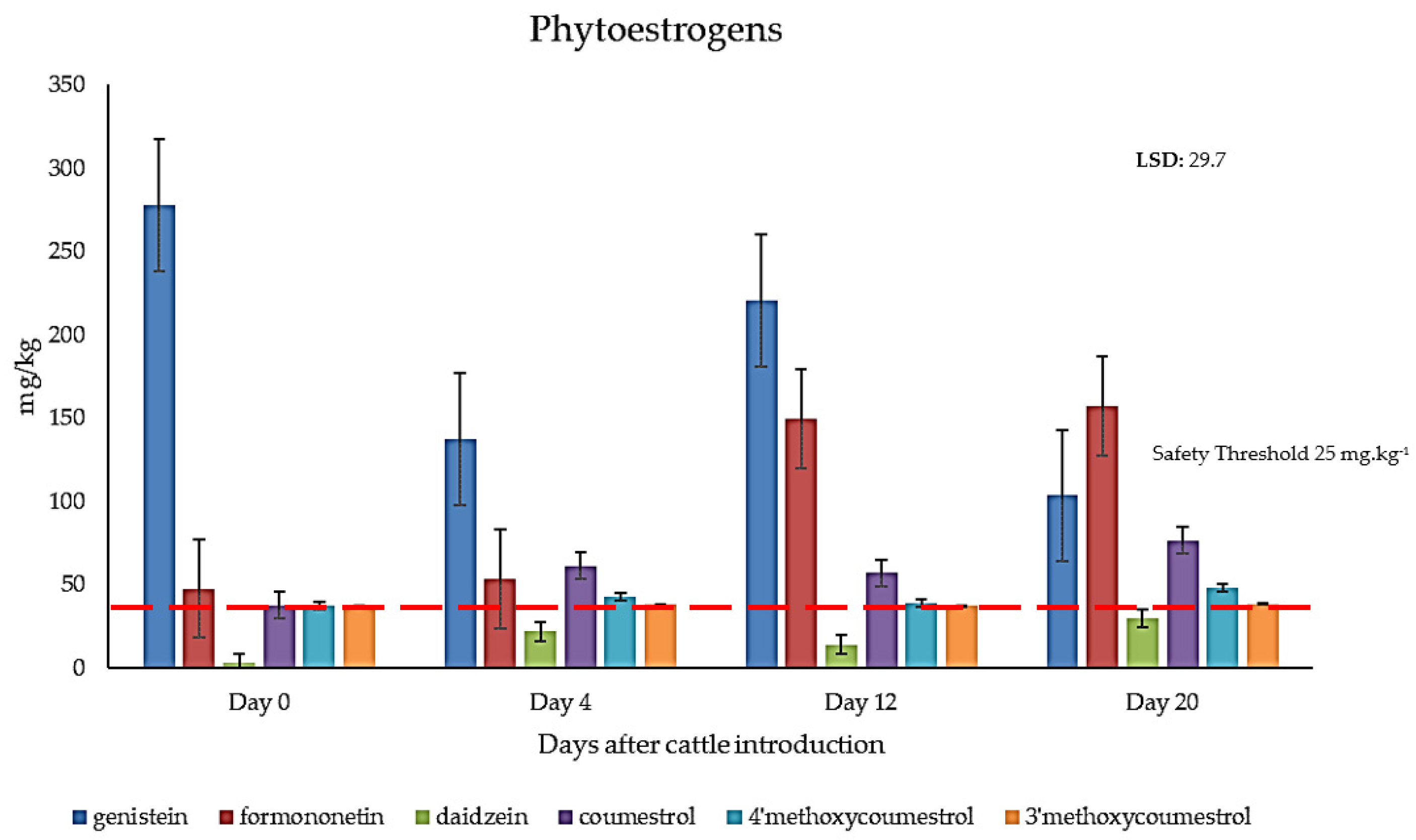
2.3. Quantification of Phytoestrogens in Field-Grown Genesis Lucerne
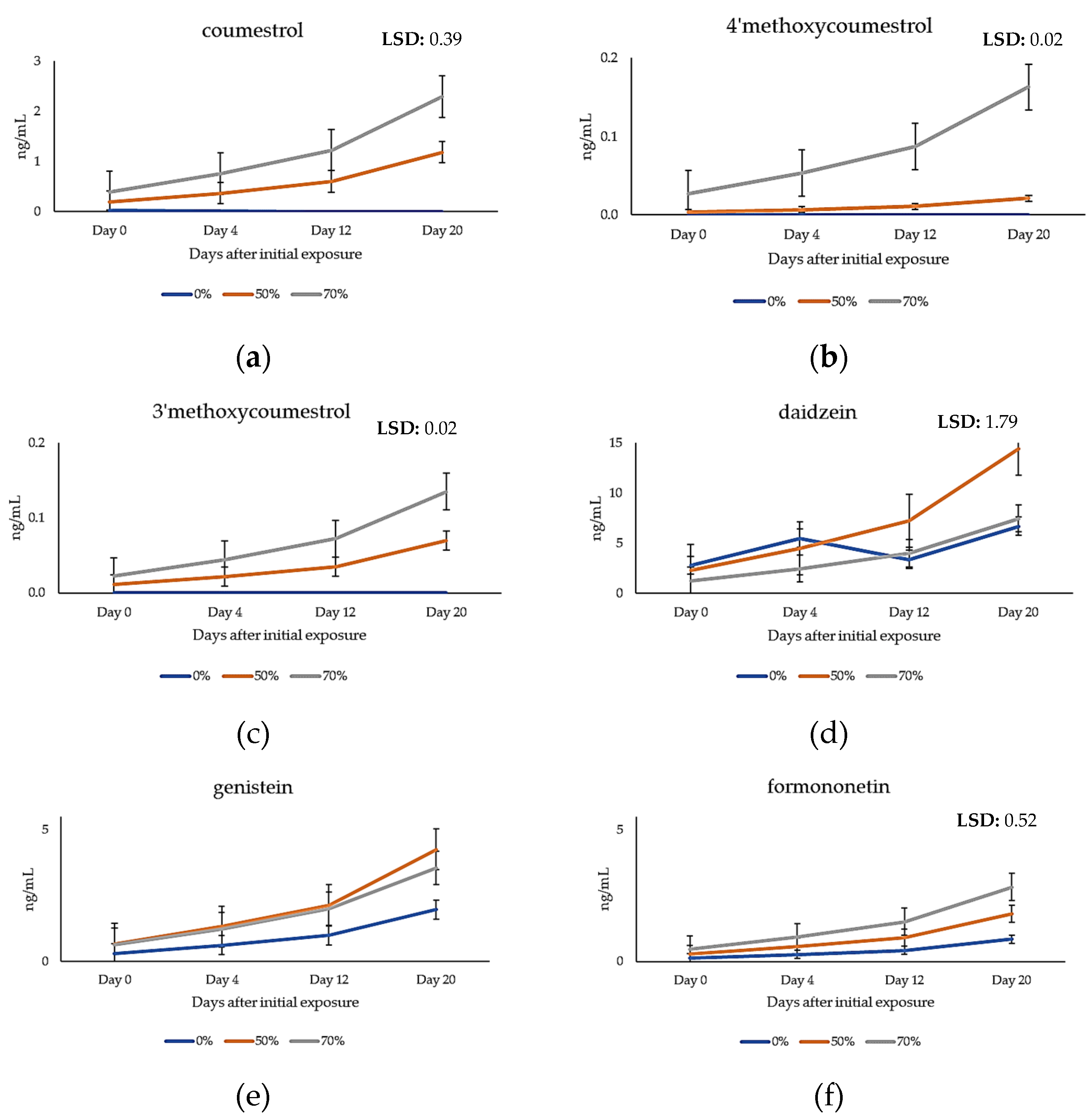
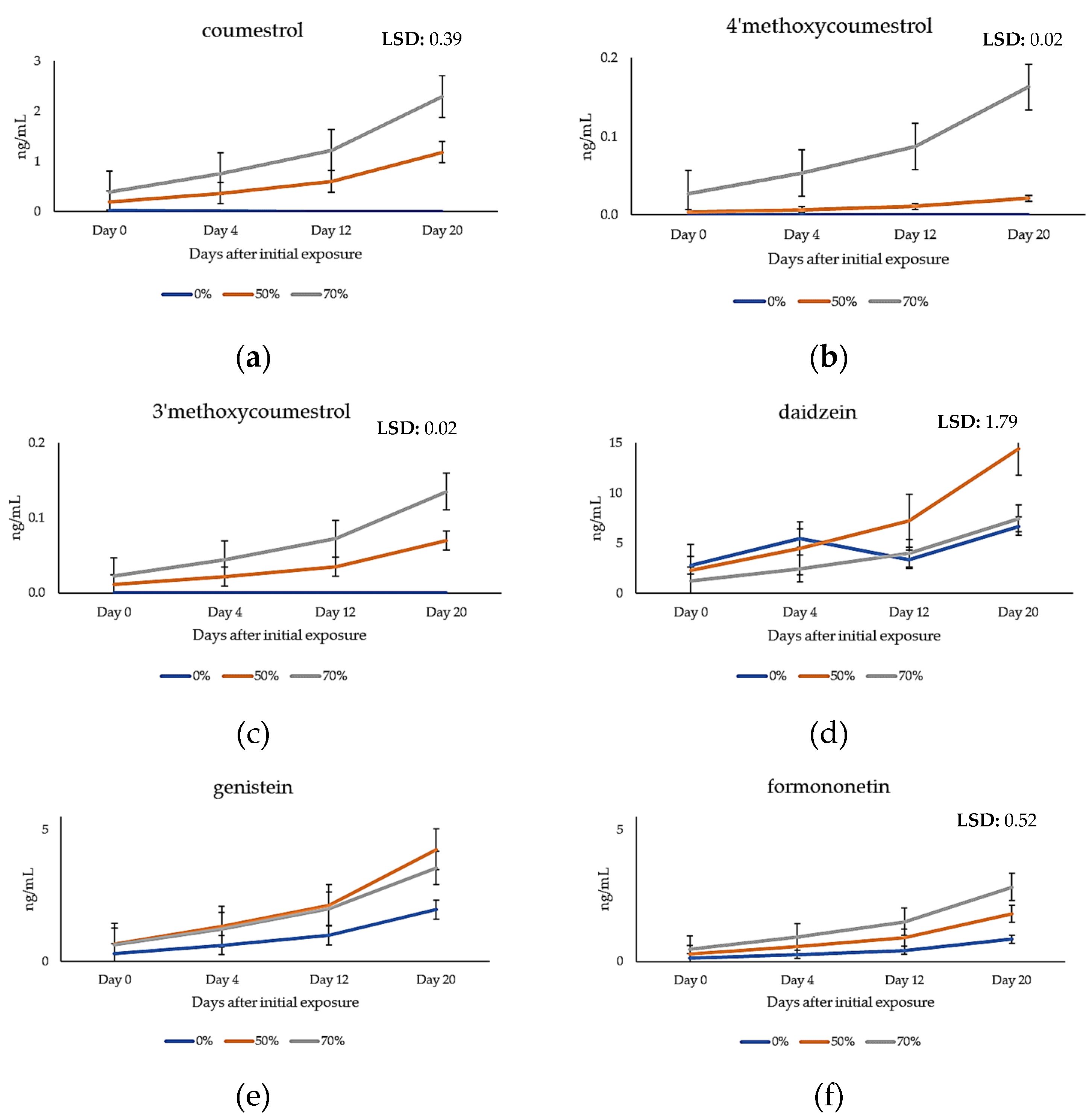
2.4. Quantification of Phytoestrogens in Plasma Samples of Grazing Cattle
3. Discussion
3.1. Metabolic Profiling of Key Phytoestrogens in Lucerne Cultivars
3.2. Metabolic Profiling of Key Phytoestrogens in Genesis Lucerne and Cattle Plasma
3.3. Metabolism of Phytoestrogens
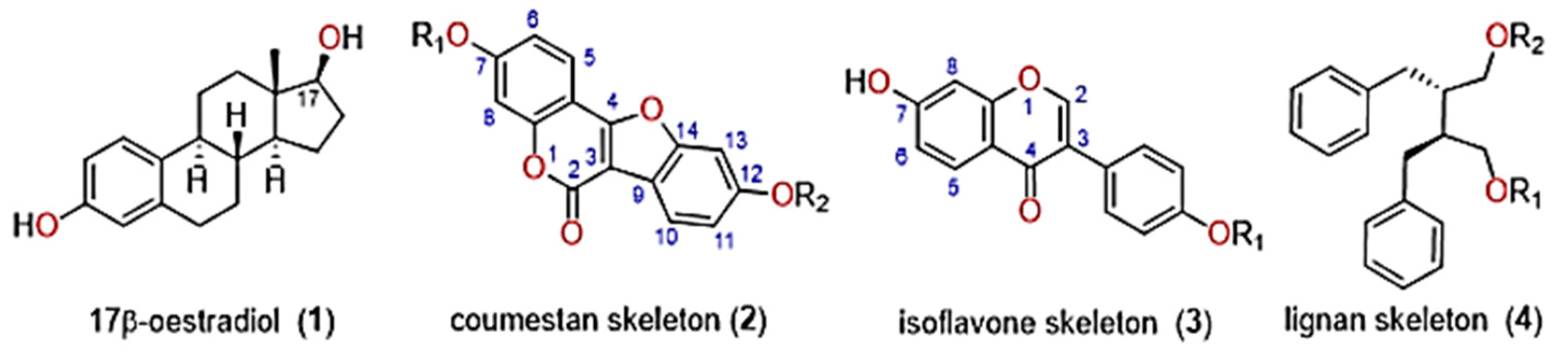
3.4. Biosynthesis of Phytoestrogens
4. Materials and Methods
4.1. Greenhouse Trial
4.2. Plant Tissue Extraction
4.3. Cattle Grazing Trial
4.4. Blood Sampling and Extractions
4.5. Metabolic Profiling of Phytoestrogens
4.6. Data Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tucak, M.; Čupić, T.; Horvat, D.; Popović, S.; Krizmanić, G.; Ravlić, M. Variation of Phytoestrogen Content and Major Agronomic Traits in Alfalfa (Medicago sativa L.) Populations. Agronomy 2020, 10, 87. [Google Scholar] [CrossRef] [Green Version]
- Puschner, B. Anti-nutritional factors in alfalfa hay. In Proceedings of the 29th National Alfalfa Symposium, Las Vegas, NV, USA, 10–12 December 2000; The California Alfalfa Workgroup and The Alfalfa Council: Las Vegas, NV, USA, 2000; Volume 30, pp. 157–162. [Google Scholar]
- Reed, K.F.M. Fertility of herbivores consuming phytoestrogen-containing Medicago and Trifolium species. Agriculture 2016, 6, 35. [Google Scholar] [CrossRef] [Green Version]
- Revell, C.K.; Ewing, M.A.; Real, D.; Nichols, P.G.H.; Sandral, G.A. Commercialisation and impacts of pasture legumes in southern Australia–lessons learnt. In Proceedings of the 22nd International Grassland Congress, Sydney, Australia, 15–19 September 2013; New South Wales Department of Primary Industry: Sydney, Australia, 2013; Volume 22, Volume 286, p. 282. [Google Scholar]
