
Stable Isotope Abundance and Fractionation in Human Diseases


 , ,
, ,
Abstract
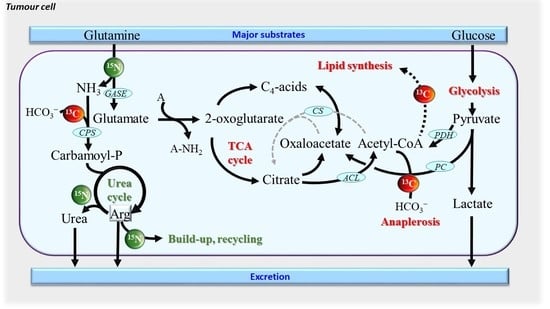
1. Introduction
2. Basics of Stable Isotopes and Metabolic Isotope Effects
2.1. General Principles
2.2. Isotopes in Metabolism Preclinical Studies
2.3. Isotopes in Human Metabolic Syndrome, Diabetes, or Nutritional Stress
2.4. Metabolic Diseases and Isotope Composition of Respired CO2
3. Isotope Fractionation in Cancer
3.1. Cancer Cell Metabolism: Why Might Isotopes Be Impacted?
3.2. Breast Cancer
3.3. Oral Squamous Cell Carcinomas
3.4. Cancer in Infants
3.5. Adrenal Gland Cancer
3.6. Hepatocarcinoma
4. Isotope Fractionation in Metal Homeostasis
4.1. Copper Isotopes in Wilson and Menkes Diseases
4.2. Multiple Metal Isotopes in Other Pathologies
5. Isotopes in Skeletal Pathologies
5.1. Oxygen Isotopes in Sickle-Cell Anaemia
5.2. Nitrogen and Carbon Isotopes in Collagen
6. Conclusions and Perspectives
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A
- δ-value or isotope composition (usually expressed in ‰): abundance of the heavy isotope with respect to that of the light isotope, as compared to the international standard material. It is denoted as δ13C for carbon, δ15N for nitrogen, etc.
- Fractionation (Δ): isotopic difference (usually expressed in ‰) between the substrate and the product of a reaction. It is given by the deviation of the isotope effect (α) from 1, as Δ = α − 1. It can be shown that it can be calculated from δ-values in substrate and product as: Δ = (δsubstrate − δproduct)/(δproduct + 1).
- Isotope effect (α): ratio of rate constants (klight/kheavy) or equilibrium constants (Klight/Kheavy) of the isotopologues of interest. For enzymatic reaction, it is the ratio of catalytic efficiency V/K: α = (V/K)light/(V/K)heavy.
- Isotopologue: isotopic analogue of a molecule, where one atom has been replaced by its isotope. For example, 13C16O2 is the 13C-isotopologue of 12C16O2. It must not be confused with “isotopomers”, which refer to isotopic isomers (for example, 13CH3–12CH2OH and 12CH3–13CH2OH are two isotopomers of ethanol).
- Isotope ratio mass spectrometer (IRMS): mass spectrometer based on a magnetic sector with (usually) fixed collectors (Faraday cups) adapted to quantify precisely the abundance of isotopic species of CO2 (13C analysis), N2 (15N analysis), CO (18O analysis), H2 (2H analysis) or SO2 (34S analysis). The historical origin and technical principles of IRMS are reviewed in [66].
- Quantitative reaction: chemical reaction that consumes all of the substrate molecules, thereby preventing any isotope fractionation. In non-quantitative reactions, substrate molecules left behind may have a δ-value different from the initial value because the reaction selects for an isotopic species (isotope effect).
- Analysis of raw material (total organic matter): typically, raw samples are analysed by elemental analysis (EA) coupled to isotope ratio mass spectrometry (IRMS). In the EA, samples are combusted to CO2 and N2 (13C and 15N analysis) or pyrolised to CO and H2 (18O and 2H analysis). The IRMS measures the mass ratio (45/44 for CO2, 29/28 for N2, etc.) that are then converted to isotope ratios (13C/12C, 14N/15N, etc.). IRMS measurements requires comparison with a reference gas of known δ value. Alternatively, the δ value of the reference gas can be determined by comparison of a certified standard sample of known δ value from the international agency for atomic energy (IAEA, Vienna, Austria). EA-IRMS analyses are adapted to raw samples (lyophilised biopsies or exeresis samples), or pre-purified tissue components (e.g., precipitated proteins, extracted lipids).
