
Measurement of Pulsatile Insulin Secretion: Rationale and Methodology

{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
1.1. Insulin Physiology
1.2. Mechanisms of Pulsatile Insulin Secretion
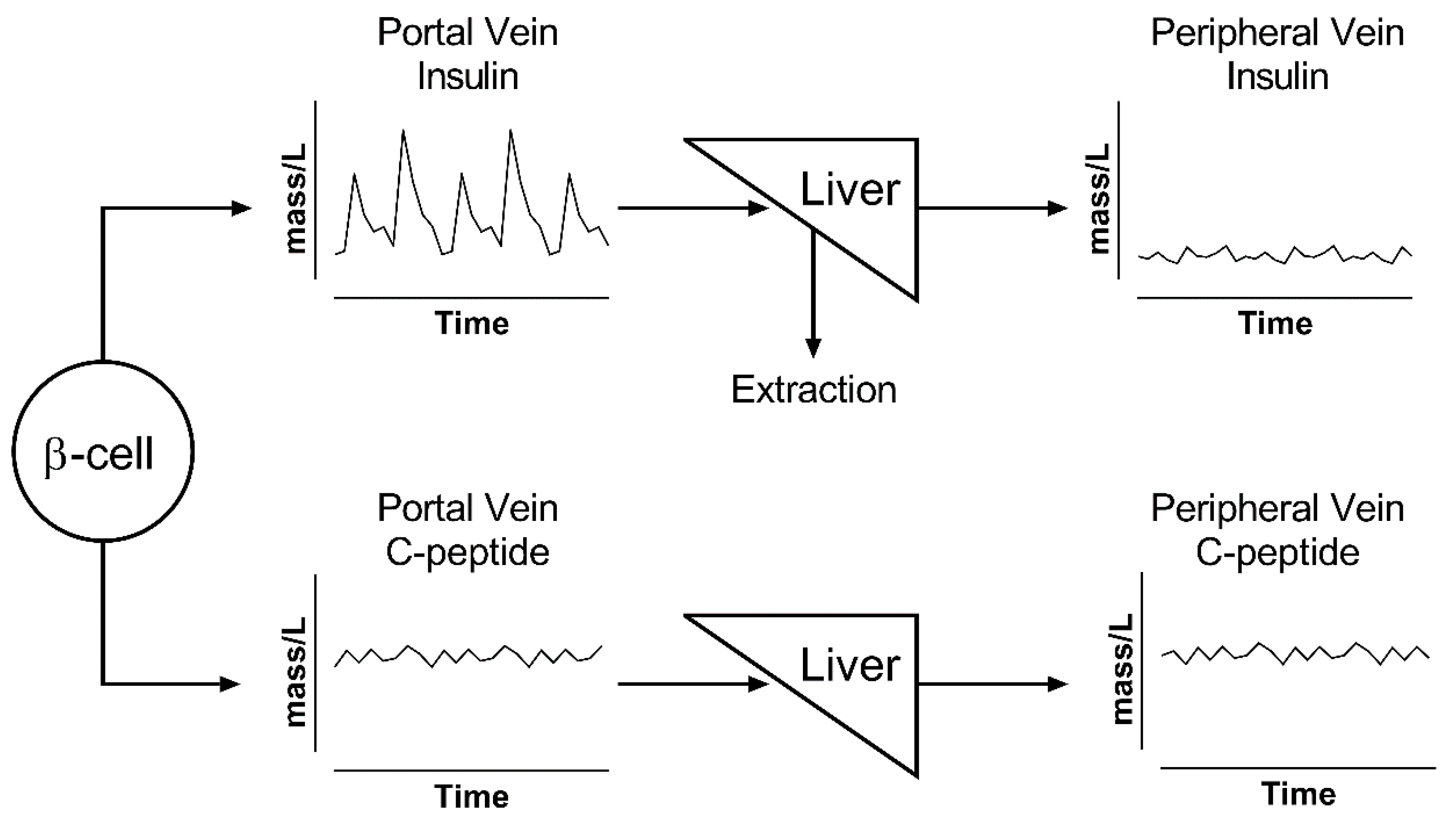
1.3. Insulin Metabolism
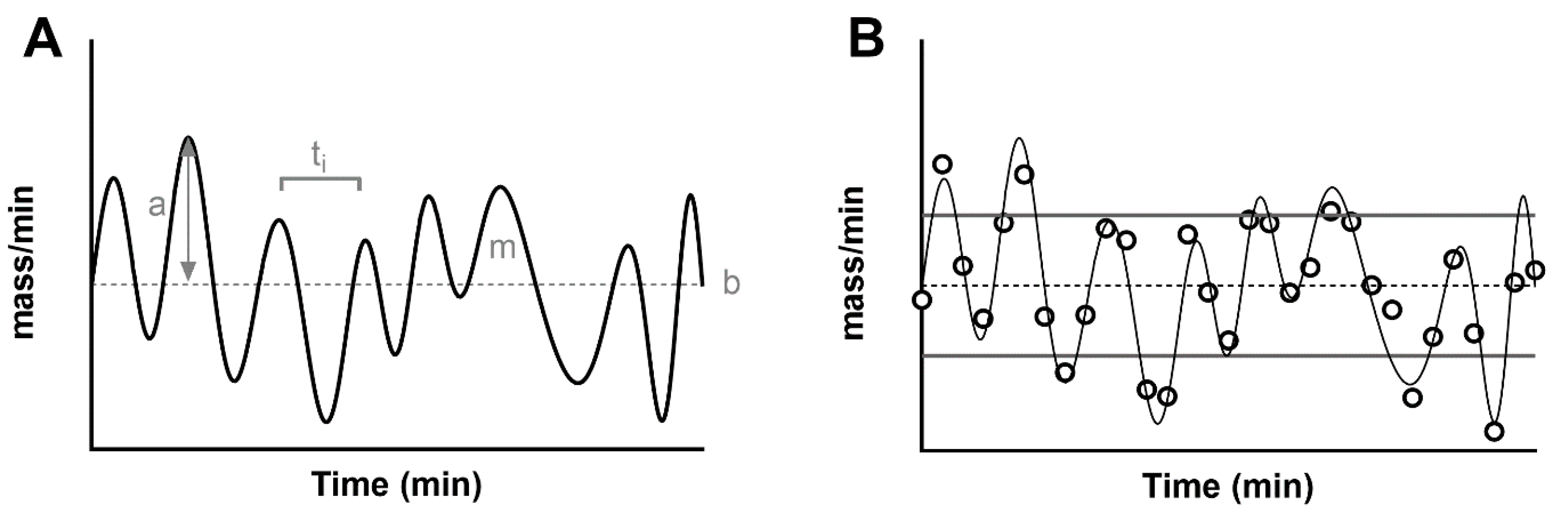
1.4. Characteristics of the Pulsatile Secretion Signal
2. Estimation of Secretion: The Deconvolution Problem
2.1. Choice of the Substrate to Measure
2.2. Choice of the Deconvolution Methodology
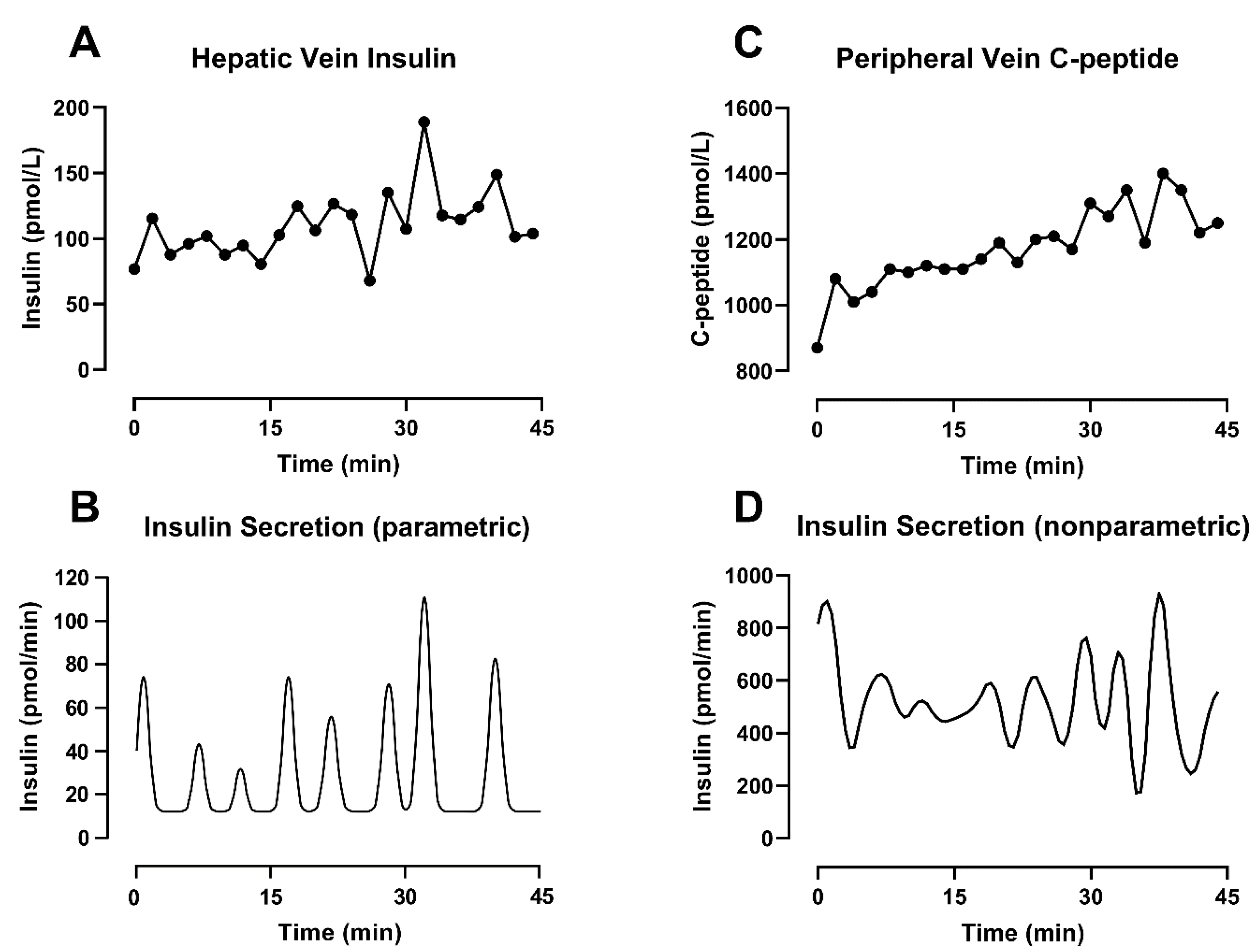
2.2.1. Parametric Deconvolution
2.2.2. Nonparametric Deconvolution
3. Summary and Future Directions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Woerle, H.J.; Meyer, C.; Dostou, J.M.; Gosmanov, N.R.; Islam, N.; Popa, E.; Wittlin, S.D.; Welle, S.L.; Gerich, J.E. Pathways for glucose disposal after meal ingestion in humans. Am. J. Physiol. Endocrinol. Metab. 2003, 284, E716–E725. [Google Scholar] [CrossRef]
- Veldhuis, J.D.; Keenan, D.M.; Pincus, S.M. Motivations and methods for analyzing pulsatile hormone secretion. Endocr. Rev. 2008, 29, 823–864. [Google Scholar] [CrossRef] [PubMed]
- Pedersen, M.G.; Bertram, R.; Sherman, A. Intra- and inter-islet synchronization of metabolically driven insulin secretion. Biophys. J. 2005, 89, 107–119. [Google Scholar] [CrossRef] [PubMed]
- Laurenti, M.C.; Dalla Man, C.; Varghese, R.T.; Andrews, J.C.; Rizza, R.A.; Matveyenko, A.; De Nicolao, G.; Cobelli, C.; Vella, A. Diabetes-associated genetic variation in TCF7L2 alters pulsatile insulin secretion in humans. JCI Insight 2020, 5, e136136. [Google Scholar] [CrossRef]
- Da Silva Xavier, G. The Cells of the Islets of Langerhans. J. Clin. Med. 2018, 7, 54. [Google Scholar] [CrossRef]
- Cabrera, O.; Berman, D.M.; Kenyon, N.S.; Ricordi, C.; Berggren, P.O.; Caicedo, A. The unique cytoarchitecture of human pancreatic islets has implications for islet cell function. Proc. Natl. Acad. Sci. USA 2006, 103, 2334–2339. [Google Scholar] [CrossRef] [PubMed]
- Kilimnik, G.; Zhao, B.; Jo, J.; Periwal, V.; Witkowski, P.; Misawa, R.; Hara, M. Altered islet composition and disproportionate loss of large islets in patients with type 2 diabetes. PLoS ONE 2011, 6, e27445. [Google Scholar] [CrossRef] [PubMed]
- Farnsworth, N.L.; Benninger, R.K. New insights into the role of connexins in pancreatic islet function and diabetes. FEBS Lett. 2014, 588, 1278–1287. [Google Scholar] [CrossRef]
- Wendt, A.; Eliasson, L. Pancreatic alpha-cells-The unsung heroes in islet function. Semin. Cell Dev. Biol. 2020, 103, 41–50. [Google Scholar] [CrossRef]
- Rodriguez-Diaz, R.; Tamayo, A.; Hara, M.; Caicedo, A. The Local Paracrine Actions of the Pancreatic alpha-Cell. Diabetes 2020, 69, 550–558. [Google Scholar] [CrossRef] [PubMed]