- Patisaul, H.B.; Jefferson, W. The pros and cons of phytoestrogens. Front. Neuroendocr. 2010, 31, 400–419. [Google Scholar] [CrossRef] [Green Version]
- Dixon, R.A. Phytoestrogens. Annu. Rev. Plant Biol. 2004, 55, 225–261. [Google Scholar] [CrossRef] [Green Version]
- Hashem, N.; Soltan, Y. Impacts of phytoestrogens on livestock production: A review. Egypt. J. Nutri. Feeds. 2016, 19, 81–89. [Google Scholar] [CrossRef] [Green Version]
- Křížová, L.; Dadáková, K.; Kašparovská, J.; Kašparovský, T. Isoflavones. Molecules 2019, 24, 1076. [Google Scholar] [CrossRef] [Green Version]
- Ferreira-Dias, G.; Botelho, M.; Zagrajczuk, A.; Rebordão, M.R.; Galvão, A.M.; Pinto Bravo, P.; Piotrowska-Tomala, K.; Szóstek, A.Z.; Wiczkowski, W.; Piskula, M.; et al. Coumestrol and its metabolite in mares’ plasma after ingestion of phytoestrogen-rich plants: Potent endocrine disruptors inducing infertility. Theriogenology 2013, 80, 684–692. [Google Scholar] [CrossRef]
- Hashem, N.; El-Azrak, K.; El-Din, A.N.; Sallam, S.; Taha, T.; Salem, M. Effects of Trifolium alexandrinum phytoestrogens on oestrous behaviour, ovarian activity and reproductive performance of ewes during the non-breeding season. Anim. Reprod. Sci. 2018, 196, 1–8. [Google Scholar] [CrossRef]
- Hughes, C.L. Phytochemical mimicry of reproductive hormones and modulation of herbivore fertility by phytoestrogens. Environ. Health Persp. 1988, 78, 171–174. [Google Scholar] [CrossRef]
- Wocławek-Potocka, I.C.; Mannelli, D.; Boruszewska, D.; Kowalczyk-Zieba, I.; Waśniewski, T.; Skarżyński, D.J. Diverse effects of phytoestrogens on the reproductive performance: Cow as a model. Int. J. Endocrinol 2013, 2013, 650984. [Google Scholar] [CrossRef] [Green Version]
