
Identification of Koala (Phascolarctos cinereus) Faecal Cortisol Metabolites Using Liquid Chromatography-Mass Spectrometry and Enzyme Immunoassays

 , ,
, ,  , and
, and 
Abstract
1. Introduction
2. Results
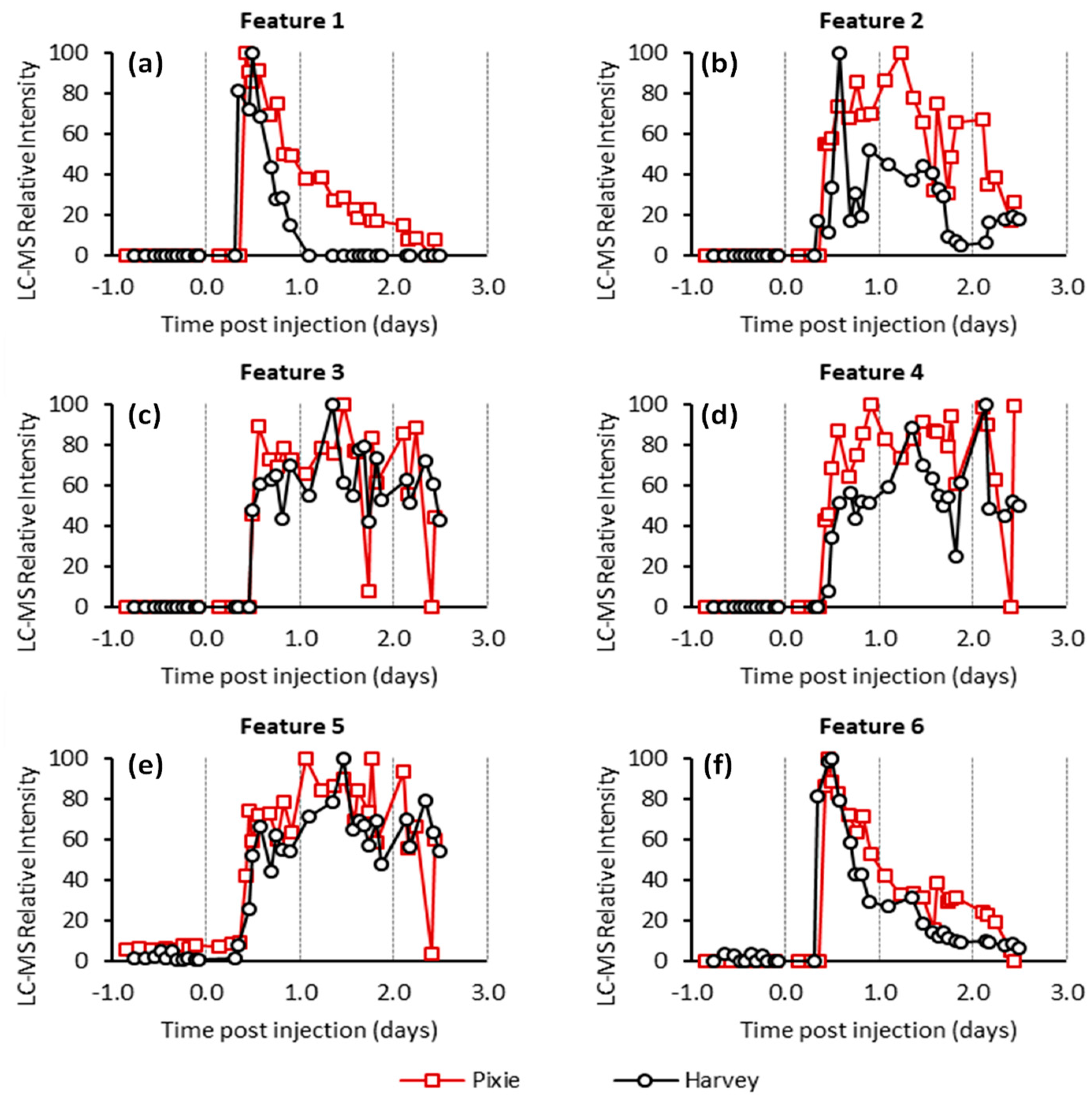
2.1. Effect of Blood Sampling and Injection
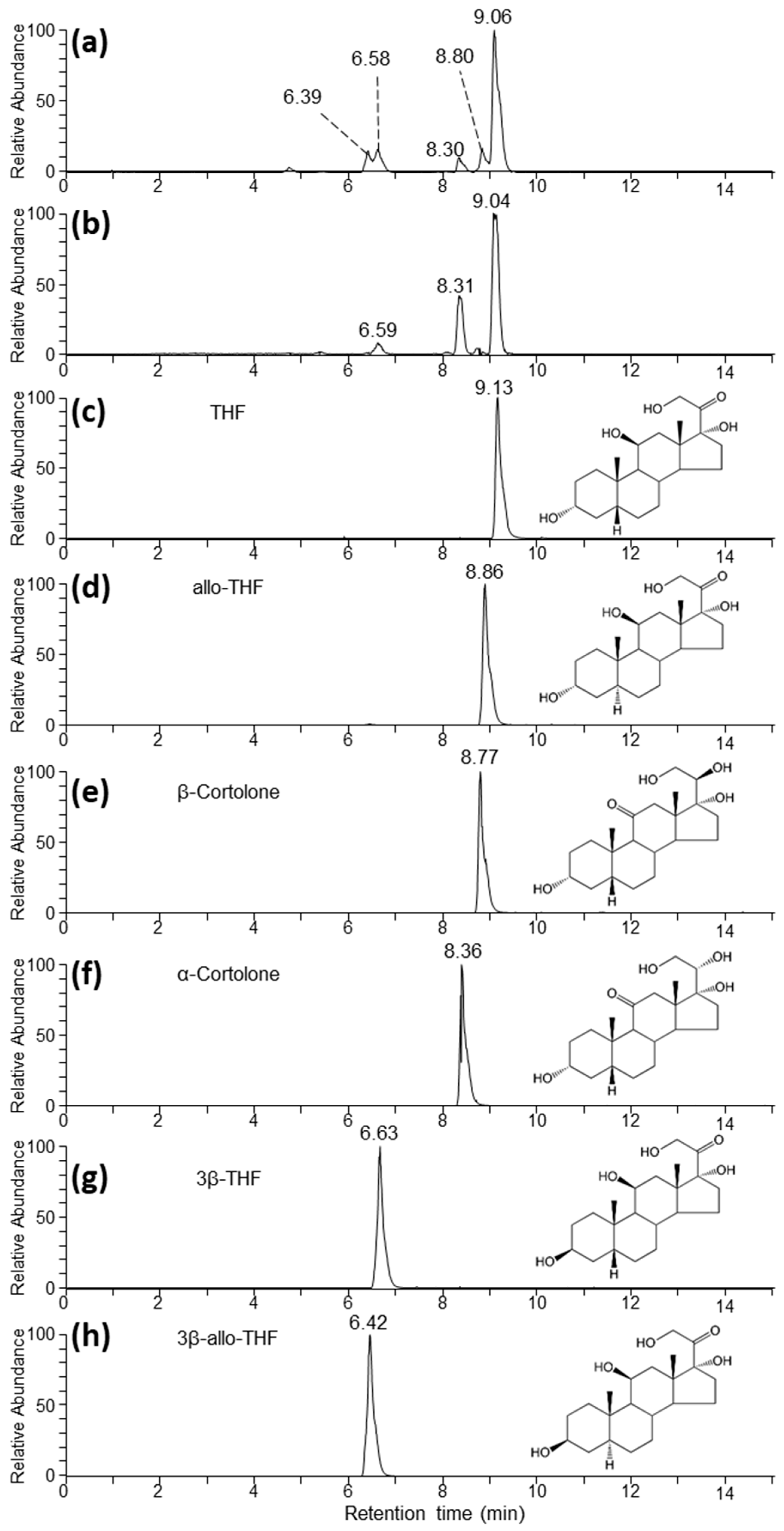
2.2. Untargeted LC-MS Analysis