- Ravier, M.A.; Guldenagel, M.; Charollais, A.; Gjinovci, A.; Caille, D.; Sohl, G.; Wollheim, C.B.; Willecke, K.; Henquin, J.C.; Meda, P. Loss of connexin36 channels alters beta-cell coupling, islet synchronization of glucose-induced Ca2+ and insulin oscillations, and basal insulin release. Diabetes 2005, 54, 1798–1807. [Google Scholar] [CrossRef] [PubMed]
- Scarl, R.T.; Corbin, K.L.; Vann, N.W.; Smith, H.M.; Satin, L.S.; Sherman, A.; Nunemaker, C.S. Intact pancreatic islets and dispersed beta-cells both generate intracellular calcium oscillations but differ in their responsiveness to glucose. Cell Calcium 2019, 83, 102081. [Google Scholar] [CrossRef] [PubMed]
- Sorensen, H.; Winzell, M.S.; Brand, C.L.; Fosgerau, K.; Gelling, R.W.; Nishimura, E.; Ahren, B. Glucagon receptor knockout mice display increased insulin sensitivity and impaired beta-cell function. Diabetes 2006, 55, 3463–3469. [Google Scholar] [CrossRef]
- Melloul, D.; Marshak, S.; Cerasi, E. Regulation of insulin gene transcription. Diabetologia 2002, 45, 309–326. [Google Scholar] [CrossRef]
- Andrali, S.S.; Sampley, M.L.; Vanderford, N.L.; Ozcan, S. Glucose regulation of insulin gene expression in pancreatic beta-cells. Biochem. J. 2008, 415, 1–10. [Google Scholar] [CrossRef]
- Ghiasi, S.M.; Dahlby, T.; Hede Andersen, C.; Haataja, L.; Petersen, S.; Omar-Hmeadi, M.; Yang, M.; Pihl, C.; Bresson, S.E.; Khilji, M.S.; et al. Endoplasmic Reticulum Chaperone Glucose-Regulated Protein 94 Is Essential for Proinsulin Handling. Diabetes 2019, 68, 747–760. [Google Scholar] [CrossRef] [PubMed]
- Greenman, I.C.; Gomez, E.; Moore, C.E.; Herbert, T.P. The selective recruitment of mRNA to the ER and an increase in initiation are important for glucose-stimulated proinsulin synthesis in pancreatic beta-cells. Biochem. J. 2005, 391, 291–300. [Google Scholar] [CrossRef]
- Arvan, P.; Halban, P.A. Sorting ourselves out: Seeking consensus on trafficking in the beta-cell. Traffic 2004, 5, 53–61. [Google Scholar] [CrossRef]
- Ramzy, A.; Asadi, A.; Kieffer, T.J. Revisiting Proinsulin Processing: Evidence That Human beta-Cells Process Proinsulin With Prohormone Convertase (PC) 1/3 but Not PC2. Diabetes 2020, 69, 1451–1462. [Google Scholar] [CrossRef]
- Bratanova-Tochkova, T.K.; Cheng, H.; Daniel, S.; Gunawardana, S.; Liu, Y.J.; Mulvaney-Musa, J.; Schermerhorn, T.; Straub, S.G.; Yajima, H.; Sharp, G.W. Triggering and augmentation mechanisms, granule pools, and biphasic insulin secretion. Diabetes 2002, 51 (Suppl. S1), S83–S90. [Google Scholar] [CrossRef]
- McKenna, J.P.; Ha, J.; Merrins, M.J.; Satin, L.S.; Sherman, A.; Bertram, R. Ca2+ Effects on ATP Production and Consumption Have Regulatory Roles on Oscillatory Islet Activity. Biophys. J. 2016, 110, 733–742. [Google Scholar] [CrossRef] [PubMed]
- Newsholme, P.; Cruzat, V.; Arfuso, F.; Keane, K. Nutrient regulation of insulin secretion and action. J. Endocrinol. 2014, 221, R105–R120. [Google Scholar] [CrossRef]