- Bennetts, H.; Underwood, E.; Shier, F.L. A specific breeding problem of sheep on subterranean clover pastures in Western Australia. Aust. Vet. J. 1946, 22, 2–12. [Google Scholar] [CrossRef]
- Latif, S.; Weston, P.A.; Barrow, R.A.; Gurusinghe, S.; Piltz, J.W.; Weston, L.A. Metabolic profiling provides unique insights to accumulation and biosynthesis of key secondary metabolites in annual pasture legumes of Mediterranean origin. Metabolites 2020, 10, 267. [Google Scholar] [CrossRef] [PubMed]
- Blache, D.; Maloney, S.K.; Revell, D.K. Use and limitations of alternative feed resources to sustain and improve reproductive performance in sheep and goats. Anim. Feed Sci. Tech. 2008, 147, 140–157. [Google Scholar] [CrossRef]
- Bora, K.S.; Sharma, A. Phytochemical and pharmacological potential of Medicago sativa: A review. Pharm. Biol. 2011, 49, 211–220. [Google Scholar] [CrossRef] [PubMed]
- Jefferson, W.N.; Patisaul, H.B.; Williams, C.J. Reproductive consequences of developmental phytoestrogen exposure. Reproduction 2012, 143, 247. [Google Scholar] [CrossRef]
- Madej, A.; Lundh, T. Risk of adverse effects of phytoestrogens in animal feed. In Bioactive Compounds in Plants–Benefits and Risks for Man and Animals; Bernhoft, A., Ed.; The Norwegian Academy of Science and Letters: Oslo, Norway, 2008; pp. 94–103. [Google Scholar]
- Phillips, D.A.; Kapulnik, P. Plant isoflavonoids, pathogens and symbionts. Trends Microbiol. 1995, 3, 58–64. [Google Scholar] [CrossRef]
- Zdunczyk, S.; Piskula, M.; Janowski, T.; Baranski, W.; Ras, M. Concentrations of isoflavones in blood plasma of dairy cows with different incidence of silent heat. Bull. Vet. Inst. Pulawy 2005, 49, 189–191. [Google Scholar]