2.3. Enzyme Immunoassay
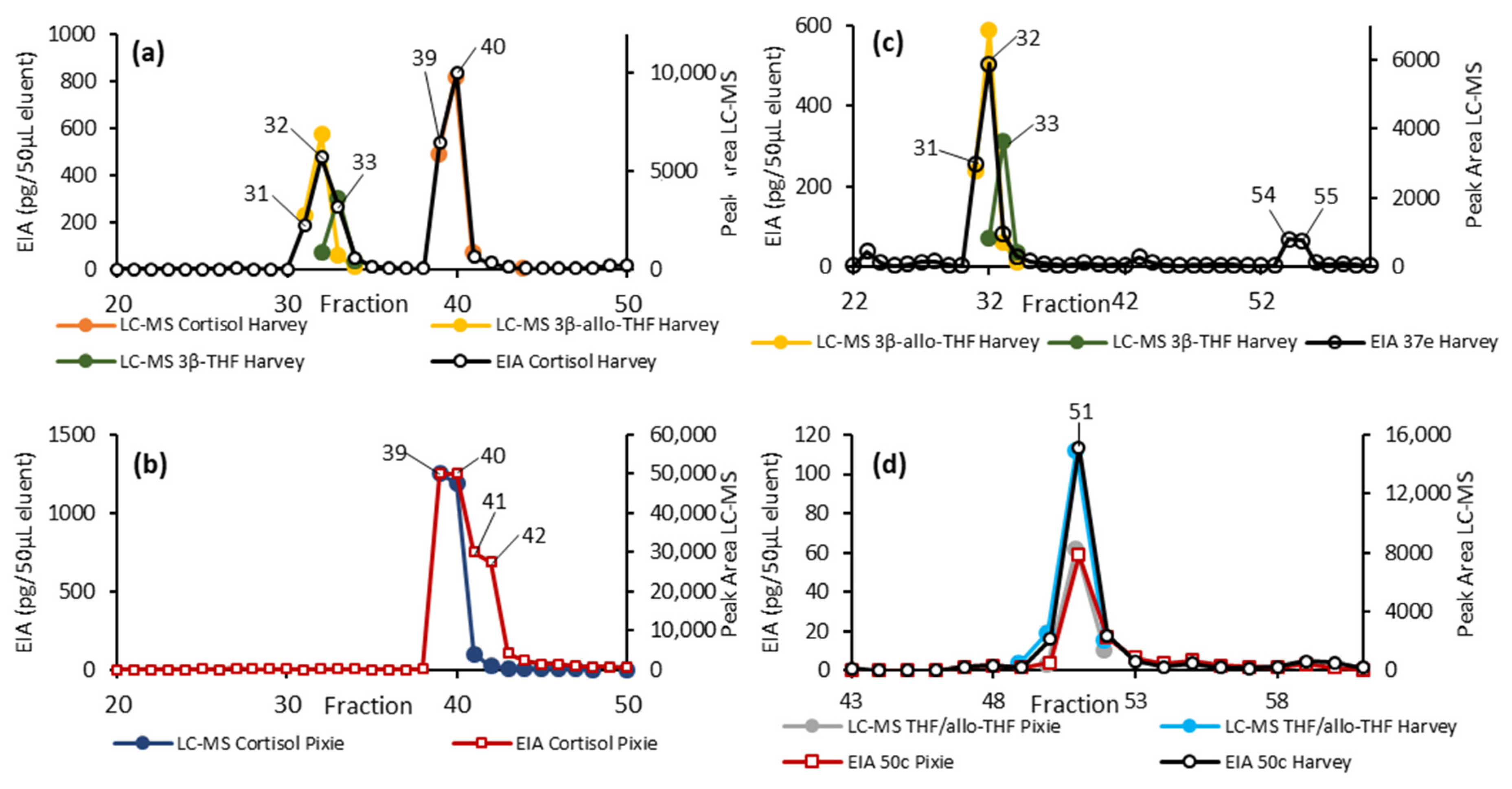
2.4. EIA and LC-MS Analysis of Fractionated Pooled Samples
3. Discussion
4. Materials and Methods
4.1. Koalas
4.2. Trial Design and Collection of Faecal Samples
4.3. Sample Preparation
4.4. EIA Analysis
4.5. Standards
4.6. HPLC Fractionation of Pooled Samples
4.7. HPLC-MS Analysis
4.8. Processing of the Untargeted HILIC LC-MS Data
4.9. Ethics
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Department of Agriculture Water and the Environment. Koala Populations in Queensland, New South Wales and the Australian Capital Territory and National Environment Law. Available online: http://www.environment.gov.au/biodiversity/threatened/species/koala. (accessed on 7 July 2020).