- Compound-specific analyses: the isotope composition of specific, targeted metabolites can be determined via three methods: (i) pre-purification with preparative chromatography followed by EA-IRMS; (ii) liquid chromatography coupled to IRMS via a chemical oxidation interface (LC-co-IRMS); and (iii) gas chromatography coupled to IRMS via a combustion interface (GC-c-IRMS). Method (i) has been used extensively in the 90s in plant biology when LC-co-IRMS and GC-c-IRMS were not available. However, this method requires large amounts of material incompatible with small medical samples. Method (ii) is currently limited to carbon isotopes and metabolites that can be resolved using water as an eluent in the LC system. Method (iii) is widely used, and there is now an enormous associated literature, reviewed in (Tea and Tcherkez [67]).
- Gas analyses: gas analysis mostly concerns CO2 produced by respiration and collected in exhaled breath air. There are presently two main techniques to determine the δ13C value in respired CO2: (i) laser-based techniques, and (ii) IRMS-based techniques. Laser-based techniques take advantage of the difference in absorptance between 12CO2 and 13CO2 to compute the 13C/12C ratio of a gas sample. This is a rather simple and instantaneous method that is now implemented routinely for the detection of stomachal ulcers to monitor 13CO2 production from 13C-urea. To perform precise measurements at natural abundance, however, laser-based systems require time-consuming calibration curves not only for CO2 mole fraction but also for δ13C values. This implies the need of gas cylinders at different CO2 concentration and δ values, and thus IRMS measurements for cross-validation. IRMS-based techniques simply use a GC-IRMS coupling whereby air constituents are separated by GC and the CO2 peak is injected into the IRMS. Analyses are slower than with laser systems but provide a precise value with the same requirements as other IRMS measurements.
- Heavy atom isotope analyses: since most IRMS systems analyse simple gases (CO2, N2, H2, SO2, CO), they cannot convey metal analyses. To do so, multi-collectors mass spectrometers are required. In such systems, the sample of interest is broken down by inductively coupled plasma (ICP) and isotope analysis is performed at the atomic level (instead of gases). The clear advantage of this type of instrument is its versatility, because many elements can be analysed just by changing source parameters, not only metals (Mg, Fe, Cu, etc.) but also macroelements (such as S).
- Other techniques: there is now a considerable interest in techniques that can provide information at the intramolecular level, and not solely a molecular average δ value. In fact, metabolic pathways are so that metabolites are fragmented and assembled and therefore strong differences in δ value are anticipated between atom positions within a metabolite. In plant biology, this topic is currently an intense area of research. However, current methods use nuclear magnetic resonance (NMR) which requires quite large amounts of material incompatible with medical samples. Alternative techniques such as Orbitrap®-based analyses are currently under consideration but not applicable to complex mixtures or small samples.
References
