
Early Postnatal Genistein Administration Affects Mice Metabolism and Reproduction in a Sexually Dimorphic Way

 , , ,
, , ,  ,
,  ,
, 
Abstract
:
1. Introduction
2. Results
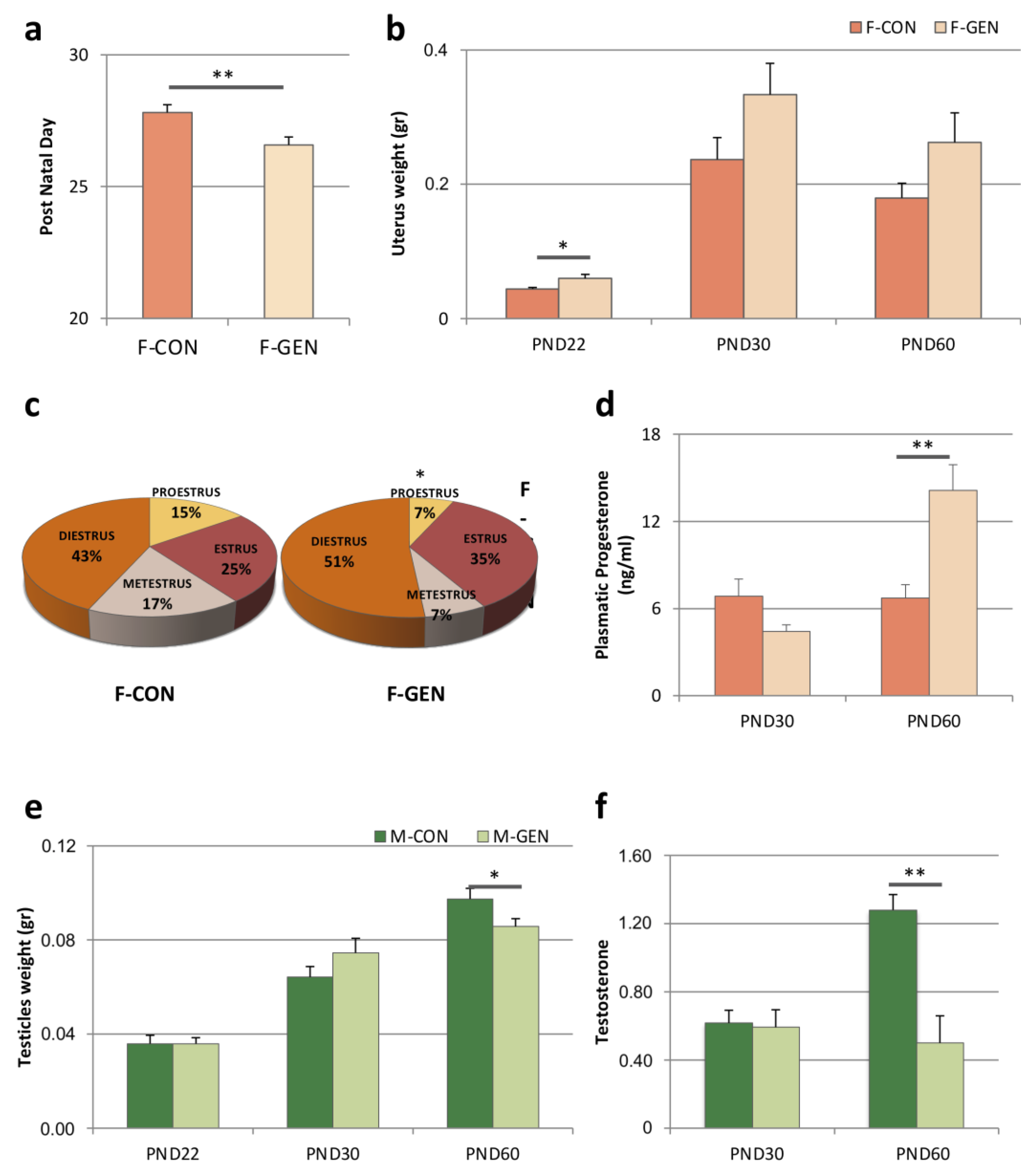
2.1. Female Reproductive Parameters
2.1.1. Vaginal Opening and Estrous Cycle
2.1.2. Uterus Weight
2.1.3. Progesterone Level
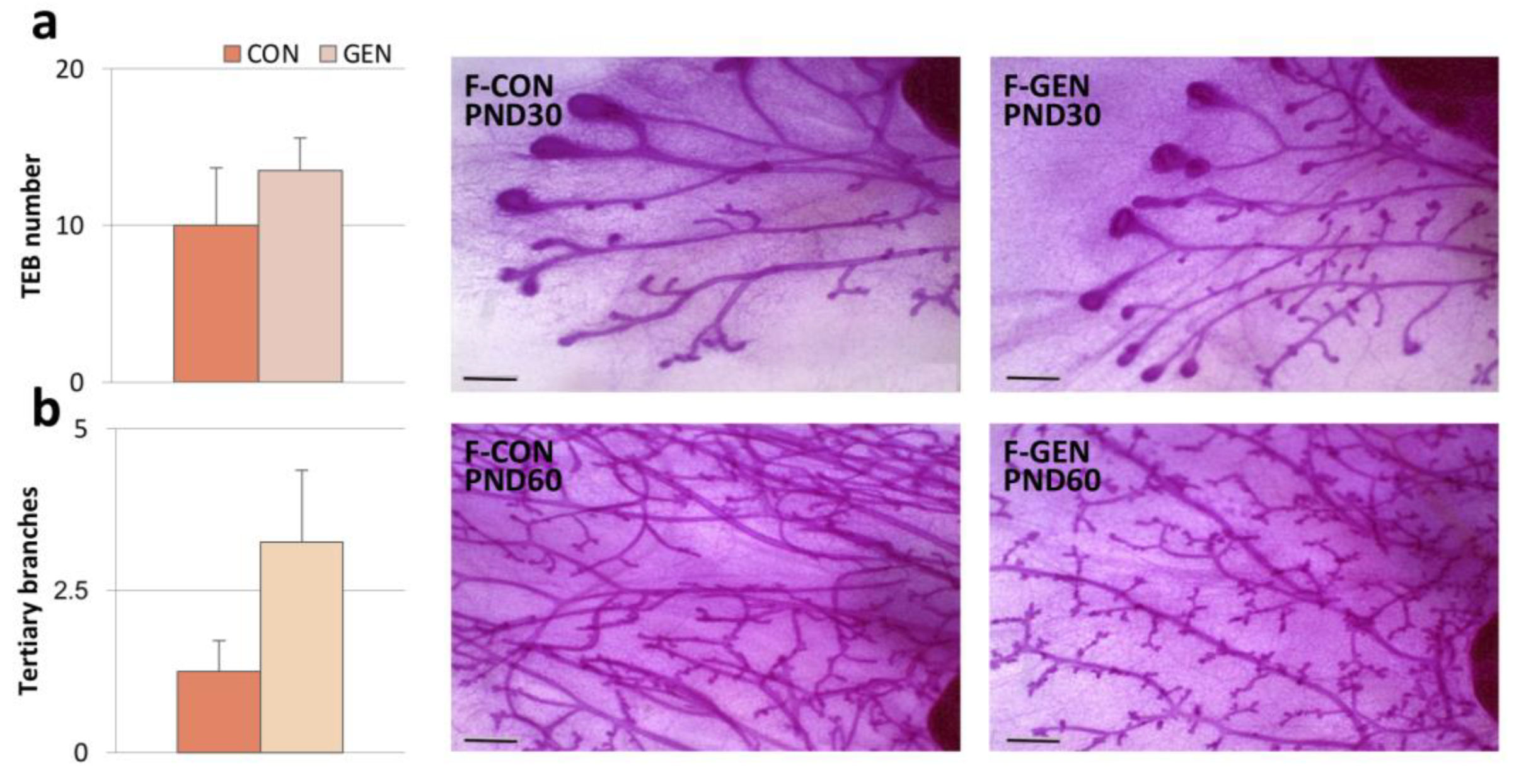
2.1.4. Mammary Gland Analysis
2.2. Male Reproductive Parameters
2.2.1. Testicle Weight
2.2.2. Testosterone Level
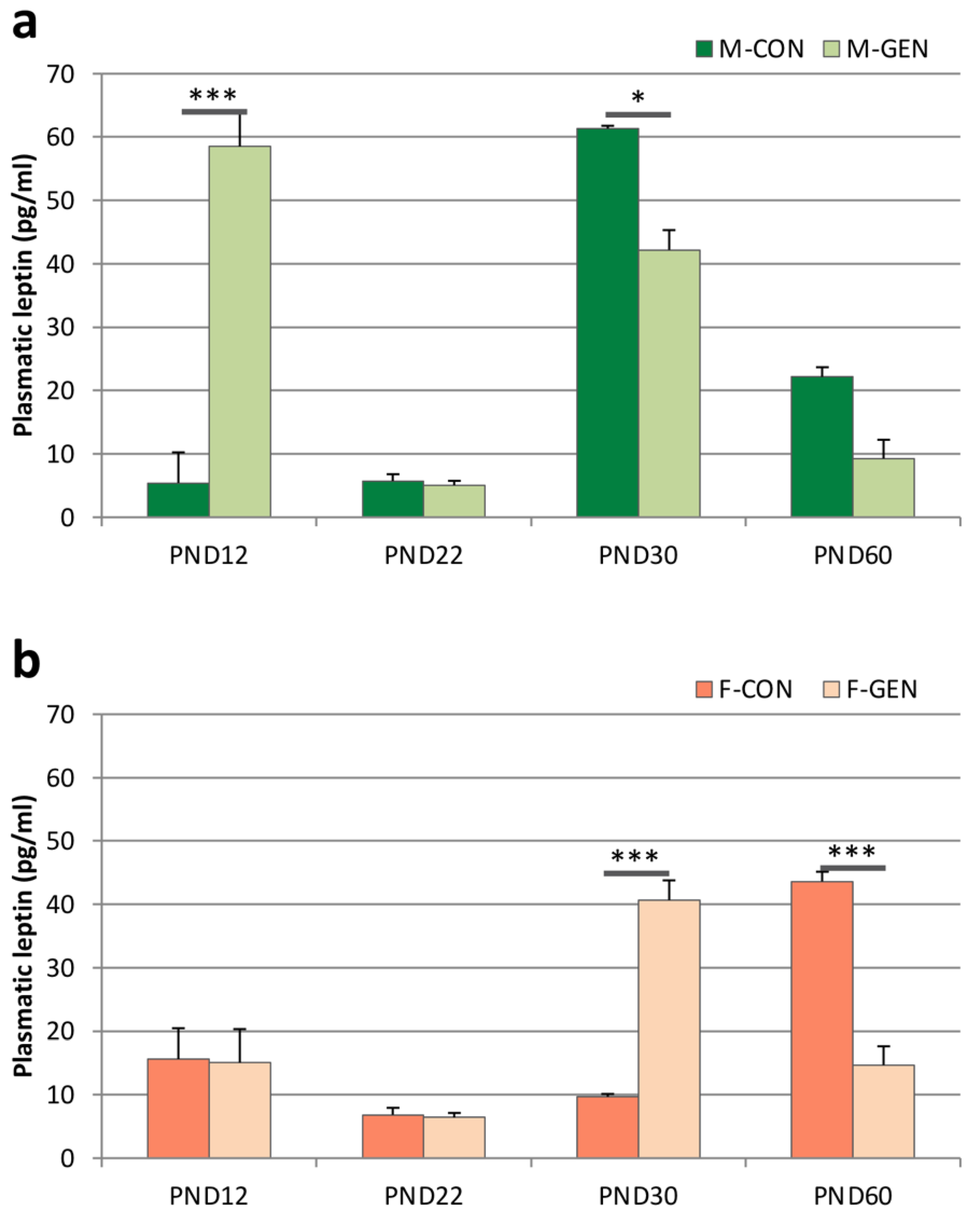
2.3. Leptin
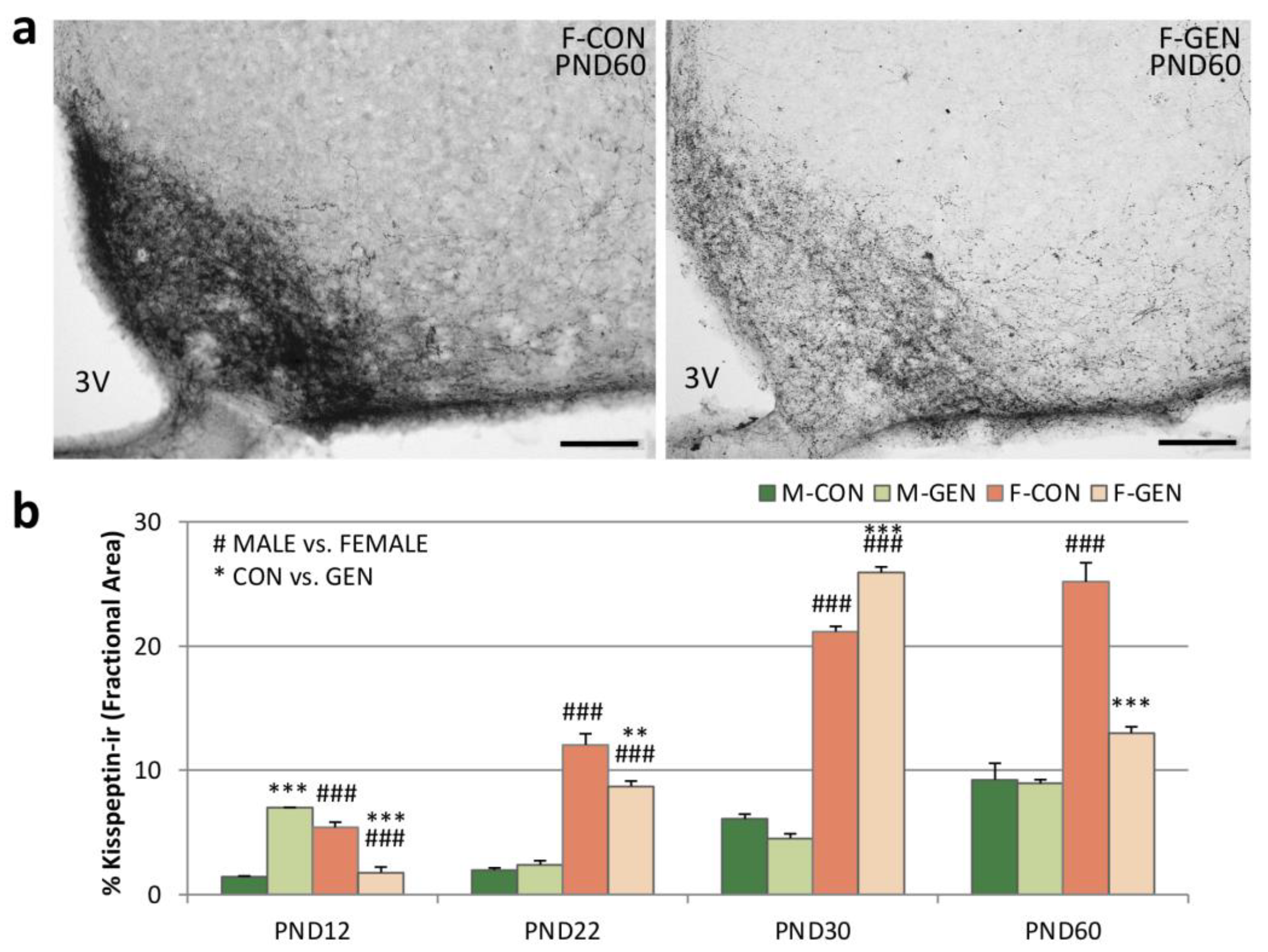
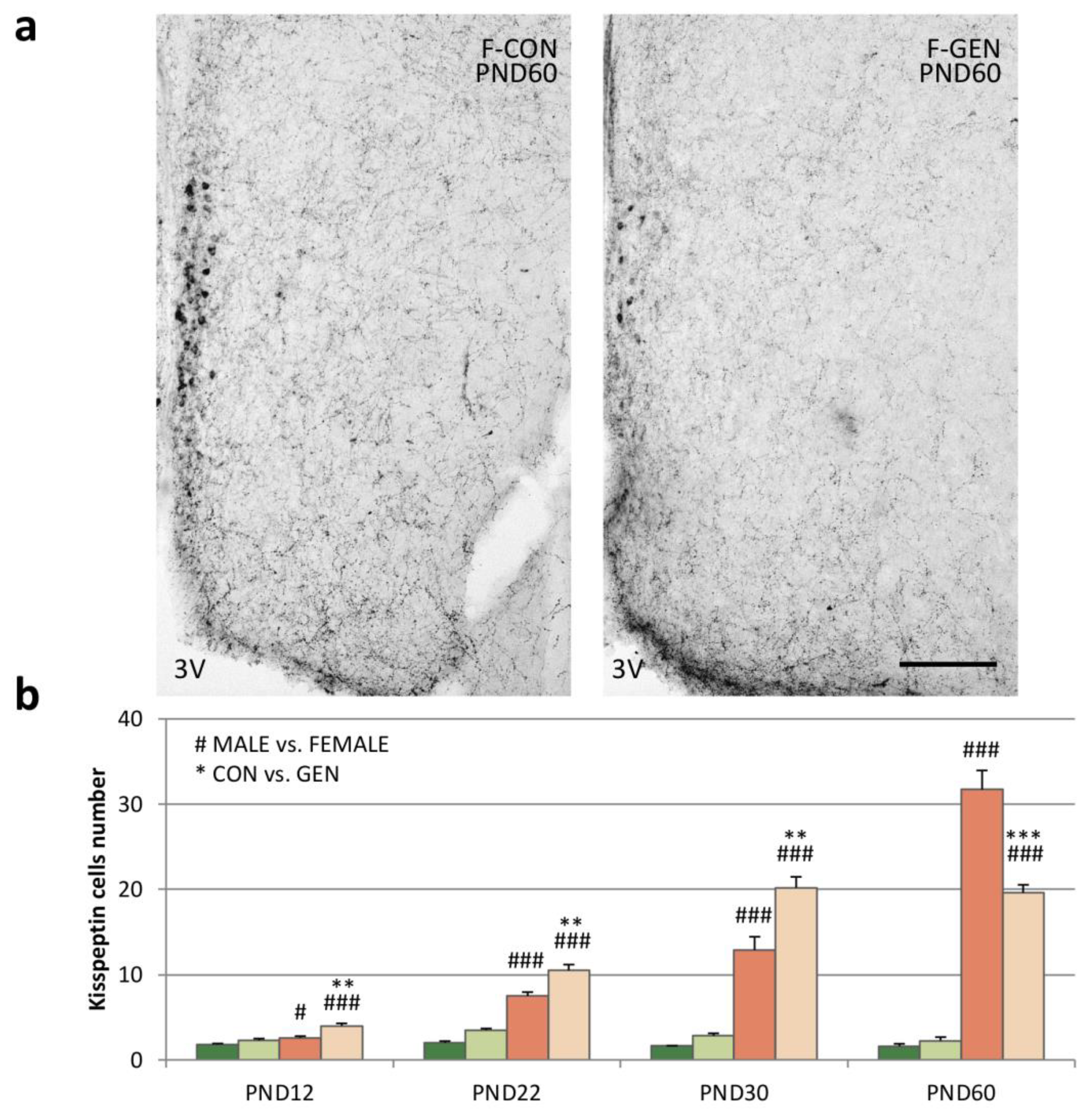
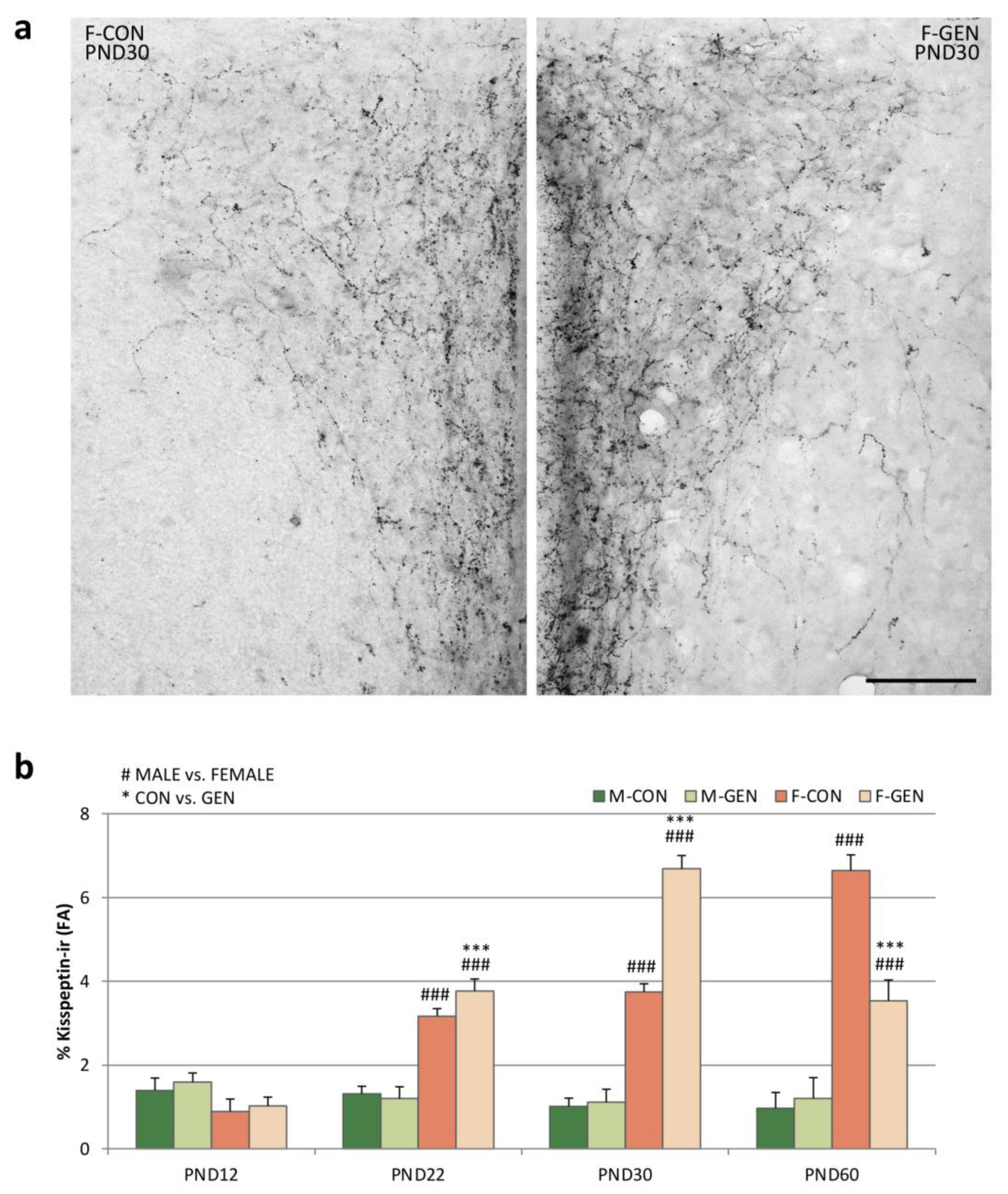
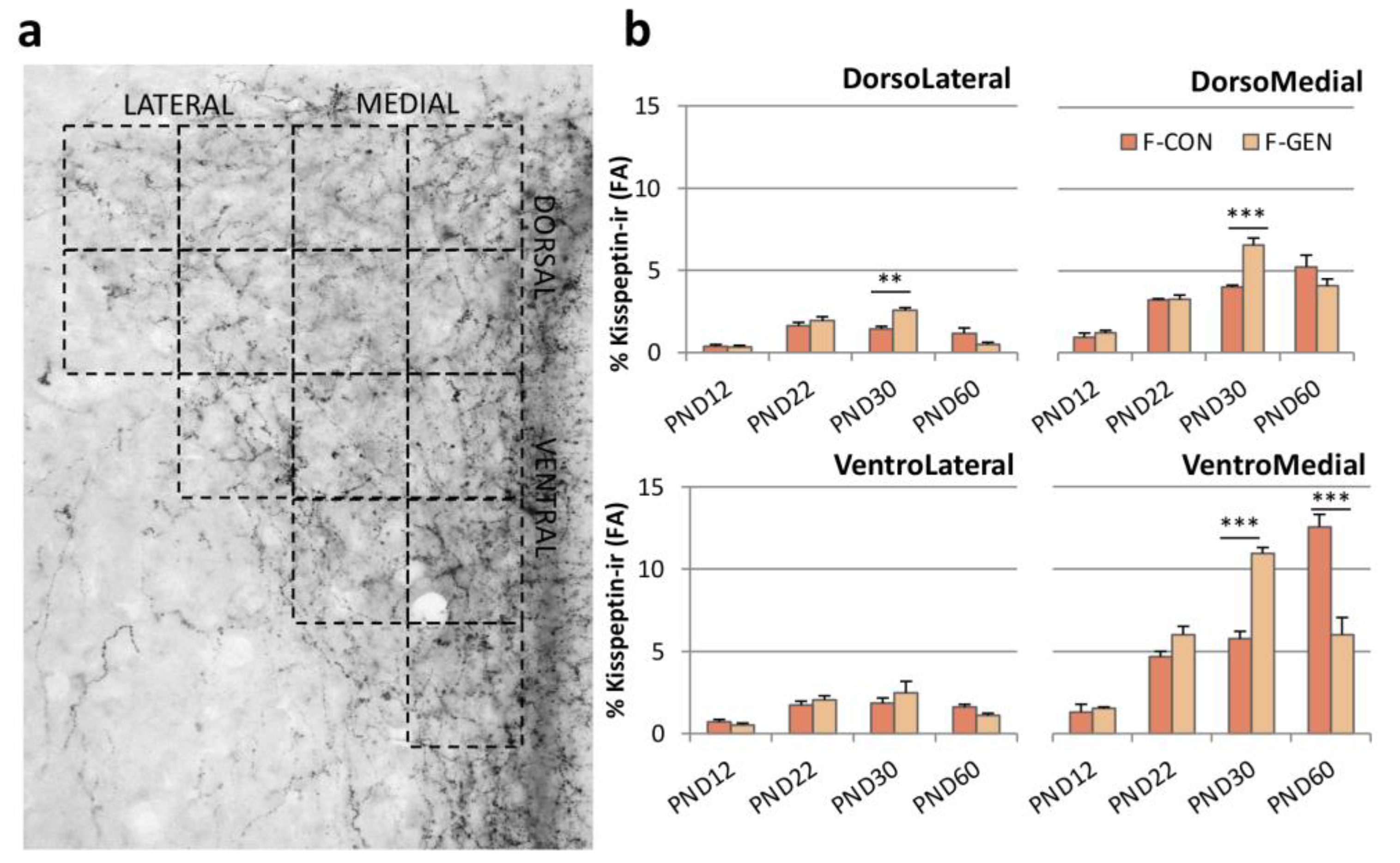
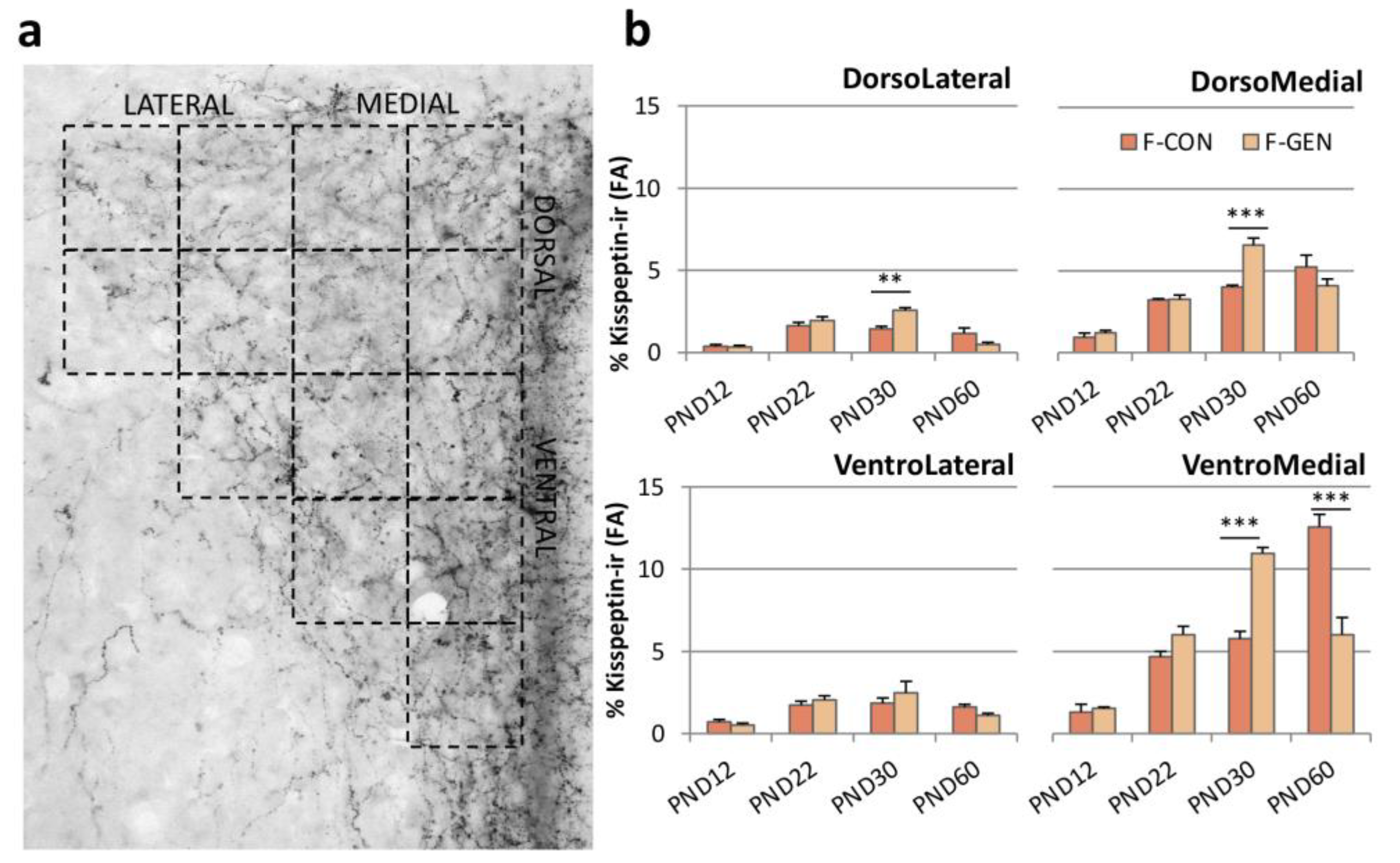
2.4. Kisspeptin System
2.5. Metabolic Parameters
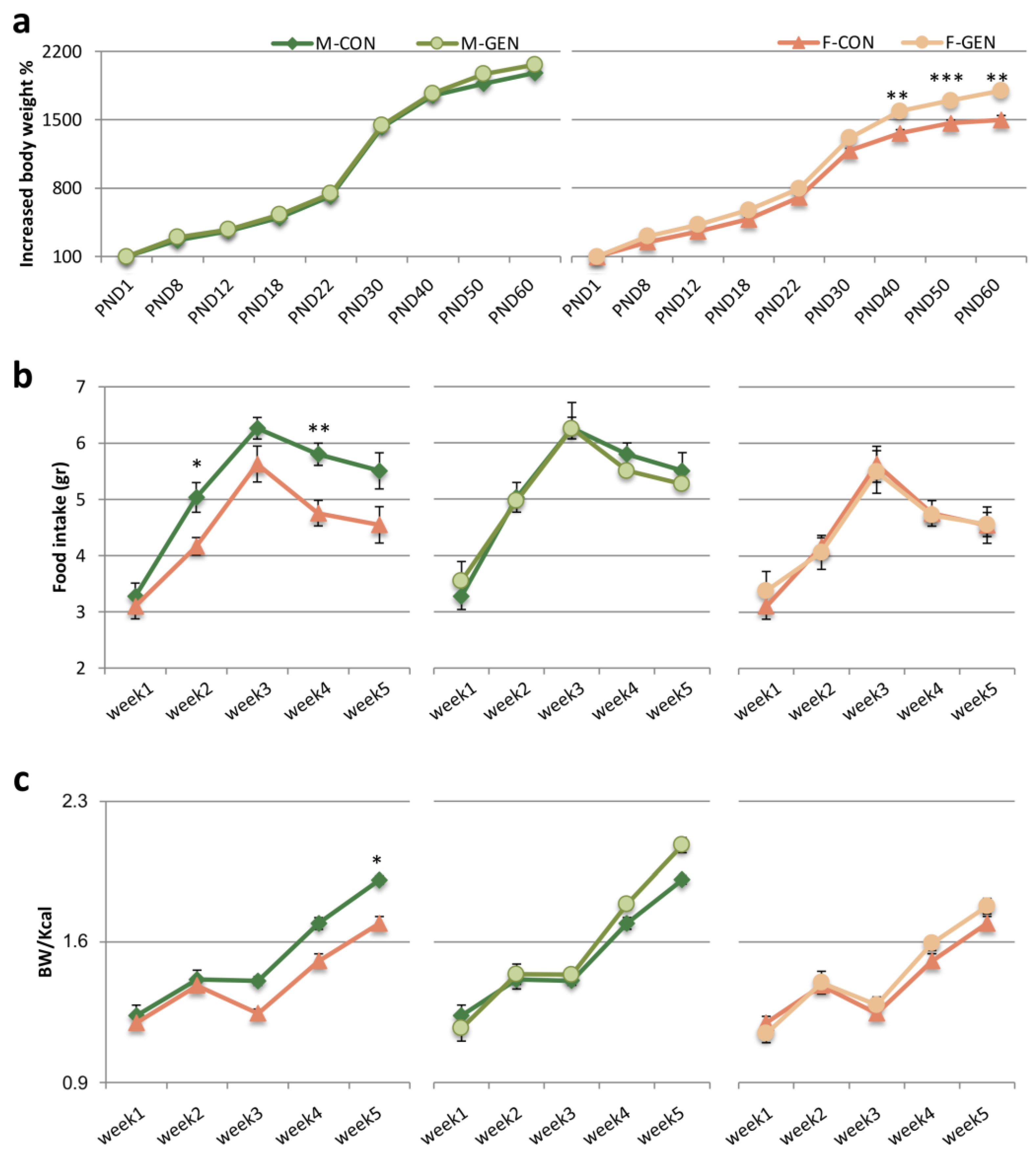
2.5.1. Body Weight, Food Consumption, and Feed Efficiency (FE)
2.5.2. Triiodothyronine (T3) and Ghrelin Levels
2.6. Hypothalamic Systems Controlling Food-Intake
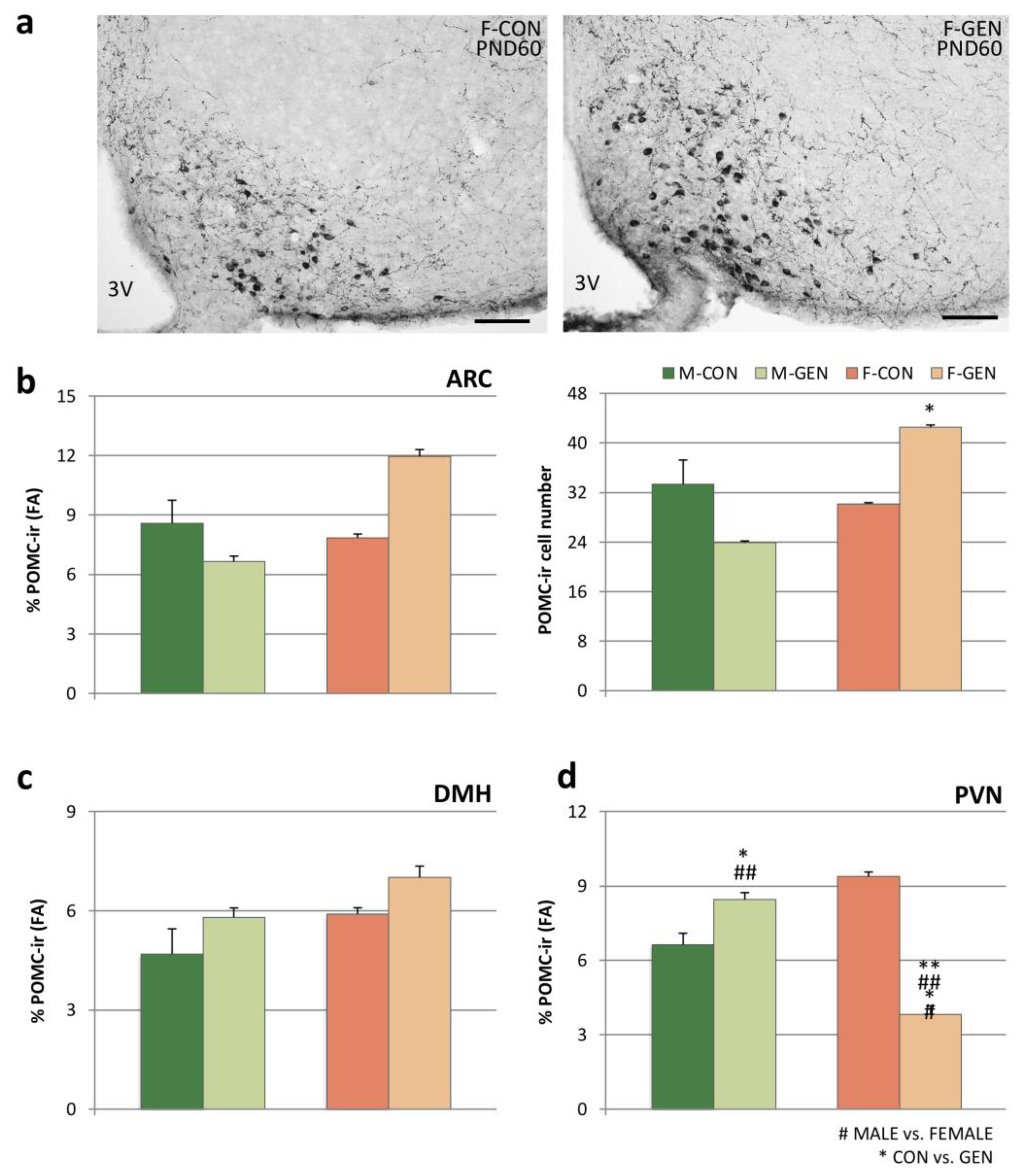
2.6.1. POMC System
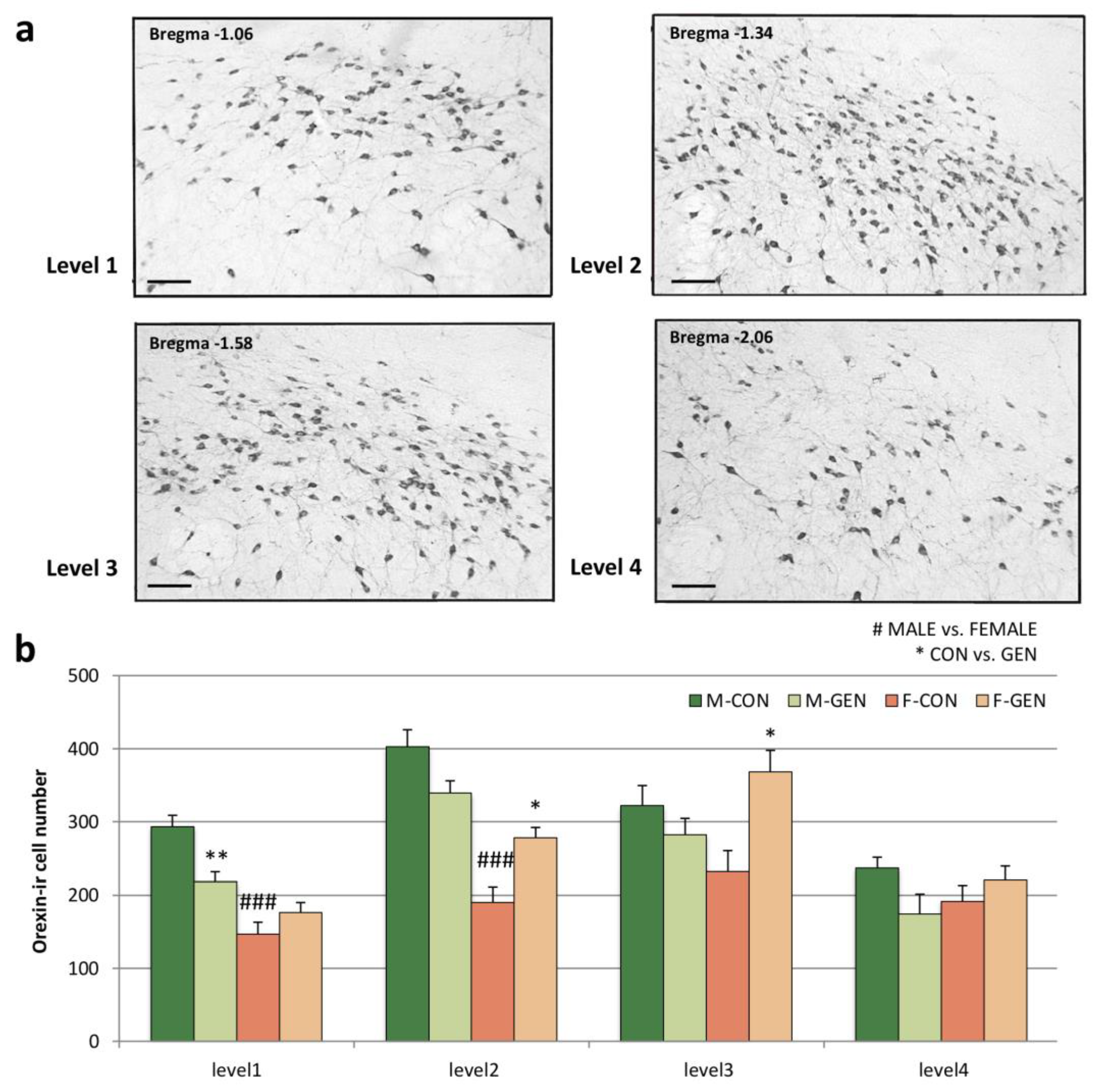
2.6.2. Orexin System
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Genistein Treatment
4.3. Reproductive and Metabolic Parameters
4.3.1. Vaginal Opening and Estrous Cycle
4.3.2. Uterus and Testicle Weight
4.3.3. Mammary Gland Analysis
4.3.4. Body Weight, Food Consumption, and Feed Efficiency (FE)
4.3.5. Hormonal Levels
4.4. Immunohistochemistry
4.4.1. Fixation and Sampling
4.4.2. Kisspeptin Immunohistochemistry
4.4.3. POMC Immunohistochemistry
4.4.4. Orexin Immunohistochemistry
4.5. Quantitative Analysis
4.5.1. Cell Counting and Fractional Area Evaluation
4.5.2. Kisspeptin
4.5.3. POMC
4.5.4. Orexin
4.6. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dixon, R. Genistein. Phytochemistry 2002, 60, 205–211. [Google Scholar] [CrossRef]
- Liggins, J.; Grimwood, R.; Bingham, S.A. Extraction and Quantification of Lignan Phytoestrogens in Food and Human Samples. Anal. Biochem. 2000, 287, 102–109. [Google Scholar] [CrossRef] [PubMed]
- Bhathena, S.J.; Velasquez, M.T. Beneficial role of dietary phytoestrogens in obesity and diabetes. Am. J. Clin. Nutr. 2002, 76, 1191–1201. [Google Scholar] [CrossRef] [Green Version]
- Aidelsburger, P.; Schauer, S.; Grabein, K.; Wasem, J. Alternative methods for the treatment of post-menopausal troubles. GMS Health Technol. Assess. 2012, 8, Doc 03. [Google Scholar] [CrossRef]
- Khan, S.; Zhao, J.; Khan, I.; A Walker, L.; DasMahapatra, A.K. Potential utility of natural products as regulators of breast cancer-associated aromatase promoters. Reprod. Biol. Endocrinol. 2011, 9, 91. [Google Scholar] [CrossRef] [Green Version]
- Hemati, N.; Asis, M.; Moradi, S.; Mollica, A.; Stefanucci, A.; Nikfar, S.; Mohammadi, E.; Farzaei, M.H.; Abdollahi, M. Effects of genistein on blood pressure: A systematic review and meta-analysis. Food Res. Int. 2020, 128, 108764. [Google Scholar] [CrossRef]
- Patisaul, H.B.; Jefferson, W. The pros and cons of phytoestrogens. Front. Neuroendocr. 2010, 31, 400–419. [Google Scholar] [CrossRef] [Green Version]
- Petrine, J.C.P.; Del Bianco-Borges, B. The influence of phytoestrogens on different physiological and pathological processes: An overview. Phytother. Res. 2021, 35, 180–197. [Google Scholar] [CrossRef] [PubMed]
- Setchell, K.D. Phytoestrogens: The biochemistry, physiology, and implications for human health of soy isoflavones. Am. J. Clin. Nutr. 1998, 68, 1333S–1346S. [Google Scholar] [CrossRef] [PubMed]