- Olofsson, C.S.; Gopel, S.O.; Barg, S.; Galvanovskis, J.; Ma, X.; Salehi, A.; Rorsman, P.; Eliasson, L. Fast insulin secretion reflects exocytosis of docked granules in mouse pancreatic B-cells. Pflug. Arch. 2002, 444, 43–51. [Google Scholar] [CrossRef]
- Zhu, X.; Hu, R.; Brissova, M.; Stein, R.W.; Powers, A.C.; Gu, G.; Kaverina, I. Microtubules Negatively Regulate Insulin Secretion in Pancreatic beta Cells. Dev. Cell 2015, 34, 656–668. [Google Scholar] [CrossRef]
- Rorsman, P.; Renstrom, E. Insulin granule dynamics in pancreatic beta cells. Diabetologia 2003, 46, 1029–1045. [Google Scholar] [CrossRef] [PubMed]
- Benninger, R.K.; Piston, D.W. Cellular communication and heterogeneity in pancreatic islet insulin secretion dynamics. Trends Endocrinol. Metab. 2014, 25, 399–406. [Google Scholar] [CrossRef] [PubMed]
- Bertram, R.; Satin, L.S.; Sherman, A.S. Closing in on the Mechanisms of Pulsatile Insulin Secretion. Diabetes 2018, 67, 351–359. [Google Scholar] [CrossRef]
- Fendler, B.; Zhang, M.; Satin, L.; Bertram, R. Synchronization of pancreatic islet oscillations by intrapancreatic ganglia: A modeling study. Biophys. J. 2009, 97, 722–729. [Google Scholar] [CrossRef]
- Zhang, M.; Fendler, B.; Peercy, B.; Goel, P.; Bertram, R.; Sherman, A.; Satin, L. Long lasting synchronization of calcium oscillations by cholinergic stimulation in isolated pancreatic islets. Biophys. J. 2008, 95, 4676–4688. [Google Scholar] [CrossRef] [PubMed]
- Johnston, N.R.; Mitchell, R.K.; Haythorne, E.; Pessoa, M.P.; Semplici, F.; Ferrer, J.; Piemonti, L.; Marchetti, P.; Bugliani, M.; Bosco, D.; et al. Beta Cell Hubs Dictate Pancreatic Islet Responses to Glucose. Cell Metab. 2016, 24, 389–401. [Google Scholar] [CrossRef]
- Salem, V.; Silva, L.D.; Suba, K.; Georgiadou, E.; Neda Mousavy Gharavy, S.; Akhtar, N.; Martin-Alonso, A.; Gaboriau, D.C.A.; Rothery, S.M.; Stylianides, T.; et al. Leader beta-cells coordinate Ca(2+) dynamics across pancreatic islets in vivo. Nat. Metab. 2019, 1, 615–629. [Google Scholar] [CrossRef] [PubMed]
- Satin, L.S.; Zhang, Q.; Rorsman, P. “Take Me To Your Leader”: An Electrophysiological Appraisal of the Role of Hub Cells in Pancreatic Islets. Diabetes 2020, 69, 830–836. [Google Scholar] [CrossRef] [PubMed]
- Gheibi, S.; Ghasemi, A. Insulin secretion: The nitric oxide controversy. EXCLI J. 2020, 19, 1227–1245. [Google Scholar] [CrossRef] [PubMed]
- Menegaz, D.; Hagan, D.W.; Almaca, J.; Cianciaruso, C.; Rodriguez-Diaz, R.; Molina, J.; Dolan, R.M.; Becker, M.W.; Schwalie, P.C.; Nano, R.; et al. Mechanism and effects of pulsatile GABA secretion from cytosolic pools in the human beta cell. Nat. Metab. 2019, 1, 1110–1126. [Google Scholar] [CrossRef]
- Rahman, F.U.; Park, D.R.; Joe, Y.; Jang, K.Y.; Chung, H.T.; Kim, U.H. Critical Roles of Carbon Monoxide and Nitric Oxide in Ca(2+) Signaling for Insulin Secretion in Pancreatic Islets. Antioxid. Redox Signal. 2019, 30, 560–576. [Google Scholar] [CrossRef] [PubMed]