- Department of Environment and Science. Koala Threats. Available online: https://www.ehp.qld.gov.au/wildlife/koalas/koala-threats.html (accessed on 15 June 2020).
- Timmermans, S.; Souffriau, J.; Libert, C. A general introduction to glucocorticoid biology. Front. Immunol. 2019, 10, 1545. [Google Scholar] [CrossRef]
- Pruett, S.B. Stress and the immune system. Pathophysiology 2003, 9, 133–153. [Google Scholar] [CrossRef]
- Touma, C.; Palme, R. Measuring fecal glucocorticoid metabolites in mammals and birds: The importance of validation. Ann. N. Y. Acad. Sci. 2005, 1046, 54–74. [Google Scholar] [CrossRef] [PubMed]
- Schimmer, B.P.; Funder, J.W. ACTH, Adrenal Steroids, and Pharmacology of the Adrenal Cortex. In Goodman & Gilman’s: The Pharmacological Basis of Therapeutics; The McGraw-Hill Companies, Inc.: New York, NY, USA, 2011; pp. 1209–1236. [Google Scholar]
- Knox, B.; Ladiges, P.; Evans, B.; Saint, R. Biology: An Australian Focus; McGraw-Hill Education: North Ryde, Australia, 2014. [Google Scholar]
- Urry, L.A. Campbell Biology, 11th ed.; Australian and New Zealand Version; Pearson Australia: Melbourne, VIC, Australia, 2017. [Google Scholar]
- Hogan, L.A.; Johnston, S.D.; Lisle, A.T.; Keeley, T.; Wong, P.; Nicolson, V.; Horsup, A.B.; Janssen, T.; Phillips, C.J. Behavioural and physiological responses of captive wombats (Lasiorhinus latifrons) to regular handling by humans. Appl. Anim. Behav. Sci. 2011, 134, 217–228. [Google Scholar] [CrossRef]
- Fanson, K.V.; Best, E.C.; Bunce, A.; Fanson, B.G.; Hogan, L.A.; Keeley, T.; Narayan, E.J.; Palme, R.; Parrott, M.L.; Sharp, T.M. One size does not fit all: Monitoring faecal glucocorticoid metabolites in marsupials. Gen. Comp. Endocrinol. 2017, 244, 146–156. [Google Scholar] [CrossRef] [PubMed]
- Davies, N.A.; Gramotnev, G.; McAlpine, C.; Seabrook, L.; Baxter, G.; Lunney, D.; Rhodes, J.R.; Bradley, A. Physiological stress in koala populations near the arid edge of their distribution. PLoS ONE 2013, 8, e79136. [Google Scholar] [CrossRef] [PubMed]
- Hogan, L.A.; Lisle, A.T.; Johnston, S.D.; Robertson, H. Non-invasive assessment of stress in captive numbats, Myrmecobius fasciatus (Mammalia: Marsupialia), using faecal cortisol measurement. Gen. Comp. Endocrinol. 2012, 179, 376–383. [Google Scholar] [CrossRef]
- Möstl, E.; Maggs, J.; Schrötter, G.; Besenfelder, U.; Palme, R. Measurement of cortisol metabolites in faeces of ruminants. Vet. Res. Commun. 2002, 26, 127–139. [Google Scholar] [CrossRef] [PubMed]
- Palme, R. Non-invasive measurement of glucocorticoids: Advances and problems. Physiol. Behav. 2019, 199, 229–243. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Mota, R.; Valdespino, C.; Rebolledo, J.A.R.; Palme, R. Determination of fecal glucocorticoid metabolites to evaluate stress response in Alouatta pigra. Int. J. Primatol. 2008, 29, 1365–1373. [Google Scholar] [CrossRef]