- Huidekoper, H.H.; Wijburg, F.A.; Wanders, R. Inborn Errors of Metabolism. In Mass Spectrometry and Stable Isotopes in Nutritional and Pediatric Research; Schierbeek, H., Ed.; John Wiley & Sons: New York, NY, USA, 2017; pp. 258–283. [Google Scholar]
- Bodamer, O.A.F.; Halliday, D. Uses of stable isotopes in clinical diagnosis and research in the paediatric population. Arch. Dis. Child. 2001, 84, 444. [Google Scholar] [CrossRef][Green Version]
- Charidemou, E.; Ashmore, T.; Griffin, J.L. The use of stable isotopes in the study of human pathophysiology. Int. J. Biochem. Cell Biol. 2017, 93, 102–109. [Google Scholar] [CrossRef]
- Turki, A.; Murthy, G.; Ueda, K.; Cheng, B.; Giezen, A.; Stockler-Ipsiroglu, S.; Elango, R. Minimally invasive 13C-breath test to examine phenylalanine metabolism in children with phenylketonuria. Mol. Genet. Metab. 2015, 115, 78–83. [Google Scholar] [CrossRef]
- Jones, P.J.H.; Leatherdale, S.T. Stable isotopes in clinical research: Safety reaffirmed. Clin. Sci. 1991, 80, 277–280. [Google Scholar] [CrossRef] [PubMed]
- Davies, P.S.W. Stable isotopes: Their use and safety in human nutrition studies. Eur. J. Clin. Nutr. 2020, 74, 362–365. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Chen, M.; He, P.; Yu, H.; Block, K.A.; Xie, Z. Composition and spatial distribution of elements and isotopes of a giant human bladder stone and environmental implications. Sci. Total Environ. 2019, 650, 835–846. [Google Scholar] [CrossRef] [PubMed]
- Endo, T.; Hayasaka, M.; Ogasawra, H.; Kimura, O.; Kotaki, Y.; Haraguchi, K. Relationships among mercury concentration, and stable isotope ratios of carbon and nitrogen in the scalp hair of residents from seven countries: Effects of marine fish and C4 plants consumption. PLoS ONE 2015, 10, e0128149. [Google Scholar] [CrossRef] [PubMed]
- Kraft, R.A.; Jahren, A.H.; Saudek, C.D. Clinical-scale investigation of stable isotopes in human blood: δ13C and δ15N from 406 patients at the Johns Hopkins Medical Institutions. Rapid Commun. Mass Spectrom. 2008, 22, 3683–3692. [Google Scholar] [CrossRef] [PubMed]
- Mekota, A.M.; Grupe, G.; Ufer, S.; Cuntz, U. Serial analysis of stable nitrogen and carbon isotopes in hair: Monitoring starvation and recovery phases of patients suffering from anorexia nervosa. Rapid Commun. Mass Spectrom. 2006, 20, 1604–1610. [Google Scholar] [CrossRef]
- Fuller, B.T.; Fuller, J.L.; Sage, N.E.; Harris, D.A.; O’Connell, T.C.; Hedges, R.E.M. Nitrogen balance and δ15N: Why you’re not what you eat during nutritional stress. Rapid Commun. Mass Spectrom. 2005, 19, 2497–2506. [Google Scholar] [CrossRef]
- Salesse, K.; Kaupová, S.; Brůžek, J.; Kuželka, V.; Velemínský, P. An isotopic case study of individuals with syphilis from the pathological-anatomical reference collection of the national museum in Prague (Czech Republic, 19th century A.D.). Int. J. Paleopathol. 2019, 25, 46–55. [Google Scholar] [CrossRef]
- Dailey-Chwalibóg, T.; Huneau, J.F.; Mathé, V.; Kolsteren, P.; Mariotti, F.; Mostak, M.R.; Alim, M.A.; Khan, M.M.S.T.; Khan, M.A.H.; Guesdon, B.; et al. Weaning and stunting affect nitrogen and carbon stable isotope natural abundances in the hair of young children. Sci. Rep. 2020, 10, 2522. [Google Scholar] [CrossRef]
- Park, J.-K.; Ahn, S.V.; Kim, M.K.; Lee, K.-S.; Koh, S.-B.; Bong, Y.-S. The association between carbon and nitrogen stable isotope ratios of human hair and metabolic syndrome. Clin. Chim. Acta 2015, 450, 72–77. [Google Scholar] [CrossRef]