- Meier, J.J.; Veldhuis, J.D.; Butler, P.C. Pulsatile insulin secretion dictates systemic insulin delivery by regulating hepatic insulin extraction in humans. Diabetes 2005, 54, 1649–1656. [Google Scholar] [CrossRef]
- Sathananthan, A.; Dalla Man, C.; Zinsmeister, A.R.; Camilleri, M.; Rodeheffer, R.J.; Toffolo, G.; Cobelli, C.; Rizza, R.A.; Vella, A. A concerted decline in insulin secretion and action occurs across the spectrum of fasting and postchallenge glucose concentrations. Clin. Endocrinol. 2012, 76, 212–219. [Google Scholar] [CrossRef] [PubMed]
- Hunt, N.J.; Lockwood, G.P.; Warren, A.; Mao, H.; McCourt, P.A.G.; Le Couteur, D.G.; Cogger, V.C. Manipulating fenestrations in young and old liver sinusoidal endothelial cells. Am. J. Physiol. Gastrointest. Liver Physiol. 2019, 316, G144–G154. [Google Scholar] [CrossRef]
- Najjar, S.M.; Perdomo, G. Hepatic Insulin Clearance: Mechanism and Physiology. Physiology 2019, 34, 198–215. [Google Scholar] [CrossRef]
- Porksen, N.; Hollingdal, M.; Juhl, C.; Butler, P.; Veldhuis, J.D.; Schmitz, O. Pulsatile insulin secretion: Detection, regulation, and role in diabetes. Diabetes 2002, 51 (Suppl. S1), S245–S254. [Google Scholar] [CrossRef] [PubMed]
- Piccinini, F.; Bergman, R.N. The Measurement of Insulin Clearance. Diabetes Care 2020, 43, 2296–2302. [Google Scholar] [CrossRef]
- Song, S.H.; McIntyre, S.S.; Shah, H.; Veldhuis, J.D.; Hayes, P.C.; Butler, P.C. Direct measurement of pulsatile insulin secretion from the portal vein in human subjects. J. Clin. Endocrinol. Metab. 2000, 85, 4491–4499. [Google Scholar] [CrossRef]
- Laurenti, M.C.; Vella, A.; Varghese, R.T.; Andrews, J.C.; Sharma, A.; Kittah, N.E.; Rizza, R.A.; Matveyenko, A.; De Nicolao, G.; Cobelli, C.; et al. Assessment of pulsatile insulin secretion derived from peripheral plasma C-peptide concentrations by nonparametric stochastic deconvolution. Am. J. Physiol. Endocrinol. Metab. 2019, 316, E687–E694. [Google Scholar] [CrossRef] [PubMed]
- Kaneko, K.; Soty, M.; Zitoun, C.; Duchampt, A.; Silva, M.; Philippe, E.; Gautier-Stein, A.; Rajas, F.; Mithieux, G. The role of kidney in the inter-organ coordination of endogenous glucose production during fasting. Mol. Metab. 2018, 16, 203–212. [Google Scholar] [CrossRef] [PubMed]
- Lewis, G.F.; Carpentier, A.C.; Pereira, S.; Hahn, M.; Giacca, A. Direct and indirect control of hepatic glucose production by insulin. Cell Metab. 2021, 33, 709–720. [Google Scholar] [CrossRef] [PubMed]
- Dimitriadis, G.; Mitrou, P.; Lambadiari, V.; Maratou, E.; Raptis, S.A. Insulin effects in muscle and adipose tissue. Diabetes Res. Clin. Pract. 2011, 93 (Suppl. S1), S52–S59. [Google Scholar] [CrossRef]