- Sheriff, M.J.; Dantzer, B.; Delehanty, B.; Palme, R.; Boonstra, R. Measuring stress in wildlife: Techniques for quantifying glucocorticoids. Oecologia 2011, 166, 869–887. [Google Scholar] [CrossRef] [PubMed]
- Davies, N.; Gillett, A.; McAlpine, C.; Seabrook, L.; Baxter, G.; Lunney, D.; Bradley, A. The effect of ACTH upon faecal glucocorticoid excretion in the koala. J. Endocrinol. 2013, 219, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Kersey, D.C.; Dehnhard, M. The use of noninvasive and minimally invasive methods in endocrinology for threatened mammalian species conservation. Gen. Comp. Endocrinol. 2014, 203, 296–306. [Google Scholar] [CrossRef] [PubMed]
- Keegan, M.T. Endocrine pharmacology. In Pharmacology and Physiology for Anesthesia; Elsevier: Amsterdam, The Netherlands, 2019; pp. 708–731. [Google Scholar]
- Walker, B.R.; Seckl, J.R. Cortisol Metabolism. In International Textbook of Obesity; John Wiley and Sons: Chichester, UK, 2001; pp. 241–268. [Google Scholar] [CrossRef]
- Palme, R.; Rettenbacher, S.; Touma, C.; El-Bahr, S.; Möstl, E. Stress hormones in mammals and birds: Comparative aspects regarding metabolism, excretion, and noninvasive measurement in fecal samples. Ann. N. Y. Acad. Sci. 2005, 1040, 162–171. [Google Scholar] [CrossRef]
- Morris, D.J.; Ridlon, J.M. Glucocorticoids and gut bacteria: “The GALF Hypothesis” in the metagenomic era. Steroids 2017, 125, 1–13. [Google Scholar] [CrossRef]
- Konjević, D.; Janicki, Z.; Slavica, A.; Severin, K.; Krapinec, K.; Božić, F.; Palme, R. Non-invasive monitoring of adrenocortical activity in free-ranging fallow deer (Dama dama L.). Eur. J. Wildl. Res. 2011, 57, 77–81. [Google Scholar] [CrossRef]
- Sheriff, M.J.; Bosson, C.O.; Krebs, C.J.; Boonstra, R. A non-invasive technique for analyzing fecal cortisol metabolites in snowshoe hares (Lepus americanus). J. Comp. Physiol. B 2009, 179, 305–313. [Google Scholar] [CrossRef]
- Palme, R.; Fischer, P.; Schildorfer, H.; Ismail, M. Excretion of infused 14C-steroid hormones via faeces and urine in domestic livestock. Anim. Reprod. Sci. 1996, 43, 43–63. [Google Scholar] [CrossRef]
- Chapman, T.; Kraan, G.; Drayer, N.; Nagel, G.; Wolthers, B.; Colenbrander, B.; Vlissingen, M.F.V. Are (13C) cortisol and (3H) cortisol metabolized identically to natural cortisol in adrenalectomized piglets? Biomed. Environ. Mass Spectrom. 1988, 17, 343–353. [Google Scholar] [CrossRef]
- Papich, M.G. Saunders Handbook of Veterinary Drugs-e-Book: Small and Large Animal; Elsevier Health Sciences: St. Louis, MO, USA, 2015. [Google Scholar]
- Murtagh, R.; Behringer, V.; Deschner, T. LC-MS as a method for non-invasive measurement of steroid hormones and their metabolites in urine and faeces of animals. Wien. Tierarztl. Mon. 2013, 100, 247–254. [Google Scholar]