- Hotta, Y.; Fujino, R.; Kimura, O.; Fujii, Y.; Haraguchi, K.; Endo, T. Assessment of diabetics by the quantification of essential elements and stable isotope ratios of carbon and nitrogen in scalp hair. Obes. Med. 2019, 15, 100106. [Google Scholar] [CrossRef]
- Ahn, S.V.; Koh, S.-B.; Lee, K.-S.; Bong, Y.-S.; Park, J.-K. Association between nitrogen stable isotope ratios in human hair and serum levels of leptin. Tohoku J. Exp. Med. 2017, 243, 133–139. [Google Scholar] [CrossRef] [PubMed]
- Henze, M.M.; Bemis, E.A.; Seifert, J.A.; Johnson, R.K.; Dong, F.; Rewers, M.; Norris, J.M. Association between change in self-reported sugar intake and a sugar biomarker (δ13C) in children at increased risk for type 1 diabetes. J. Nutr. Sci. 2020, 9, e16. [Google Scholar] [CrossRef]
- Ghosh, C.; Mandal, S.; Pal, M.; Mukhopadhyay, P.; Ghosh, S.; Pradhan, M. 13C isotopic abundances in natural nutrients: A newly formulated test meal for non-invasive diagnosis of type 2 diabetes. J. Breath Res. 2017, 11, 026005. [Google Scholar] [CrossRef] [PubMed]
- Petzke, K.J.; Feist, T.; Fleig, W.E.; Metges, C.C. Nitrogen isotopic composition in hair protein is different in liver cirrhotic patients. Rapid Commun Mass Spectrom 2006, 20, 2973–2978. [Google Scholar] [CrossRef] [PubMed]
- Tea, I.; Martineau, E.; Antheaume, I.; Lalande, J.; Mauve, C.; Gilard, F.; Barillé-Nion, S.; Blackburn, A.C.; Tcherkez, G. 13C and 15N natural isotope abundance reflects breast cancer cell metabolism. Sci. Rep. 2016, 6. [Google Scholar] [CrossRef] [PubMed]
- Tea, I.M.E.; Giraudeau, P.; Akoka, S.; Barillé-Nion, S. Method to Characterize the Origin and/or the State of Pathological or Healthy cells, and Its Applications in Biology. E.P. Patent No. 2686686B1, 14 October 2015. Available online: https://patents.google.com/patent/EP2686686B1/en (accessed on 7 May 2021).
- Bogusiak, K.; Puch, A.; Mostowski, R.; Kozakiewicz, M.; Paneth, P.; Kobos, J. Characteristic of Oral Squamous Cell Carcinoma Tissues Using Isotope Ratio Mass Spectrometry. J. Clin. Med. 2020, 9, 3760. [Google Scholar] [CrossRef] [PubMed]
- Taran, K.; Frączek, T.; Kamiński, R.; Sitkiewicz, A.; Kobos, J.; Paneth, P. The first protocol of stable isotope ratio assessment in tumor tissues based on original research. Pol. J. Pathol. 2015, 66, 288–295. [Google Scholar] [CrossRef]
- Taran, K.; Frączek, T.; Sikora-Szubert, A.; Sitkiewicz, A.; Młynarski, W.; Kobos, J.; Paneth, P. The first investigation of Wilms’ tumour atomic structure-nitrogen and carbon isotopic composition as a novel biomarker for the most individual approach in cancer disease. Oncotarget 2016, 7, 76726. [Google Scholar] [CrossRef][Green Version]
- Taran, K.; Frączek, T.; Sitkiewicz, A.; Paneth, P.; Kobos, J. Rhabdomyosarcoma in children in the light of isotope ratio mass spectrometry. Pol. J. Pathol. 2015, 66, 383–388. [Google Scholar] [CrossRef]
- Balter, V.; Nogueira da Costa, A.; Bondanese, V.P.; Jaouen, K.; Lamboux, A.; Sangrajrang, S.; Vincent, N.; Fourel, F.; Télouk, P.; Gigou, M.; et al. Natural variations of copper and sulfur stable isotopes in blood of hepatocellular carcinoma patients. Proc. Natl. Acad. Sci. USA 2015, 112, 982–985. [Google Scholar] [CrossRef]
- Tanaka, Y.K.; Hirata, T. Stable Isotope Composition of Metal Elements in Biological Samples as Tracers for Element Metabolism. Anal. Sci. Int. J. Jpn. Soc. Anal. Chem. 2018, 34, 645–655. [Google Scholar] [CrossRef] [PubMed]