- Nozaki, Y.; Petersen, M.C.; Zhang, D.; Vatner, D.F.; Perry, R.J.; Abulizi, A.; Haedersdal, S.; Zhang, X.-M.; Butrico, G.M.; Samuel, V.T.; et al. Metabolic control analysis of hepatic glycogen synthesis in vivo. Proc. Natl. Acad. Sci. USA 2020, 117, 8166–8176. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, C.S.; Begg, D.P. The regulation of food intake by insulin in the central nervous system. J. Neuroendocrinol. 2021, 33, e12952. [Google Scholar] [CrossRef] [PubMed]
- Sparacino, G.; Cobelli, C. Impulse response model in reconstruction of insulin secretion by deconvolution: Role of input design in the identification experiment. Ann. Biomed. Eng. 1997, 25, 398–416. [Google Scholar] [CrossRef]
- Simon, C.; Brandenberger, G. Ultradian oscillations of insulin secretion in humans. Diabetes 2002, 51 (Suppl. S1), S258–S261. [Google Scholar] [CrossRef] [PubMed]
- Lang, D.A.; Matthews, D.R.; Peto, J.; Turner, R.C. Cyclic oscillations of basal plasma glucose and insulin concentrations in human beings. N. Engl. J. Med. 1979, 301, 1023–1027. [Google Scholar] [CrossRef]
- Matveyenko, A.V.; Liuwantara, D.; Gurlo, T.; Kirakossian, D.; Dalla Man, C.; Cobelli, C.; White, M.F.; Copps, K.D.; Volpi, E.; Fujita, S.; et al. Pulsatile portal vein insulin delivery enhances hepatic insulin action and signaling. Diabetes 2012, 61, 2269–2279. [Google Scholar] [CrossRef]
- Porksen, N.; Nyholm, B.; Veldhuis, J.D.; Butler, P.C.; Schmitz, O. In humans at least 75% of insulin secretion arises from punctuated insulin secretory bursts. Am. J. Physiol. 1997, 273, E908–E914. [Google Scholar] [CrossRef]
- Kushler, R.H.; Brown, M.B. A model for the identification of hormone pulses. Stat. Med. 1991, 10, 329–340. [Google Scholar] [CrossRef]
- Vidal, A.; Zhang, Q.; Medigue, C.; Fabre, S.; Clement, F. DynPeak: An algorithm for pulse detection and frequency analysis in hormonal time series. PLoS ONE 2012, 7, e39001. [Google Scholar] [CrossRef] [PubMed]
- Horton, K.W.; Carlson, N.E.; Grunwald, G.K.; Mulvahill, M.J.; Polotsky, A.J. A population-based approach to analyzing pulses in time series of hormone data. Stat. Med. 2017, 36, 2576–2589. [Google Scholar] [CrossRef]
- Campioni, M.; Toffolo, G.; Basu, R.; Rizza, R.A.; Cobelli, C. Minimal model assessment of hepatic insulin extraction during an oral test from standard insulin kinetic parameters. Am. J. Physiol. Endocrinol. Metab. 2009, 297, E941–E948. [Google Scholar] [CrossRef]
- Van Cauter, E.; Mestrez, F.; Sturis, J.; Polonsky, K.S. Estimation of insulin secretion rates from C-peptide levels. Comparison of individual and standard kinetic parameters for C-peptide clearance. Diabetes 1992, 41, 368–377. [Google Scholar] [CrossRef]
- Varghese, R.T.; Dalla Man, C.; Laurenti, M.C.; Piccinini, F.; Sharma, A.; Shah, M.; Bailey, K.R.; Rizza, R.A.; Cobelli, C.; Vella, A. Performance of individually measured vs population-based C-peptide kinetics to assess beta-cell function in the presence and absence of acute insulin resistance. Diabetes Obes. Metab. 2018, 20, 549–555. [Google Scholar] [CrossRef]