- Weltring, A.; Schaebs, F.S.; Perry, S.E.; Deschner, T. Simultaneous measurement of endogenous steroid hormones and their metabolites with LC–MS/MS in faeces of a New World primate species, Cebus capucinus. Physiol. Behav. 2012, 105, 510–521. [Google Scholar] [CrossRef]
- Möstl, E.; Rettenbacher, S.; Palme, R. Measurement of Corticosterone Metabolites in Birds’ Droppings: An Analytical Approach. Ann. N. Y. Acad. Sci. 2005, 1046, 17–34. [Google Scholar] [CrossRef] [PubMed]
- Möstl, E.; Palme, R. Hormones as indicators of stress. Domest. Anim. Endocrinol. 2002, 23, 67–74. [Google Scholar] [CrossRef]
- Narayan, E.J.; Webster, K.; Nicolson, V.; Mucci, A.; Hero, J.-M. Non-invasive evaluation of physiological stress in an iconic Australian marsupial: The Koala (Phascolarctos cinereus). Gen. Comp. Endocrinol. 2013, 187, 39–47. [Google Scholar] [CrossRef] [PubMed]
- Bosson, C.O.; Palme, R.; Boonstra, R. Assessment of the stress response in Columbian ground squirrels: Laboratory and field validation of an enzyme immunoassay for fecal cortisol metabolites. Physiol. Biochem. Zool. 2009, 82, 291–301. [Google Scholar] [CrossRef] [PubMed]
- Hovland, A.L.; Rød, A.M.S.; Eriksen, M.S.; Palme, R.; Nordgreen, J.; Mason, G.J. Faecal cortisol metabolites as an indicator of adrenocortical activity in farmed silver foxes (Vulpes vulpes). Appl. Anim. Behav. Sci. 2017, 197, 75–80. [Google Scholar] [CrossRef]
- Santamaria, F.; Palme, R.; Schlagloth, R.; Klobetz-Rassam, E.; Henning, J. Seasonal Variations of Faecal Cortisol Metabolites in Koalas in South East Queensland. Animals 2021, 11, 1622. [Google Scholar] [CrossRef] [PubMed]
- Frigerio, D.; Dittami, J.; Möstl, E.; Kotrschal, K. Excreted corticosterone metabolites co-vary with ambient temperature and air pressure in male Greylag geese (Anser anser). Gen. Comp. Endocrinol. 2004, 137, 29–36. [Google Scholar] [CrossRef]
- Palme, R.; Mostl, E. Measurement of cortisol metabolites in faeces of sheep as a parameter of cortisol concentration in blood. Z. Säugetierkunde Int. J. Mamm. Biol. 1997, 62, 192–197. [Google Scholar]
- Touma, C.; Sachser, N.; Möstl, E.; Palme, R. Effect of sex and time of day on metabolism and excretion of corticosterone in urine and feces of mice. Gen. Comp. Endocrinol. 2003, 130, 267–278. [Google Scholar] [CrossRef]
- Quillfeldt, P.; Möstl, E. Resource allocation in Wilson’s storm-petrels Oceanites oceanicus determined by measurement of glucocorticoid excretion. Acta Ethol. 2003, 5, 115–122. [Google Scholar] [CrossRef]
- Creek, D.J.; Jankevics, A.; Breitling, R.; Watson, D.G.; Barrett, M.P.; Burgess, K.E. Toward global metabolomics analysis with hydrophilic interaction liquid chromatography–mass spectrometry: Improved metabolite identification by retention time prediction. Anal. Chem. 2011, 83, 8703–8710. [Google Scholar] [CrossRef] [PubMed]