- Toubhans, B.; Gourlan, A.T.; Telouk, P.; Lutchman-Singh, K.; Francis, L.W.; Conlan, R.S.; Margarit, L.; Gonzalez, D.; Charlet, L. Cu isotope ratios are meaningful in ovarian cancer diagnosis. J. Trace Elem. Med. Biol. 2020, 62, 126611. [Google Scholar] [CrossRef] [PubMed]
- Hastuti, A.; Costas-Rodríguez, M.; Anoshkina, Y.; Parnall, T.; Madura, J.A., 2nd; Vanhaecke, F. High-precision isotopic analysis of serum and whole blood Cu, Fe and Zn to assess possible homeostasis alterations due to bariatric surgery. Anal. Bioanal. Chem. 2020, 412, 727–738. [Google Scholar] [CrossRef] [PubMed]
- Hastuti, A.A.M.B.; Costas-Rodríguez, M.; Matsunaga, A.; Ichinose, T.; Hagiwara, S.; Shimura, M.; Vanhaecke, F. Cu and Zn isotope ratio variations in plasma for survival prediction in hematological malignancy cases. Sci. Rep. 2020, 10, 16389. [Google Scholar] [CrossRef]
- Gordon, G.W.; Monge, J.; Channon, M.B.; Wu, Q.; Skulan, J.L.; Anbar, A.D.; Fonseca, R. Predicting multiple myeloma disease activity by analyzing natural calcium isotopic composition. Leukemia 2014, 28, 2112–2115. [Google Scholar] [CrossRef]
- Tanaka, Y.-k.; Yajima, N.; Higuchi, Y.; Yamato, H.; Hirata, T. Calcium isotope signature: New proxy for net change in bone volume for chronic kidney disease and diabetic rats. Metallomics 2017, 9, 1745–1755. [Google Scholar] [CrossRef]
- Carroll, G.M.A.; Inskip, S.; Waters-Rist, A. Pathophysiological Stable Isotope Fractionation: Assessing the Impact of Anemia on Enamel Apatite d18O and d13C Values and Bone Collagen d15N and d13C Values. Bioarchaeology Int. 2018, 2, 117–146. [Google Scholar] [CrossRef]
- Reitsema, L.J. Beyond diet reconstruction: Stable isotope applications to human physiology, health, and nutrition. Am. J. Hum. Biol. 2013, 25, 445–456. [Google Scholar] [CrossRef]
- Scorrano, G.; Brilli, M.; Martínez-Labarga, C.; Giustini, F.; Pacciani, E.; Chilleri, F.; Scaldaferri, F.; Gasbarrini, A.; Gasbarrini, G.; Rickards, O. Palaeodiet reconstruction in a woman with probable celiac disease: A stable isotope analysis of bone remains from the archaeological site of Cosa (Italy). Am. J. Phys. Anthropol. 2014, 154, 349–356. [Google Scholar] [CrossRef] [PubMed]
- Bernardo, K.; Jousse, C.; Fafournoux, P.; Schiphorst, A.M.; Grand, M.; Robins, R.J.; Hankard, R.; De Luca, A. Protein restricted diet during gestation and/or lactation in mice affects 15N natural isotopic abundance of organs in the offspring: Effect of diet 15N content and growth. PLoS ONE 2018, 13, e0205271. [Google Scholar] [CrossRef]
- Mantha, O.L.; Polakof, S.; Huneau, J.-F.; Mariotti, F.; Poupin, N.; Zalko, D.; Fouillet, H. Early changes in tissue amino acid metabolism and nutrient routing in rats fed a high-fat diet: Evidence from natural isotope abundances of nitrogen and carbon in tissue proteins. Br. J. Nutr. 2018, 119, 981–991. [Google Scholar] [CrossRef] [PubMed]
- Huneau, J.-F.; Mantha, O.L.; Hermier, D.; Mathé, V.; Galmiche, G.; Mariotti, F.; Fouillet, H. Natural isotope abundances of carbon and nitrogen in tissue proteins and amino acids as biomarkers of the decreased carbohydrate oxidation and increased amino acid oxidation induced by caloric restriction under a maintained protein intake in obese rats. Nutrients 2019, 11, 1087. [Google Scholar] [CrossRef]