- Laedtke, T.; Kjems, L.; Porksen, N.; Schmitz, O.; Veldhuis, J.; Kao, P.C.; Butler, P.C. Overnight inhibition of insulin secretion restores pulsatility and proinsulin/insulin ratio in type 2 diabetes. Am. J. Physiol. Endocrinol. Metab. 2000, 279, E520–E528. [Google Scholar] [CrossRef]
- Porksen, N. Early changes in beta-cell function and insulin pulsatility as predictors for type 2 diabetes. Diabetes Nutr. Metab. 2002, 15, 9–14. [Google Scholar] [PubMed]
- Meier, J.J.; Pennartz, C.; Schenker, N.; Menge, B.A.; Schmidt, W.E.; Heise, T.; Kapitza, C.; Veldhuis, J.D. Hyperglycaemia is associated with impaired pulsatile insulin secretion: Effect of basal insulin therapy. Diabetes Obes. Metab. 2013, 15, 258–263. [Google Scholar] [CrossRef]
- Meneilly, G.S.; Veldhuis, J.D.; Elahi, D. Deconvolution analysis of rapid insulin pulses before and after six weeks of continuous subcutaneous administration of glucagon-like peptide-1 in elderly patients with type 2 diabetes. J. Clin. Endocrinol. Metab. 2005, 90, 6251–6256. [Google Scholar] [CrossRef][Green Version]
- Sparacino, G.; Cobelli, C. A stochastic deconvolution method to reconstruct insulin secretion rate after a glucose stimulus. IEEE Trans. Biomed. Eng. 1996, 43, 512–529. [Google Scholar] [CrossRef]
- Cobelli, C.; Dalla Man, C.; Toffolo, G.; Basu, R.; Vella, A.; Rizza, R. The oral minimal model method. Diabetes 2014, 63, 1203–1213. [Google Scholar] [CrossRef] [PubMed]
- Balks, H.J.; Schmidt, A.; Prank, K.; Hemmer, F.; von zur Muhlen, A.; Brabant, G. Temporal pattern of pancreatic insulin and C-peptide secretion and of plasma glucose levels after nutritional stimulation. J. Clin. Endocrinol. Metab. 1992, 75, 1198–1203. [Google Scholar] [CrossRef]
- De Nicolao, G.; Sparacino, G.; Cobelli, C. Nonparametric input estimation in physiological systems: Problems, methods, and case studies. Automatica 1997, 33, 851–870. [Google Scholar] [CrossRef]
- Bertero, M. Linear Inverse and III-Posed Problems. In Advances in Electronics and Electron Physics; Hawkes, P.W., Ed.; Academic Press: Cambridge, MA, USA, 1989; Volume 75, pp. 1–120. [Google Scholar]
- Carlson, N.E.; Johnson, T.D.; Brown, M.B. A Bayesian approach to modeling associations between pulsatile hormones. Biometrics 2009, 65, 650–659. [Google Scholar] [CrossRef]
- Johnson, M.L.; Pipes, L.; Veldhuis, P.P.; Farhy, L.S.; Boyd, D.G.; Evans, W.S. AutoDecon, a deconvolution algorithm for identification and characterization of luteinizing hormone secretory bursts: Description and validation using synthetic data. Anal. Biochem. 2008, 381, 8–17. [Google Scholar] [CrossRef][Green Version]
- Phillips, D.L. A Technique for the Numerical Solution of Certain Integral Equations of the First Kind. J. ACM 1962, 9, 84–97. [Google Scholar] [CrossRef]
- Tikhonov, A. Solution of Incorrectly Formulated Problems and the Regularization Method. Sov. Math. 1963, 4, 1035–1038. [Google Scholar]