- Stoessel, D.; Nowell, C.J.; Jones, A.J.; Ferrins, L.; Ellis, K.M.; Riley, J.; Rahmani, R.; Read, K.D.; McConville, M.J.; Avery, V.M. Metabolomics and lipidomics reveal perturbation of sphingolipid metabolism by a novel anti-trypanosomal 3-(oxazolo [4, 5-b] pyridine-2-yl) anilide. Metabolomics 2016, 12, 126. [Google Scholar] [CrossRef]
- Creek, D.J.; Jankevics, A.; Burgess, K.E.; Breitling, R.; Barrett, M.P. IDEOM: An Excel interface for analysis of LC–MS-based metabolomics data. Bioinformatics 2012, 28, 1048–1049. [Google Scholar] [CrossRef] [PubMed]
- Smith, C.A.; Want, E.J.; O’Maille, G.; Abagyan, R.; Siuzdak, G. XCMS: Processing mass spectrometry data for metabolite profiling using nonlinear peak alignment, matching, and identification. Anal. Chem. 2006, 78, 779–787. [Google Scholar] [CrossRef]
- RC Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2013. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Feature | Mass (Da) | RT (min) | Polarity | Koalas | Comment |
|---|---|---|---|---|---|
| 1 | 462.22579 | 7.12 | Pos | Harvey | Consistent with hydrocortisone succinate (mass = 462.22537, MF = C25H34O8) |
| 462.22484 | 7.25 | Pos | Pixie | ||
| 2 | 366.24031 | 6.28 | Neg | Harvey | Consistent with tetrahydrocortisol or isomers (mass = 366.24063, MF = C21H34O5) |
| 366.23961 | 5.38 | Neg | Pixie | ||
| 3 | 382.23521 | 7.35 | Neg | Harvey | Related to feature 2 by addition of an oxygen (mass = 382.23554, MF = C21H34O6) |
| 382.23475 | 7.43 | Neg | Pixie | ||
| 4 | 384.25102 | 7.46 | Neg | Harvey | Related to feature 2 by addition of water (mass = 384.25119), MF = C21H36O6) |
| 384.25048 | 7.53 | Neg | Pixie | ||
| 5 | 352.22484 | 7.46 | Neg | Harvey | Related to feature 2 by the loss of CH2 (mass = 352.22497, MF = C20H32O5) |
| 352.22445 | 7.47 | Neg | Pixie | ||
| 6 | 483.28366 | 4.35 | Pos | Harvey | Consistent with the ammonium adduct of tetrahydrocortisol succinate (or isomer) (mass = 483.28322, MF = C25H41NO8) |
| 483.28373 | 4.26 | Pos | Pixie |
| Abbreviation | Standard | Trivial Name |
|---|---|---|
| THF | 5β-pregnan-3α,11β,17α,21-tetrol-20-one | tetrahydrocortisol |
| allo-THF | 5α-pregnan-3α,11β,17α, 21-tetrol-20-one | allotetrahydrocortisol |
| 3β-THF | 5β-pregnan-3β,11β,17α,21-tetrol-20-one | 3β-tetrahydrocortisol |
| 3β-allo-THF | 5α-pregnan-3β,11β,17α,21-tetrol-20-one | 3β-allotetrahydrocortisol |
| α-cortolone | 5β-pregnan-3α,17α,20α,21-tetrol-11-one | α-cortolone |
| β-cortolone | 5β-pregnan-3α,17α,20β,21-tetrol-11-one | β-cortolone |
| Compound | Cortisol | 50c | 37e EIAs |
|---|---|---|---|
| THF | <0.001 | 20.7 | <0.001 |
| Allo-THF | 1.4 | <0.01 | 0.5 |
| 3β-THF | 0.1 | <0.001 | 0.2 |
| 3β-allo-THF | 25.0 | <0.001 | 100.0 |
| α-cortolone | <0.0001 | <0.01 | 0.1 |
| β-cortolone | <0.0001 | <0.01 | <0.001 |