- Franke, A.; Custer, L.J.; Tanaka, Y. Isoflavones in human breast milk and other biological fluids. Am. J. Clin. Nutr. 1998, 68, 1466S–1473S. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Losa, S.M.; Todd, K.L.; Sullivan, A.W.; Cao, J.; Mickens, J.A.; Patisaul, H.B. Neonatal exposure to genistein adversely impacts the ontogeny of hypothalamic kisspeptin signaling pathways and ovarian development in the peripubertal female rat. Reprod. Toxicol. 2011, 31, 280–289. [Google Scholar] [CrossRef] [Green Version]
- Marraudino, M.; Farinetti, A.; Arevalo, M.-A.; Gotti, S.; Panzica, G.; Garcia-Segura, L.-M. Sexually Dimorphic Effect of Genistein on Hypothalamic Neuronal Differentiation in Vitro. Int. J. Mol. Sci. 2019, 20, 2465. [Google Scholar] [CrossRef] [Green Version]
- Patisaul, H.B.; Fortino, A.E.; Polston, E.K. Neonatal genistein or bisphenol-A exposure alters sexual differentiation of the AVPV. Neurotoxicol. Teratol. 2006, 28, 111–118. [Google Scholar] [CrossRef]
- Ponti, G.; Farinetti, A.; Marraudino, M.; Panzica, G.; Gotti, S. Postnatal genistein administration selectively abolishes sexual dimorphism in specific hypothalamic dopaminergic system in mice. Brain Res. 2019, 1724, 146434. [Google Scholar] [CrossRef] [PubMed]
- Ponti, G.; Rodriguez-Gomez, A.; Farinetti, A.; Marraudino, M.; Filice, F.; Foglio, B.; Sciacca, G.; Panzica, G.; Gotti, S. Early postnatal genistein administration permanently affects nitrergic and vasopressinergic systems in a sex-specific way. Neuroscience 2017, 346, 203–215. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Gomez, A.; Filice, F.; Gotti, S.; Panzica, G. Perinatal exposure to genistein affects the normal development of anxiety and aggressive behaviors and nitric oxide system in CD1 male mice. Physiol. Behav. 2014, 133, 107–114. [Google Scholar] [CrossRef] [PubMed]
- Timper, K.; Brüning, J.C. Hypothalamic circuits regulating appetite and energy homeostasis: Pathways to obesity. Dis. Model. Mech. 2017, 10, 679–689. [Google Scholar] [CrossRef] [Green Version]
- Messina, G.; Dalia, C.; Tafuri, D.; Monda, V.; Palmieri, F.; Dato, A.; Russo, A.; De Blasio, S.; Messina, A.; De Luca, V.; et al. Orexin-A controls sympathetic activity and eating behavior. Front. Psychol. 2014, 5, 997. [Google Scholar] [CrossRef] [Green Version]
- Cowley, M.; Smart, J.L.; Rubinstein, M.; Cerdán, M.G.; Diano, S.; Horvath, T.L.; Cone, R.D.; Low, M.J. Leptin activates anorexigenic POMC neurons through a neural network in the arcuate nucleus. Nat. Cell Biol. 2001, 411, 480–484. [Google Scholar] [CrossRef]
- Morello, G.; Imperatore, R.; Palomba, L.; Finelli, C.; Labruna, G.; Pasanisi, F.; Sacchetti, L.; Buono, L.; Piscitelli, F.; Orlando, P.; et al. Orexin-A represses satiety-inducing POMC neurons and contributes to obesity via stimulation of endocannabinoid signaling. Proc. Natl. Acad. Sci. USA 2016, 113, 4759–4764. [Google Scholar] [CrossRef] [Green Version]
- Bäckberg, M.; Hervieu, G.; Wilson, S.; Meister, B. Orexin receptor-1 (OX-R1) immunoreactivity in chemically identified neurons of the hypothalamus: Focus on orexin targets involved in control of food and water intake. Eur. J. Neurosci. 2002, 15, 315–328. [Google Scholar] [CrossRef]
- Kishi, T.; Aschkenasi, C.J.; Choi, B.J.; Lopez, M.E.; Lee, C.E.; Liu, H.; Hollenberg, A.N.; Friedman, J.M.; Elmquist, J.K. Neuropeptide Y Y1 receptor mRNA in rodent brain: Distribution and colocalization with melanocortin-4 receptor. J. Comp. Neurol. 2004, 482, 217–243. [Google Scholar] [CrossRef]