- Kelly, R.W.; Adams, N.R.; Lindsay, D.R. Effect of coumestans on reproduction in the ewe. Aust. J. Agric. Res. 1976, 27, 253–259. [Google Scholar] [CrossRef]
- Sirtori, C.R.; Arnoldi, A.; Johnson, S.K. Phyto-oestrogens: End of a tale? Ann. Med. 2005, 37, 423–438. [Google Scholar] [CrossRef]
- Fields, R.L.; Moot, D.J.; Sedcole, J.R.; Barrell, G.K. Recovery of ovulation rate in ewes following their removal from an oestrogenic lucerne forage. Anim. Prod. Sci. 2019, 59, 493–498. [Google Scholar] [CrossRef]
- Burton, J.; Wells, M. The effect of phytoestrogens on the female genital tract. J. Clin. Pathol. 2002, 55, 401–407. [Google Scholar] [CrossRef]
- Smith, J.F.; Jagusch, K.T.; Brunswick, L.F.C.; Kelly, R.W. Coumestans in lucerne and ovulation in ewes. N. Z. J. Agric. Res. 1979, 22, 411–416. [Google Scholar] [CrossRef] [Green Version]
- Cederroth, C.R.; Auger, J.; Zimmermann, C.; Eustache, F.; Nef, S. Soy, phyto-oestrogens and male reproductive function: A review. Int. J. Androl. 2010, 33, 304–316. [Google Scholar] [CrossRef]
- Borzym, E.; Bobowiec, R.; Kosior-Korzecka, U.; Martelli, F.; Burmaczuk, A. Disturbances of cow oocyte maturation by phytoestrogens. Med. Weter. 2008, 64, 1107–1111. [Google Scholar]
- Nwannenna, A.I.; Lundh, T.; Madej, A.; Fredriksson, G.; Björnhag, G. Clinical changes in ovariectomized ewes exposed to phytoestrogens and 17β-estradiol implants. Exp. Biol. Med. 1995, 208, 92–97. [Google Scholar] [CrossRef]
- Adams, N.R. Detection of the effects of phyto-oestrogens on sheep and cattle. J. Anim. Sci. 1995, 73, 1509–1515. [Google Scholar] [CrossRef]
- Williamson, L.N.; Bartlett, M.G. Quantitative liquid chromatography/time-of-flight mass spectrometry. Biomed. Chromatogr. 2007, 21, 567–576. [Google Scholar] [CrossRef]