| EIA Code | Details | Description |
|---|---|---|
| 69a | Standard | 5β-androstane-3α,11β-diol-17-one (11β-hydroxyaetiocholanolone) |
| Targeted structure | 5β-3α,11β-diol | |
| Antibody against | 5β-androstane-3α,11β-diol-17-one-CMO:BSA | |
| Label | 5β-androstane-3α,11β-diol-17-one-CMO-biotinyl-LC | |
| Reference | Frigerio et al. [36] | |
| 72T | Standard | 5β-androstane-3α-ol-11,17-dione (11-oxoaetiocholanolone) |
| Targeted structure | 5β-3α-ol-11-one | |
| Antibody against | 5β-androstane-3α-ol-11,17-dione-CMO:BSA | |
| Label | 5β-androstane-3α-ol-11,17-dione-CMO-biotinyl-3,6,9-trioxaundecanediamin | |
| Reference | Möstl et al. [13] | |
| Cortisol | Standard | 4-pregnene-11β,17α,21-triol-3,20-dione(cortisol) |
| Targeted structure | 11β,17α,21-triol-20-one | |
| Antibody against | cortisol-3-CMO:BSA | |
| Label | cortisol-3-CMO-DADOO-biotin | |
| Reference | Palme and Möstl [37] | |
| 37e | Standard | 5α-pregnane-3β,11β,21-triol-20-one(3β-allotetrahydrocorticosterone) |
| Targeted structure | 5α-3ß,11ß-diol | |
| Antibody against | 5α-pregnane-3β,11β,21-triol-20-one-CMO | |
| Label | 5α-pregnane-3β,11β,21-triol-20-one-CMO-biotinyl-LC | |
| Reference | Touma et al. [38] | |
| 50c | Standard | 5β-pregnane-3α,11β,21-triol-20-one(tetrahydrocorticosterone) |
| Targeted structure | 5β-3α,11β-diol | |
| Antibody against | 5β-pregnane-3α,11β,21-triol-20-one-CMO | |
| Label | 5β-pregnane-3α,11β,21-triol-20-one-21-HS-biotinyl-LC | |
| Reference | Quillfeldt and Möstl [39] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Santamaria, F.; Barlow, C.K.; Schlagloth, R.; Schittenhelm, R.B.; Palme, R.; Henning, J. Identification of Koala (Phascolarctos cinereus) Faecal Cortisol Metabolites Using Liquid Chromatography-Mass Spectrometry and Enzyme Immunoassays. Metabolites 2021, 11, 393. https://doi.org/10.3390/metabo11060393
Santamaria F, Barlow CK, Schlagloth R, Schittenhelm RB, Palme R, Henning J. Identification of Koala (Phascolarctos cinereus) Faecal Cortisol Metabolites Using Liquid Chromatography-Mass Spectrometry and Enzyme Immunoassays. Metabolites. 2021; 11(6):393. https://doi.org/10.3390/metabo11060393
Chicago/Turabian StyleSantamaria, Flavia, Christopher K. Barlow, Rolf Schlagloth, Ralf B. Schittenhelm, Rupert Palme, and Joerg Henning. 2021. "Identification of Koala (Phascolarctos cinereus) Faecal Cortisol Metabolites Using Liquid Chromatography-Mass Spectrometry and Enzyme Immunoassays" Metabolites 11, no. 6: 393. https://doi.org/10.3390/metabo11060393
APA StyleSantamaria, F., Barlow, C. K., Schlagloth, R., Schittenhelm, R. B., Palme, R., & Henning, J. (2021). Identification of Koala (Phascolarctos cinereus) Faecal Cortisol Metabolites Using Liquid Chromatography-Mass Spectrometry and Enzyme Immunoassays. Metabolites, 11(6), 393. https://doi.org/10.3390/metabo11060393