- van Swieten, M.; Pandit, R.; Adan, R.; van der Plasse, G. The neuroanatomical function of leptin in the hypothalamus. J. Chem. Neuroanat. 2014, 61-62, 207–220. [Google Scholar] [CrossRef] [PubMed]
- Marraudino, M.; Miceli, D.; Farinetti, A.; Ponti, G.; Panzica, G.; Gotti, S. Kisspeptin innervation of the hypothalamic paraventricular nucleus: Sexual dimorphism and effect of estrous cycle in female mice. J. Anat. 2017, 230, 775–786. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clarkson, J.; Herbison, A.E. Postnatal Development of Kisspeptin Neurons in Mouse Hypothalamus; Sexual Dimorphism and Projections to Gonadotropin-Releasing Hormone Neurons. Endocrinology 2006, 147, 5817–5825. [Google Scholar] [CrossRef] [PubMed]
- Herbison, A.E. Control of puberty onset and fertility by gonadotropin-releasing hormone neurons. Nat. Rev. Endocrinol. 2016, 12, 452–466. [Google Scholar] [CrossRef] [PubMed]
- Castellano, J.M.; Tena-Sempere, M. Metabolic control of female puberty: Potential therapeutic targets. Expert Opin. Ther. Targets 2016, 20, 1181–1193. [Google Scholar] [CrossRef]
- Qiu, X.; Dao, H.; Wang, M.; Heston, A.; Garcia, K.M.; Sangal, A.; Dowling, A.R.; Faulkner, L.D.; Molitor, S.; Elias, C.F.; et al. Insulin and Leptin Signaling Interact in the Mouse Kiss1 Neuron during the Peripubertal Period. PLoS ONE 2015, 10, e0121974. [Google Scholar] [CrossRef] [Green Version]
- Smith, J.; Acohido, B.V.; Clifton, D.K.; Steiner, R.A. KiSS-1 Neurones Are Direct Targets for Leptin in the ob/ob Mouse. J. Neuroendocr. 2006, 18, 298–303. [Google Scholar] [CrossRef]
- Cravo, R.; Margatho, L.; Osborne-Lawrence, S.; Donato, J.; Atkin, S.; Bookout, A.; Rovinsky, S.; Frazao, R.; Lee, C.; Gautron, L.; et al. Characterization of Kiss1 neurons using transgenic mouse models. Neuroscience 2011, 173, 37–56. [Google Scholar] [CrossRef] [Green Version]
- Louis, G.W.; Greenwald-Yarnell, M.; Phillips, R.; Coolen, L.; Lehman, M.; Myers, M.G. Molecular Mapping of the Neural Pathways Linking Leptin to the Neuroendocrine Reproductive Axis. Endocrinology 2011, 152, 2302–2310. [Google Scholar] [CrossRef] [Green Version]
- Quennell, J.H.; Mulligan, A.C.; Tups, A.; Liu, X.; Phipps, S.J.; Kemp, C.J.; Herbison, A.; Grattan, D.; Anderson, G.M. Leptin Indirectly Regulates Gonadotropin-Releasing Hormone Neuronal Function. Endocrinology 2009, 150, 2805–2812. [Google Scholar] [CrossRef] [Green Version]
- Fu, L.-Y.; Pol, A.N.V.D. Kisspeptin Directly Excites Anorexigenic Proopiomelanocortin Neurons but Inhibits Orexigenic Neuropeptide Y Cells by an Indirect Synaptic Mechanism. J. Neurosci. 2010, 30, 10205–10219. [Google Scholar] [CrossRef] [Green Version]
- Higo, S.; Iijima, N.; Ozawa, H. Characterisation ofKiss1r(Gpr54)-Expressing Neurones in the Arcuate Nucleus of the Female Rat Hypothalamus. J. Neuroendocr. 2017, 29, 29. [Google Scholar] [CrossRef] [PubMed]
- Orlando, G.; Leone, S.; Ferrante, C.; Chiavaroli, A.; Mollica, A.; Stefanucci, A.; Macedonio, G.; Dimmito, M.P.; Leporini, L.; Menghini, L.; et al. Effects of Kisspeptin-10 on Hypothalamic Neuropeptides and Neurotransmitters Involved in Appetite Control. Molecules 2018, 23, 3071. [Google Scholar] [CrossRef] [Green Version]
- Carrillo, B.; Collado, P.; Díaz, F.; Chowen, J.A.; Pérez-Izquierdo, M. Ángeles; Pinos, H. Physiological and brain alterations produced by high-fat diet in male and female rats can be modulated by increased levels of estradiol during critical periods of development. Nutr. Neurosci. 2017, 22, 29–39. [Google Scholar] [CrossRef] [PubMed]