- Smith, C.A.; O’Maille, G.; Want, E.J.; Qin, C.; Trauger, S.A.; Brandon, T.R.; Custodio, D.E.; Abagyan, R.; Siuzdak, G. METLIN: A metabolite mass spectral database. Ther. Drug Monit. 2005, 27, 747–751. [Google Scholar] [CrossRef] [PubMed]
- Retana- Márquez, S.; Hernández, H.; Flores, J.A.; Muñoz-Gutiérrez, M.; Duarte, G.; Vielma, J.; Fitz-Rodríguez, G.; Fernández, I.G.; Keller, M.; Delgadillo, J.A. Effects of phytoestrogens on mammalian reproductive physiology. Trop. Subtrop. 2012, 15, 129–145. [Google Scholar]
- Kelly, R.W.; Lindsay, D.R. Plasma coumestrol levels and cervical mucus responses in ewes ingesting coumestan-rich feeds. Aust. J. Agric. Res. 1978, 29, 115–121. [Google Scholar] [CrossRef]
- Shutt, D.A.; Braden, A.W.H.; Lindner, H.R. Plasma coumestrol levels in sheep following administration of synthetic coumestrol or ingestion of medic hay (Medicago littoralis). Aust. J. Agric. Res. 1969, 20, 65–69. [Google Scholar] [CrossRef]
- Davies, H.L. Studies in uterotrophic activities in strains of subterranean clover. Aust. J. Agric. Res. 1989, 40, 871–877. [Google Scholar] [CrossRef]
- Butkutė, B.; Padarauskas, A.; Cesevičienė, J.; Taujenis, L.; Norkevičienė, E. Phytochemical composition of temperate perennial legumes. Crop Pasture Sci. 2018, 69, 1020–1030. [Google Scholar] [CrossRef]
- Cox, R.; Braden, A.W. The metabolism and physiological effects of phyto-oestrogens in livestock. Aust. Sot. Anim. Prod. 1974, 10, 122–129. [Google Scholar]
- Braden, A.W.H.; Hart, N.; Lamberton, J.A. Oestrogenic activity and metabolism of certain isoflavones in sheep. Aust. J. Agric. Res. 1967, 18, 348–355. [Google Scholar] [CrossRef]