- Krawczyk-Stańdo, D.; Rudnicki, M. Regularization Parameter Selection in Discrete Ill-Posed Problems—The Use of the U-Curve. Int. J. Appl. Math. Comput. Sci. 2007, 17, 157–164. [Google Scholar] [CrossRef]
- Karoui, A.; Bear, L.; Migerditichan, P.; Zemzemi, N. Evaluation of Fifteen Algorithms for the Resolution of the Electrocardiography Imaging Inverse Problem Using ex-vivo and in-silico Data. Front. Physiol. 2018, 9, 1708. [Google Scholar] [CrossRef] [PubMed]
- Matthews, D.R.; Naylor, B.A.; Jones, R.G.; Ward, G.M.; Turner, R.C. Pulsatile insulin has greater hypoglycemic effect than continuous delivery. Diabetes 1983, 32, 617–621. [Google Scholar] [CrossRef] [PubMed]
- Sturis, J.; Scheen, A.J.; Leproult, R.; Polonsky, K.S.; van Cauter, E. 24-hour glucose profiles during continuous or oscillatory insulin infusion. Demonstration of the functional significance of ultradian insulin oscillations. J. Clin. Investig. 1995, 95, 1464–1471. [Google Scholar] [CrossRef]
- Juhl, C.B.; Gjedsted, J.; Nielsen, M.F.; Schmitz, O. Increased action of pulsatile compared to non-pulsatile insulin delivery during a meal-like glucose exposure simulated by computerized infusion in healthy humans. Metabolism 2012, 61, 1177–1181. [Google Scholar] [CrossRef]
- Komjati, M.; Bratusch-Marrain, P.; Waldhausl, W. Superior efficacy of pulsatile versus continuous hormone exposure on hepatic glucose production in vitro. Endocrinology 1986, 118, 312–319. [Google Scholar] [CrossRef] [PubMed]
- Zhao, G.; Wirth, D.; Schmitz, I.; Meyer-Hermann, M. A mathematical model of the impact of insulin secretion dynamics on selective hepatic insulin resistance. Nat. Commun. 2017, 8, 1362. [Google Scholar] [CrossRef] [PubMed]
- Satin, L.S.; Butler, P.C.; Ha, J.; Sherman, A.S. Pulsatile insulin secretion, impaired glucose tolerance and type 2 diabetes. Mol. Aspects Med. 2015, 42, 61–77. [Google Scholar] [CrossRef] [PubMed]
- Schofield, C.J.; Sutherland, C. Disordered insulin secretion in the development of insulin resistance and Type 2 diabetes. Diabet. Med. 2012, 29, 972–979. [Google Scholar] [CrossRef] [PubMed]
- Goodner, C.J.; Koerker, D.J.; Weigle, D.S.; McCulloch, D.K. Decreased insulin- and glucagon-pulse amplitude accompanying beta-cell deficiency induced by streptozocin in baboons. Diabetes 1989, 38, 925–931. [Google Scholar] [CrossRef] [PubMed]
- O’Rahilly, S.; Turner, R.C.; Matthews, D.R. Impaired pulsatile secretion of insulin in relatives of patients with non-insulin-dependent diabetes. N. Engl. J. Med. 1988, 318, 1225–1230. [Google Scholar] [CrossRef] [PubMed]
- Westacott, M.J.; Farnsworth, N.L.; St Clair, J.R.; Poffenberger, G.; Heintz, A.; Ludin, N.W.; Hart, N.J.; Powers, A.C.; Benninger, R.K.P. Age-Dependent Decline in the Coordinated [Ca(2+)] and Insulin Secretory Dynamics in Human Pancreatic Islets. Diabetes 2017, 66, 2436–2445. [Google Scholar] [CrossRef] [PubMed]