- Marraudino, M.; Bonaldo, B.; Farinetti, A.; Panzica, G.; Ponti, G.; Gotti, S. Metabolism Disrupting Chemicals and Alteration of Neuroendocrine Circuits Controlling Food Intake and Energy Metabolism. Front. Endocrinol. 2019, 9, 766. [Google Scholar] [CrossRef]
- Jungbauer, A.; Medjakovic, S. Phytoestrogens and the metabolic syndrome. J. Steroid Biochem. Mol. Biol. 2014, 139, 277–289. [Google Scholar] [CrossRef] [PubMed]
- Cao, J.; Echelberger, R.; Liu, M.; Sluzas, E.; McCaffrey, K.; Buckley, B.; Patisaul, H.B. Soy but not bisphenol A (BPA) or the phytoestrogen genistin alters developmental weight gain and food intake in pregnant rats and their offspring. Reprod. Toxicol. 2015, 58, 282–294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, H.-K.; Nelson-Dooley, C.; Della-Fera, M.A.; Yang, J.-Y.; Zhang, W.; Duan, J.; Hartzell, D.L.; Hamrick, M.W.; Baile, C.A. Genistein Decreases Food Intake, Body Weight, and Fat Pad Weight and Causes Adipose Tissue Apoptosis in Ovariectomized Female Mice. J. Nutr. 2006, 136, 409–414. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naaz, A.; Yellayi, S.; Zakroczymski, M.A.; Bunick, D.; Doerge, D.R.; Lubahn, D.B.; Helferich, W.G.; Cooke, P.S. The Soy Isoflavone Genistein Decreases Adipose Deposition in Mice. Endocrinology 2003, 144, 3315–3320. [Google Scholar] [CrossRef]
- Penza, M.; Montani, C.; Romani, A.; Vignolini, P.; Pampaloni, B.; Tanini, A.; Brandi, M.L.; Magdalena, P.A.; Nadal, A.; Ottobrini, L.; et al. Genistein Affects Adipose Tissue Deposition in a Dose-Dependent and Gender-Specific Manner. Endocrinology 2006, 147, 5740–5751. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harmon, A.W.; Harp, J.B. Differential effects of flavonoids on 3T3-L1 adipogenesis and lipolysis. Am. J. Physiol. Physiol. 2001, 280, C807–C813. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zanella, I.; Marrazzo, E.; Biasiotto, G.; Penza, M.; Romani, A.; Vignolini, P.; Caimi, L.; Di Lorenzo, D. Soy and the soy isoflavone genistein promote adipose tissue development in male mice on a low-fat diet. Eur. J. Nutr. 2015, 54, 1095–1107. [Google Scholar] [CrossRef] [PubMed]
- Strakovsky, R.S.; Lezmi, S.; Flaws, J.A.; Schantz, S.L.; Pan, Y.-X.; Helferich, W.G. Genistein Exposure During the Early Postnatal Period Favors the Development of Obesity in Female, But Not Male Rats. Toxicol. Sci. 2013, 138, 161–174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.B.; Yan, J.D.; Yang, S.Q.; Guo, J.P.; Zhang, X.; Sun, X.X.; Na, X.L.; Dai, S.C. Maternal Genistein Intake Can Reduce Body Weight in Male Offspring. Biomed. Environ. Sci. 2015, 28, 769–772. [Google Scholar]
- Mueller, J.K.; Heger, S. Endocrine disrupting chemicals affect the Gonadotropin releasing hormone neuronal network. Reprod. Toxicol. 2014, 44, 73–84. [Google Scholar] [CrossRef]
- Patisaul, H.B. Effects of Environmental Endocrine Disruptors and Phytoestrogens on the Kisspeptin System. Adv. Exp. Med. Biol. 2013, 784, 455–479. [Google Scholar] [CrossRef] [PubMed]
- Patisaul, H.B.; Todd, K.L.; Mickens, J.A.; Adewale, H.B. Impact of neonatal exposure to the ERα agonist PPT, bisphenol-A or phytoestrogens on hypothalamic kisspeptin fiber density in male and female rats. NeuroToxicology 2009, 30, 350–357. [Google Scholar] [CrossRef] [Green Version]