- Virgona, J.; Ferguson, N.; Berntsen, E. Managing pasture composition: Doing the simple things well. In Proceedings of the Grassland Society of Southern Australia Inc. 60th Annual Conference Proceedings, Creswick, VIC, Australia, 17–18 July 2019; Grassland Society of Southern Australia: Carrum Downs, VIC, Australia, 2019; pp. 25–31. [Google Scholar]
- McIntyre, G. A method for unbiased selective sampling, using ranked sets. Am. Stat. 2005, 59, 230–232. [Google Scholar] [CrossRef]
- Ludwig, I.A.; Mena, P.; Calani, L.; Borges, G.; Pereira-Caro, G.; Bresciani, L.; Del Rio, D.; Lean, M.E.J.; Crozier, A. New insights into the bioavailability of red raspberry anthocyanins and ellagitannins. Free Radic. Biol. Med. 2015, 89, 758–769. [Google Scholar] [CrossRef] [Green Version]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cultivar | Fresh | ||||||
|---|---|---|---|---|---|---|---|
| Coumestrol | 3′Methoxycoumestrol | 4′Methoxycoumestrol | Daidzein | Formononetin | Genistein | TOTAL | |
| Genesis | 11.5 ± 3.7 | 8.7 ± 4.8 | 9.8 ± 7.9 | 2.9 ± 2.5 | 5.5 ± 2.6 | 57.0 ± 21.0 | 95.4 |
| SARDI 7 | 25.2 ± 3.7 | 16.8 ± 5.8 | 14.6 ± 2.2 | 3.6 ± 3.4 | 9.2 ± 3.4 | 137.0 ± 21.1 | 206.5 |
| Magna 804 | 21.8 ± 2.7 | 8.1 ± 4.4 | 11.6 ± 3.5 | 27.1 ± 7.7 | 15.6 ± 4.7 | 17.8 ± 4.2 | 102.1 |
| Cortez | 7.8 ± 0.6 | 6.2 ± 2.0 | 8.6 ± 2.6 | 3.6 ± 3.6 | 14.4 ± 6.4 | 26.0 ± 9.3 | 66.5 |
| Aurora | 4.1 ± 1.6 | 10.7 ± 2.8 | 5.4 ± 2.5 | 56.9 ± 11.0 | 9.8 ± 3.5 | 51.3 ± 11.5 | 138.2 |
| Silverado | 43.0 ± 17.4 | 46.1 ± 5.4 | 19.4 ± 7.8 | 12.8 ± 3.2 | 121.2 ± 36.0 | 91.3 ± 25.9 | 333.8 |
| Force 5 | 73.0 ± 32.7 | 4.1 ± 1.5 | 14.9 ± 3.2 | 32.8 ± 21.3 | 50.7 ± 36.0 | 79.5 ± 32.1 | 255 |
| LSD value | 17.5 | 5.1 | 12.5 | 15.6 | 29.4 | 38.4 | 55.3 |
| Dried | |||||||
| Genesis | 44.2 ± 13.4 | 7.0 ± 1.1 | 19.7 ± 10.0 | 9.2 ± 1.0 | 16.6 ± 6.1 | 2.4 ± 1.4 | 99.1 |
| SARDI 7 | 26.7 ± 8.2 | 10.0 ± 0.0 | 19.3 ± 5.8 | 17.4 ± 5.0 | 6.1 ± 1.6 | 6.5 ± 1.2 | 86 |