- Erion, K.; Corkey, B.E. beta-Cell Failure or beta-Cell Abuse? Front. Endocrinol. 2018, 9, 532. [Google Scholar] [CrossRef]
- Corbin, K.L.; Waters, C.D.; Shaffer, B.K.; Verrilli, G.M.; Nunemaker, C.S. Islet Hypersensitivity to Glucose Is Associated With Disrupted Oscillations and Increased Impact of Proinflammatory Cytokines in Islets From Diabetes-Prone Male Mice. Endocrinology 2016, 157, 1826–1838. [Google Scholar] [CrossRef] [PubMed]
- Laurenti, M.C.; Dalla Man, C.; Varghese, R.T.; Andrews, J.C.; Jones, J.G.; Barosa, C.; Rizza, R.A.; Matveyenko, A.; De Nicolao, G.; Bailey, K.R.; et al. Insulin Pulse Characteristics and Insulin Action in Non-diabetic Humans. J. Clin. Endocrinol. Metab. 2021, 106, 1702–1709. [Google Scholar] [CrossRef] [PubMed]
- Pincus, S. Approximate entropy (ApEn) as a complexity measure. Chaos 1995, 5, 110–117. [Google Scholar] [CrossRef] [PubMed]
- Menge, B.A.; Gruber, L.; Jorgensen, S.M.; Deacon, C.F.; Schmidt, W.E.; Veldhuis, J.D.; Holst, J.J.; Meier, J.J. Loss of inverse relationship between pulsatile insulin and glucagon secretion in patients with type 2 diabetes. Diabetes 2011, 60, 2160–2168. [Google Scholar] [CrossRef] [PubMed]
- Prieto, T.E.; Myklebust, J.B.; Hoffmann, R.G.; Lovett, E.G.; Myklebust, B.M. Measures of postural steadiness: Differences between healthy young and elderly adults. IEEE Trans. Biomed. Eng. 1996, 43, 956–966. [Google Scholar] [CrossRef]
- Brown, R.J.; Rother, K.I. Effects of beta-cell rest on beta-cell function: A review of clinical and preclinical data. Pediatr. Diabetes 2008, 9, 14–22. [Google Scholar] [CrossRef]
- Corezola do Amaral, M.E.; Kravets, V.; Dwulet, J.M.; Farnsworth, N.L.; Piscopio, R.; Schleicher, W.E.; Miranda, J.G.; Benninger, R.K.P. Caloric restriction recovers impaired beta-cell-beta-cell gap junction coupling, calcium oscillation coordination, and insulin secretion in prediabetic mice. Am. J. Physiol. Endocrinol. Metab. 2020, 319, E709–E720. [Google Scholar] [CrossRef] [PubMed]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Laurenti, M.C.; Matveyenko, A.; Vella, A. Measurement of Pulsatile Insulin Secretion: Rationale and Methodology. Metabolites 2021, 11, 409. https://doi.org/10.3390/metabo11070409
Laurenti MC, Matveyenko A, Vella A. Measurement of Pulsatile Insulin Secretion: Rationale and Methodology. Metabolites. 2021; 11(7):409. https://doi.org/10.3390/metabo11070409
Chicago/Turabian StyleLaurenti, Marcello C., Aleksey Matveyenko, and Adrian Vella. 2021. "Measurement of Pulsatile Insulin Secretion: Rationale and Methodology" Metabolites 11, no. 7: 409. https://doi.org/10.3390/metabo11070409
APA StyleLaurenti, M. C., Matveyenko, A., & Vella, A. (2021). Measurement of Pulsatile Insulin Secretion: Rationale and Methodology. Metabolites, 11(7), 409. https://doi.org/10.3390/metabo11070409