- Bateman, H.L.; Patisaul, H.B. Disrupted female reproductive physiology following neonatal exposure to phytoestrogens or estrogen specific ligands is associated with decreased GnRH activation and kisspeptin fiber density in the hypothalamus. NeuroToxicology 2008, 29, 988–997. [Google Scholar] [CrossRef] [Green Version]
- Cederroth, C.R.; Vinciguerra, M.; Kühne, F.; Madani, R.; Doerge, D.R.; Visser, T.J.; Foti, M.; Rohner-Jeanrenaud, F.; Vassalli, J.-D.; Nef, S. A Phytoestrogen-Rich Diet Increases Energy Expenditure and Decreases Adiposity in Mice. Environ. Health Perspect. 2007, 115, 1467–1473. [Google Scholar] [CrossRef]
- Fernandez-Garcia, J.; Carrillo, B.; Tezanos, P.; Collado, P.; Pinos, H. Genistein during Development Alters Differentially the Expression of POMC in Male and Female Rats. Metabolism 2021, 11, 293. [Google Scholar] [CrossRef]
- Laffan, S.B.; Posobiec, L.M.; Uhl, J.E.; Vidal, J.D. Species Comparison of Postnatal Development of the Female Reproductive System. Birth Defects Res. 2018, 110, 163–189. [Google Scholar] [CrossRef]
- Paine, I.S.; Lewis, M.T. The Terminal End Bud: The Little Engine that Could. J. Mammary Gland. Biol. Neoplasia 2017, 22, 93–108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Macias, H.; Hinck, L. Mammary gland development. Wiley Interdiscip. Rev. Dev. Biol. 2012, 1, 533–557. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marraudino, M.; Martini, M.; Trova, S.; Farinetti, A.; Ponti, G.; Gotti, S.; Panzica, G. Kisspeptin system in ovariectomized mice: Estradiol and progesterone regulation. Brain Res. 2018, 1688, 8–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farinetti, A.; Marraudino, M.; Ponti, G.; Panzica, G.; Gotti, S. Chronic treatment with tributyltin induces sexually dimorphic alterations in the hypothalamic POMC system of adult mice. Cell Tissue Res. 2018, 374, 587–594. [Google Scholar] [CrossRef]
- Ferguson, A.V.; Samson, W.K. The orexin/hypocretin system: A critical regulator of neuroendocrine and autonomic function. Front. Neuroendocr. 2003, 24, 141–150. [Google Scholar] [CrossRef]
- Paxinos, G.; Franklin, K.B.J.; Franklin, K.B.J. The Mouse Brain in Stereotaxic Coordinates, 2nd ed.; Academic Press: San Diego, CA, USA, 2001. [Google Scholar]
- Crain, S.M.; Shen, K.-F. Low doses of cyclic AMP-phosphodiesterase inhibitors rapidly evoke opioid receptor-mediated thermal hyperalgesia in naïve mice which is converted to prominent analgesia by cotreatment with ultra-low-dose naltrexone. Brain Res. 2008, 1231, 16–24. [Google Scholar] [CrossRef]
- Jefferson, W.N.; Padilla-Banks, E.; Newbold, R.R. Studies of the Effects of Neonatal Exposure to Genistein on the Developing Female Reproductive System. J. AOAC Int. 2006, 89, 1189–1196. [Google Scholar] [CrossRef]
- Jefferson, W.N.; Padilla-Banks, E.; Newbold, R.R. Disruption of the developing female reproductive system by phytoestrogens: Genistein as an example. Mol. Nutr. Food Res. 2007, 51, 832–844. [Google Scholar] [CrossRef]
- Rudel, R.A.; Fenton, S.E.; Ackerman, J.M.; Euling, S.Y.; Makris, S.L. Environmental Exposures and Mammary Gland Development: State of the Science, Public Health Implications, and Research Recommendations. Environ. Health Perspect. 2011, 119, 1053–1061. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muñoz-De-Toro, M.; Markey, C.M.; Wadia, P.R.; Luque, E.H.; Rubin, B.S.; Sonnenschein, C.; Soto, A.M. Perinatal Exposure to Bisphenol-A Alters Peripubertal Mammary Gland Development in Mice. Endocrinology 2005, 146, 4138–4147. [Google Scholar] [CrossRef]