- Petzke, K.J.; Fuller, B.T.; Metges, C.C. Advances in natural stable isotope ratio analysis of human hair to determine nutritional and metabolic status. Curr. Opin. Clin. Nutr. Metab. Care 2010, 13, 532–540. [Google Scholar] [CrossRef]
- Liu, S.V.; Moore, L.B.; Halliday, T.M.; Jahren, A.H.; Savla, J.; Hedrick, V.E.; Marinik, E.L.; Davy, B.M. Short-term changes in added sugar consumption by adolescents reflected in the carbon isotope ratio of fingerstick blood. Nutr. Health 2018, 24, 251–259. [Google Scholar] [CrossRef]
- De Luca, A.; Laugier, S.; Tea, I.; Robins, R.J.; Saulnier, P.-J.; Torremocha, F.; Piguel, X.; Maréchaud, R.; Hankard, R.; Hadjadj, S. Impact on bulk 15N natural isotopic abundance in hair of kidney function in type 2 diabetic nephropathy. e-SPEN J. 2014, 9, e204–e209. [Google Scholar] [CrossRef]
- Sick, H.; Roos, N.; Saggau, E.; Haas, K.; Meyn, V.; Walch, B.; Trugo, N. Amino acid utilization and isotope discrimination of amino nitrogen in nitrogen metabolism of rat liver in vivo. Z. Fur Ernahr. 1997, 36, 340–346. [Google Scholar] [CrossRef]
- Kårlund, A.; Kääriäinen, T.; Kostamo, V.; Kokkola, T.; Kolehmainen, M.; Lakka, T.; Pihlajamäki, J.; Manninen, A. Oxygen-18 and Carbon-13 isotopes in eCO2 and erythrocytes carbonic anhydrase activity of Finnish prediabetic population. J. Breath Res. 2020, in press. [Google Scholar]
- Boriosi, J.P.; Maki, D.G.; Yngsdal-Krenz, R.A.; Wald, E.R.; Porter, W.P.; Cook, M.E.; Bütz, D.E. Changes in breath carbon isotope composition as a potential biomarker of inflammatory acute phase response in mechanically ventilated pediatric patients. J. Anal. At. Spectrom. 2014, 29, 599–605. [Google Scholar] [CrossRef]
- Keshet, R.; Szlosarek, P.; Carracedo, A.; Erez, A. Rewiring urea cycle metabolism in cancer to support anabolism. Nat. Rev. Cancer 2018, 18, 634–645. [Google Scholar] [CrossRef] [PubMed]
- Lloyd, S.M.; Arnold, J.; Sreekumar, A. Metabolomic profiling of hormone-dependent cancers: A bird’s eye view. Trends Endocrinol. Metab. TEM 2015, 26, 477–485. [Google Scholar] [CrossRef] [PubMed]
- Cai, D.; Wang, J.; Gao, B.; Li, J.; Wu, F.; Zou, J.X.; Xu, J.; Jiang, Y.; Zou, H.; Huang, Z.; et al. RORγ is a targetable master regulator of cholesterol biosynthesis in a cancer subtype. Nat. Commun. 2019, 10, 4621. [Google Scholar] [CrossRef]
- Beckwith, J.B. Wilms Tumor and Other Renal Tumors of Childhood: An Update. J. Urol. 1986, 136, 320–324. [Google Scholar] [CrossRef]
- Vujanić, G.M.; Sandstedt, B.; Harms, D.; Kelsey, A.; Leuschner, I.; de Kraker, J. Revised International Society of Paediatric Oncology (SIOP) working classification of renal tumors of childhood. Med. Pediatr. Oncol. 2002, 38, 79–82. [Google Scholar] [CrossRef]
- Neville, H.L.; Ritchey, M.L. Wilms’ tumor. Overview of National Wilms’ Tumor Study Group results. Urol. Clin. N. Am. 2000, 27, 435–442. [Google Scholar] [CrossRef]
- Faria, P.; Beckwith, J.B.; Mishra, K.; Zuppan, C.; Weeks, D.A.; Breslow, N.; Green, D.M. Focal versus diffuse anaplasia in Wilms tumor--new definitions with prognostic significance: A report from the National Wilms Tumor Study Group. Am. J. Surg. Pathol. 1996, 20, 909–920. [Google Scholar] [CrossRef]
- Dome, J.S.; Cotton, C.A.; Perlman, E.J.; Breslow, N.E.; Kalapurakal, J.A.; Ritchey, M.L.; Grundy, P.E.; Malogolowkin, M.; Beckwith, J.B.; Shamberger, R.C.; et al. Treatment of anaplastic histology Wilms’ tumor: Results from the fifth National Wilms’ Tumor Study. J. Clin. Oncol. Off. J. Am. Soc. Clin. Oncol. 2006, 24, 2352–2358. [Google Scholar] [CrossRef]