| Magna 804 | 27.8 ± 9.9 | 32.8 ± 1.7 | 18.7 ± 12.7 | 13.2 ± 1.2 | 17.8 ± 6.2 | 18.3 ± 10.7 | 128.8 |
| Cortez | 52.8 ± 26.1 | 14.3 ± 2.0 | 17.9 ± 13.5 | 10.1 ± 2.5 | 84.2 ± 26.1 | 2.6 ± 1.6 | 181.9 |
| Aurora | 65.9 ± 4.7 | 8.3 ± 2.8 | 18.5 ± 17.0 | 23.6 ± 14.0 | 0.2 ± 0.1 | 5.6 ± 2.4 | 122.1 |
| Silverado | 24.0 ± 24.0 | 56.1 ± 1.2 | 17.9 ± 1.3 | 0.5 ± 0.5 | 2.5 ± 1.1 | 8.3 ± 3.3 | 109.4 |
| Force 5 | 33.3 ± 4.1 | 3.4 ± 0.4 | 13.6 ± 4.1 | 7.1 ± 3.1 | 2.1 ± 1.4 | 3.7 ± 3.3 | 82 |
| LSD value | 23.8 | 34.8 | 10.8 | 9.9 | 21.5 | 11.5 | 51.8 |
| Day 0 | Day 4 | Day 12 | Day 20 | |||||
|---|---|---|---|---|---|---|---|---|
| C | T | C | T | C | T | C | T | |
| genistein | 0.312 ± 0.01 | 0.651 ± 0.05 | 0.613 ± 0.06 | 1.279 ± 0.14 | 0.996 ± 0.04 | 2.079 ± 1.10 | 1.971 ± 1.08 | 3.912 ± 1.98 |
| formononetin | 0.134 ± 0.01 | 0.379 ± 0.02 | 0.263 ± 0.04 | 0.745 ± 0.07 | 0.427 ± 0.08 | 1.210 ± 0.91 | 0.845 ± 0.08 | 2.316 ± 0.91 |
| daidzein | 2.796 ± 1.12 | 1.758 ± 1.02 | 5.497 ± 2.12 | 3.456 ± 1.24 | 3.383 ± 1.15 | 5.616 ± 2.58 | 6.693 ± 3.35 | 10.925 ± 3.98 |
| coumestrol | TD | 0.285 ± 0.01 | TD | 0.561 ± 0.04 | ND | 0.911 ± 0.18 | ND | 1.739 ± 1.08 |
| 4′methoxycoumestrol | ND | 0.015 ± 0.01 | ND | 0.030 ± 0.01 | ND | 0.049 ± 0.01 | ND | 0.092 ± 0.08 |
| 3′methoxycoumestrol | ND | 0.017 ± 0.01 | ND | 0.033 ± 0.01 | ND | 0.054 ± 0.02 | ND | 0.205 ± 0.04 |
| p-value | 0.003 | 0.003 | 0.009 | 0.007 | ||||
| * Cultivar | Dormancy | WAR | Seed Coat |
|---|---|---|---|
| 1 Force 5 | 5 | Semi winter dormant | Absent |
| 2 Aurora | 6 | Semi winter dormant | Absent |
| 3 SARDI 7 | 7 | Winter active | Present |
| 3 Genesis | 7 | Winter active | Present |
| 4 Magna 804 | 8 | Highly winter active | Absent |