- Li, R.; El Zowalaty, A.E.; Chen, W.; Dudley, E.A.; Ye, X. Segregated responses of mammary gland development and vaginal opening to prepubertal genistein exposure in Bscl2(-/-) female mice with lipodystrophy. Reprod. Toxicol. 2015, 54, 76–83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruiz-Pino, F.; Miceli, D.; Franssen, D.; Vazquez, M.J.; Farinetti, A.; Castellano, J.M.; Panzica, G.; Tena-Sempere, M. Environmentally Relevant Perinatal Exposures to Bisphenol A Disrupt Postnatal Kiss1/NKB Neuronal Maturation and Puberty Onset in Female Mice. Environ. Health Perspect. 2019, 127, 107011. [Google Scholar] [CrossRef]
- Luque, R.M.; Kineman, R.; Tena-Sempere, M. Regulation of Hypothalamic Expression of KiSS-1 and GPR54 Genes by Metabolic Factors: Analyses Using Mouse Models and a Cell Line. Endocrinology 2007, 148, 4601–4611. [Google Scholar] [CrossRef] [PubMed]
- Szkudelska, K.; Nogowski, L. Genistein—A dietary compound inducing hormonal and metabolic changes. J. Steroid Biochem. Mol. Biol. 2007, 105, 37–45. [Google Scholar] [CrossRef] [PubMed]
- Szkudelski, T.; Nogowski, L.; Pruszyńska-Oszmałek, E.; Kaczmarek, P.; Szkudelska, K. Genistein restricts leptin secretion from rat adipocytes. J. Steroid Biochem. Mol. Biol. 2005, 96, 301–307. [Google Scholar] [CrossRef]
- Nogowski, L.; Szkudelska, K.; Szkudelski, T.; Pruszyńska-Oszmałek, E. The effect of a phytoestrogen, genistein, on the hormonal and metabolic status of pregnant rats. J. Anim. Feed. Sci. 2006, 15, 275–286. [Google Scholar] [CrossRef]
- Li, R.-Z.; Ding, X.-W.; Geetha, T.; Al-Nakkash, L.; Broderick, T.L.; Babu, J.R. Beneficial Effect of Genistein on Diabetes-Induced Brain Damage in the ob/ob Mouse Model. Drug Des. Dev. Ther. 2020, ume 14, 3325–3336. [Google Scholar] [CrossRef]
- Weigt, C.; Hertrampf, T.; Zoth, N.; Fritzemeier, K.H.; Diel, P. Impact of estradiol, ER subtype specific agonists and genistein on energy homeostasis in a rat model of nutrition induced obesity. Mol. Cell. Endocrinol. 2012, 351, 227–238. [Google Scholar] [CrossRef]
- Heindel, J.J.; Saal, F.S.V.; Blumberg, B.; Bovolin, P.; Calamandrei, G.; Ceresini, G.; Cohn, B.A.; Fabbri, E.; Gioiosa, L.; Kassotis, C.; et al. Parma consensus statement on metabolic disruptors. Environ. Health 2015, 14, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Snedeker, S.M.; Hay, A. Do Interactions Between Gut Ecology and Environmental Chemicals Contribute to Obesity and Diabetes? Environ. Health Perspect. 2012, 120, 332–339. [Google Scholar] [CrossRef]
- Heindel, J.J.; Blumberg, B.; Cave, M.; Machtinger, R.; Mantovani, A.; Mendez, M.A.; Nadal, A.; Palanza, P.; Panzica, G.; Sargis, R.; et al. Metabolism disrupting chemicals and metabolic disorders. Reprod. Toxicol. 2017, 68, 3–33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ryökkynen, A.; Kukkonen, J.; Nieminen, P. Effects of dietary genistein on mouse reproduction, postnatal development and weight-regulation. Anim. Reprod. Sci. 2006, 93, 337–348. [Google Scholar] [CrossRef]
- Wright, S.M.; Salter, A.M. Effects of Soy Protein on Plasma Cholesterol and Bile Acid Excretion in Hamsters. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 1998, 119, 247–254. [Google Scholar] [CrossRef]
- Nieminen, P.; Mustonen, A.-M.; Lindström-Seppä, P.; Asikainen, J.; Mussalo-Rauhamaa, H.; Kukkonen, J. Phytosterols Act as Endocrine and Metabolic Disruptors in the European Polecat (Mustela putorius). Toxicol. Appl. Pharmacol. 2002, 178, 22–28. [Google Scholar] [CrossRef]