- Shamberger, R.C.; Anderson, J.R.; Breslow, N.E.; Perlman, E.J.; Beckwith, J.B.; Ritchey, M.L.; Haase, G.M.; Donaldson, M.; Grundy, P.E.; Weetman, R.; et al. Long-term outcomes for infants with very low risk Wilms tumor treated with surgery alone in National Wilms Tumor Study-5. Ann. Surg. 2010, 251, 555–558. [Google Scholar] [CrossRef] [PubMed]
- Pinheiro, C.; Granja, S.; Longatto-Filho, A.; Faria, A.M.; Fragoso, M.C.; Lovisolo, S.M.; Lerário, A.M.; Almeida, M.Q.; Baltazar, F.; Zerbini, M.C. Metabolic reprogramming: A new relevant pathway in adult adrenocortical tumors. Oncotarget 2015, 6, 44403–44421. [Google Scholar] [CrossRef]
- Lo, M.; Wang, Y.Z.; Gout, P.W. The x(c)- cystine/glutamate antiporter: A potential target for therapy of cancer and other diseases. J. Cell. Physiol. 2008, 215, 593–602. [Google Scholar] [CrossRef] [PubMed]
- Philippot, P.; Van Zuilen, M.; Lepot, K.; Thomazo, C.; Farquhar, J.; Van Kranendonk, M.J. Early Archaean Microorganisms Preferred Elemental Sulfur, Not Sulfate. Science 2007, 317, 1534. [Google Scholar] [CrossRef] [PubMed]
- Tcherkez, G.; Tea, I. 32S/34S isotope fractionation in plant sulphur metabolism. New Phytol. 2013, 200. [Google Scholar] [CrossRef]
- Albalat, E.; Telouk, P.; Balter, V.; Fujii, T.; Bondanese, V.P.; Plissonnier, M.-L.; Vlaeminck-Guillem, V.; Baccheta, J.; Thiam, N.; Miossec, P.; et al. Sulfur isotope analysis by MC-ICP-MS and application to small medical samples. J. Anal. At. Spectrom. 2016, 31, 1002–1011. [Google Scholar] [CrossRef]
- Albarède, F.; Télouk, P.; Balter, V. Medical Applications of Isotope Metallomics. Rev. Mineral. Geochem. 2017, 82, 851–885. [Google Scholar] [CrossRef]
- Albarede, F.; Télouk, P.; Balter, V.; Bondanese, V.P.; Albalat, E.; Oger, P.; Bonaventura, P.; Miossec, P.; Fujii, T. Medical applications of Cu, Zn, and S isotope effects. Metallomics 2016, 8, 1056–1070. [Google Scholar] [CrossRef]
- Balter, V.; Lamboux, A.; Zazzo, A.; Télouk, P.; Leverrier, Y.; Marvel, J.; Moloney, A.P.; Monahan, F.J.; Schmidt, O.; Albarède, F. Contrasting Cu, Fe, and Zn isotopic patterns in organs and body fluids of mice and sheep, with emphasis on cellular fractionation. Metallomics 2013, 5, 1470–1482. [Google Scholar] [CrossRef]
- Kaler, S.G. ATP7A-related copper transport diseases—emerging concepts and future trends. Nat. Rev. Neurol. 2011, 7, 15–29. [Google Scholar] [CrossRef]
- Lamboux, A.; Couchonnal-Bedoya, E.; Guillaud, O.; Laurencin, C.; Lion-François, L.; Belmalih, A.; Mintz, E.; Brun, V.; Bost, M.; Lachaux, A.; et al. The blood copper isotopic composition is a prognostic indicator of the hepatic injury in Wilson disease. Metallomics 2020, 12, 1781–1790. [Google Scholar] [CrossRef] [PubMed]
- Reitsema, L.J.; Crews, D.E. Brief communication: Oxygen isotopes as a biomarker for sickle-cell disease? Results from transgenic mice expressing human hemoglobin S genes. Am J Phys Anthr. 2011, 145, 495–498. [Google Scholar] [CrossRef] [PubMed]
- Curto, A.; Mahoney, P.; Maurer, A.-F.; Barrocas-Dias, C.; Fernandes, T.; Fahy, G.E. Effect of different healing stages on stable isotope ratios in skeletal lesions. Am. J. Phys. Anthropol. 2020, 171, 285–297. [Google Scholar] [CrossRef] [PubMed]
- Wilkinson, D.J. Historical and contemporary stable isotope tracer approaches to studying mammalian protein metabolism. Mass Spectrom. Rev. 2018, 37, 57–80. [Google Scholar] [CrossRef] [PubMed]