| 5 Silverado | 9 | Highly winter active | Absent |
| 4 Cortez | 9 | Highly winter active | Absent |
| Treatment Number | Lucerne% | Other Species * % |
|---|---|---|
| Control | 0 | 100 |
| 1 | 50 | 50 |
| 2 | 70 | 30 |
| Name | Molecular Formula | Molecular Mass | [M-H] | Basis for Identification a | RT |
|---|---|---|---|---|---|
| Isoflavones | |||||
| daidzein | C15H10O4 | 254.24 | 253.0506 | STD | 8.11 |
| formononetin | C16H12O4 | 268.26 | 267.0663 | STD | 8.87 |
| genistein | C15H10O5 | 270.24 | 268.0455 | STD | 7.75 |
| Coumestans | |||||
| coumestrol | C15H8O5 | 268.22 | 267.0299 | STD | 7.88 |
| 3′methoxycoumestrol | C16H10O6 | 298.25 | 297.0405 | AM | 10.55 |
| 4′methoxycoumestrol | C16H10O5 | 282.25 | 281.0579 | AM | 8.77 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wyse, J.M.; Latif, S.; Gurusinghe, S.; Berntsen, E.D.; Weston, L.A.; Stephen, C.P. Characterization of Phytoestrogens in Medicago sativa L. and Grazing Beef Cattle. Metabolites 2021, 11, 550. https://doi.org/10.3390/metabo11080550
Wyse JM, Latif S, Gurusinghe S, Berntsen ED, Weston LA, Stephen CP. Characterization of Phytoestrogens in Medicago sativa L. and Grazing Beef Cattle. Metabolites. 2021; 11(8):550. https://doi.org/10.3390/metabo11080550
Chicago/Turabian StyleWyse, Jessica M., Sajid Latif, Saliya Gurusinghe, Erica D. Berntsen, Leslie A. Weston, and Cyril P. Stephen. 2021. "Characterization of Phytoestrogens in Medicago sativa L. and Grazing Beef Cattle" Metabolites 11, no. 8: 550. https://doi.org/10.3390/metabo11080550
APA StyleWyse, J. M., Latif, S., Gurusinghe, S., Berntsen, E. D., Weston, L. A., & Stephen, C. P. (2021). Characterization of Phytoestrogens in Medicago sativa L. and Grazing Beef Cattle. Metabolites, 11(8), 550. https://doi.org/10.3390/metabo11080550