- Ryökkynen, A.; Mustonen, A.-M.; Pyykönen, T.; Nieminen, P. Endocrine and metabolic alterations in the mink (Mustela vison) due to chronic phytoestrogen exposure. Chemosphere 2006, 64, 1753–1760. [Google Scholar] [CrossRef] [PubMed]
- Cheung, C.C.; Clifton, D.K.; Steiner, R.A. Proopiomelanocortin Neurons Are Direct Targets for Leptin in the Hypothalamus. Endocrinology 1997, 138, 4489–4492. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Faulkner, L.D.; Hill, J.W. Cross-talk between metabolism and reproduction: The role of POMC and SF1 neurons. Front. Endocrinol. 2012, 2, 98. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marraudino, M.; Bo, E.; Carlini, E.; Farinetti, A.; Ponti, G.; Zanella, I.; Di Lorenzo, D.; Panzica, G.C.; Gotti, S. Hypothalamic Expression of Neuropeptide Y (NPY)and Pro-OpioMelanoCortin (POMC) in Adult Male Mice is Affected by Chronic Exposure to Endocrine Disruptors. Metabolites 2021, 11, 368. [Google Scholar] [CrossRef]
- Chieffi, S.; Carotenuto, M.; Monda, V.; Valenzano, A.; Villano, I.; Precenzano, F.; Tafuri, D.; Salerno, M.; Filippi, N.; Nuccio, F.; et al. Orexin System: The Key for a Healthy Life. Front. Physiol. 2017, 8, 357. [Google Scholar] [CrossRef] [Green Version]
- Brownell, S.E.; Conti, B. Age- and gender-specific changes of hypocretin immunopositive neurons in C57Bl/6 mice. Neurosci. Lett. 2010, 472, 29–32. [Google Scholar] [CrossRef] [Green Version]
- Muschamp, J.W.; Dominguez, J.M.; Sato, S.M.; Shen, R.-Y.; Hull, E. A Role for Hypocretin (Orexin) in Male Sexual Behavior. J. Neurosci. 2007, 27, 2837–2845. [Google Scholar] [CrossRef] [Green Version]
- Kalra, S.P.; Kalra, P.S. Neuroendocrine Control of Energy Homeostasis: Update on New Insights. Prog. Brain Res. 2010, 181, 17–33. [Google Scholar] [CrossRef] [PubMed]
- Biglari, N.; Gaziano, I.; Schumacher, J.; Radermacher, J.; Paeger, L.; Klemm, P.; Chen, W.; Corneliussen, S.; Wunderlich, C.M.; Sue, M.; et al. Functionally distinct POMC-expressing neuron subpopulations in hypothalamus revealed by intersectional targeting. Nat. Neurosci. 2021, 24, 913–929. [Google Scholar] [CrossRef] [PubMed]
- Gaskins, G.T.; Moenter, S.M. Orexin A Suppresses Gonadotropin-Releasing Hormone (GnRH) Neuron Activity in the Mouse. Endocrinology 2012, 153, 3850–3860. [Google Scholar] [CrossRef] [Green Version]
- Hosseini, A.; Khazali, H. Central Orexin A Affects Reproductive Axis by Modulation of Hypothalamic Kisspeptin/Neurokinin B/Dynorphin Secreting Neurons in the Male Wistar Rats. Neuromolecular Med. 2018, 20, 525–536. [Google Scholar] [CrossRef]
- Barreiro, M.L.; Pineda, R.; Navarro, V.M.; López, M.; Suominen, J.S.; Pinilla, L.; Señaris, R.; Toppari, J.; Aguilar, E.; Diéguez, C.; et al. Orexin 1 Receptor Messenger Ribonucleic Acid Expression and Stimulation of Testosterone Secretion by Orexin-A in Rat Testis. Endocrinology 2004, 145, 2297–2306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manno, F.A.M. Measurement of the digit lengths and the anogenital distance in mice. Physiol. Behav. 2008, 93, 364–368. [Google Scholar] [CrossRef] [PubMed]
- Cimafranca, M.A.; Davila, J.; Ekman, G.C.; Andrews, R.N.; Neese, S.L.; Peretz, J.; Woodling, K.A.; Helferich, W.G.; Sarkar, J.; Flaws, J.; et al. Acute and Chronic Effects of Oral Genistein Administration in Neonatal Mice1. Biol. Reprod. 2010, 83, 114–121. [Google Scholar] [CrossRef] [Green Version]
- Doerge, D.R.; Twaddle, N.C.; Banks, E.P.; Jefferson, W.N.; Newbold, R.R. Pharmacokinetic analysis in serum of genistein administered subcutaneously to neonatal mice. Cancer Lett. 2002, 184, 21–27. [Google Scholar] [CrossRef]
- Bo, E.; Farinetti, A.; Marraudino, M.; Sterchele, D.; Eva, C.; Gotti, S.; Panzica, G. Adult exposure to tributyltin affects hypothalamic neuropeptide Y, Y1 receptor distribution, and circulating leptin in mice. Andrology 2016, 4, 723–734. [Google Scholar] [CrossRef]
- Macchi, E.; Cucuzza, A.S.; Badino, P.; Odore, R.; Re, F.; Bevilacqua, L.; Malfatti, A. Seasonality of reproduction in wild boar (Sus scrofa) assessed by fecal and plasmatic steroids. Theriogenology 2010, 73, 1230–1237. [Google Scholar] [CrossRef] [PubMed]
- Watson, R.E.; Wiegand, S.J.; Clough, R.W.; Hoffman, G.E. Use of cryoprotectant to maintain long-term peptide immunoreactivity and tissue morphology. Peptides 1986, 7, 155–159. [Google Scholar] [CrossRef]
- Franceschini, I.; Lomet, D.; Cateau, M.; Delsol, G.; Tillet, Y.; Caraty, A. Kisspeptin immunoreactive cells of the ovine preoptic area and arcuate nucleus co-express estrogen receptor alpha. Neurosci. Lett. 2006, 401, 225–230. [Google Scholar] [CrossRef] [PubMed]
- Pinos, H.; Pérez-Izquierdo, M.; Carrillo, B.; Collado, P. Effects of undernourishment on the hypothalamic orexinergic system. Physiol. Behav. 2011, 102, 17–21. [Google Scholar] [CrossRef]
- Mize, R.; Holdefer, R.N.; Nabors, L. Quantitative immunocytochemistry using an image analyzer. I. Hardware evaluation, image processing, and data analysis. J. Neurosci. Methods 1988, 26, 1–23. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plasma Concentration of T3 (ng/mL) | ||||
| M-CON | M-GEN | F-CON | F-GEN | |
| (mean ± SEM) | (mean ± SEM) | (mean ± SEM) | (mean ± SEM) | |
| PND30 | 5.3 ± 0.32 | 5.8 ± 0.28 | 5.6 ± 0.19 | 6.2 ± 0.34 |
| PND60 | 6.4 ± 0.06 | 5.7 ± 0.11 | 7.5 ± 0.19 | 6.3 ± 0.09 |
| Plasma Concentration of ghrelin (ng/mL) | ||||
| M-CON | M-GEN | F-CON | F-GEN | |
| (mean ± SEM) | (mean ± SEM) | (mean ± SEM) | (mean ± SEM) | |
| PND30 | 0.30 ± 0.04 | 0.40 ± 0.06 | 0.34 ± 0.07 | 0.7 ± 0.11 |
| PND60 | 0.87 ± 0.18 | 4.23 ± 0.69 | 4.33 ± 0.73 | 2.76 ± 0.26 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Marraudino, M.; Ponti, G.; Moussu, C.; Farinetti, A.; Macchi, E.; Accornero, P.; Gotti, S.; Collado, P.; Keller, M.; Panzica, G. Early Postnatal Genistein Administration Affects Mice Metabolism and Reproduction in a Sexually Dimorphic Way. Metabolites 2021, 11, 449. https://doi.org/10.3390/metabo11070449
Marraudino M, Ponti G, Moussu C, Farinetti A, Macchi E, Accornero P, Gotti S, Collado P, Keller M, Panzica G. Early Postnatal Genistein Administration Affects Mice Metabolism and Reproduction in a Sexually Dimorphic Way. Metabolites. 2021; 11(7):449. https://doi.org/10.3390/metabo11070449
Chicago/Turabian StyleMarraudino, Marilena, Giovanna Ponti, Chantal Moussu, Alice Farinetti, Elisabetta Macchi, Paolo Accornero, Stefano Gotti, Paloma Collado, Matthieu Keller, and Giancarlo Panzica. 2021. "Early Postnatal Genistein Administration Affects Mice Metabolism and Reproduction in a Sexually Dimorphic Way" Metabolites 11, no. 7: 449. https://doi.org/10.3390/metabo11070449
APA StyleMarraudino, M., Ponti, G., Moussu, C., Farinetti, A., Macchi, E., Accornero, P., Gotti, S., Collado, P., Keller, M., & Panzica, G. (2021). Early Postnatal Genistein Administration Affects Mice Metabolism and Reproduction in a Sexually Dimorphic Way. Metabolites, 11(7), 449. https://doi.org/10.3390/metabo11070449