- Tea, I.; Tcherkez, G. Natural Isotope Abundance in Metabolites: Techniques and Kinetic Isotope Effect Measurement in Plant, Animal, and Human Tissues. Methods Enzymol. 2017. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
| Disease | Metabolic Mechanism | Isotopic Marker | Matrix | Ref. |
|---|---|---|---|---|
| Nervous anorexia, nutritional stress | Aa metabolism | 13C, 15N | Hair | [10,11] |
| Syphilis | Aa metabolism | 13C, 15N | Collagen | [12] |
| Chronic malnutrition and potential growth retardation (stunted children) | Aa metabolism | 13C, 15N | Hair | [13] |
| Patients with metabolic syndrome | Glycaemia Aa metabolism | 13C, 15N | Hair | [14] |
| Diabetic patients | Sugar metabolism | 13C, 15N | Hair | [15,16,17,18] |
| Cirrhotic patients | Aa metabolism | 13C, 15N | Hair, bulk protein | [19] |
| Breast cancer | Urea cycle, glycolysis, lipid synthesis, anaplerosis | 13C, 15N | Tissue biopsies cultured cells | [20,21] |
| Oral squamous cell carcinomas | ND | 13C, 15N | Tissue biopsies | [22] |
| Ganglioneuroma (benign tumours), neuroblastoma and nephroblastoma Wilm’s tumours | Aa metabolism | 13C, 15N | Tissue biopsies | [23,24] |
| Rhabdomyosarcoma | ND | 13C, 15N | Tissue biopsies | [25] |
| Adrenal gland cancers | Aa metabolism Glycolysis | 13C, 15N | Serum | Unpublished data |
| Hepatocarcinoma | Glutathione metabolism, | 34S | Serum and erythrocytes | [26] |
| Wilson disease | Cu metabolism | 65Cu | Serum | [27] |
| Menkes disease | Cu and Aa metabolism | 15N | Hair | Unpublished data |
| Ovarian cancer | Cu metabolism | 65Cu | Serum | [28] |
| Homeostasis alterations after bariatric surgery | Zn homeostasis | 66Zn | Serum and Whole blood | [29] |
| Hematological malignancy | Metal homeostasis | 65Cu, 66Zn | serum | [30] |
| Anaemia | Fe deficiency | 56Fe | Whole blood | [27] |
| Multiple myeloma | Bone formation (apatite deposition) | 44Ca | Serum and urine | [31] |
| Chronic kidney disease or diabetes | Bone formation (apatite deposition) | 44Ca | Serum | [32] |
| Anaemia in skeleton fragments | Respiratory biochemistry | 18O | Bone and enamel apatite | [33] |
| Osteopenia and osteoporosis in female skeleton | Urea excretion and/or renal function | 15N | Bone collagen | [34] |
| Cealiac disease in skeleton | Aa metabolism | 13C, 15N | Bone collagen | [35] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tea, I.; De Luca, A.; Schiphorst, A.-M.; Grand, M.; Barillé-Nion, S.; Mirallié, E.; Drui, D.; Krempf, M.; Hankard, R.; Tcherkez, G. Stable Isotope Abundance and Fractionation in Human Diseases. Metabolites 2021, 11, 370. https://doi.org/10.3390/metabo11060370
Tea I, De Luca A, Schiphorst A-M, Grand M, Barillé-Nion S, Mirallié E, Drui D, Krempf M, Hankard R, Tcherkez G. Stable Isotope Abundance and Fractionation in Human Diseases. Metabolites. 2021; 11(6):370. https://doi.org/10.3390/metabo11060370
Chicago/Turabian StyleTea, Illa, Arnaud De Luca, Anne-Marie Schiphorst, Mathilde Grand, Sophie Barillé-Nion, Eric Mirallié, Delphine Drui, Michel Krempf, Régis Hankard, and Guillaume Tcherkez. 2021. "Stable Isotope Abundance and Fractionation in Human Diseases" Metabolites 11, no. 6: 370. https://doi.org/10.3390/metabo11060370
APA StyleTea, I., De Luca, A., Schiphorst, A.-M., Grand, M., Barillé-Nion, S., Mirallié, E., Drui, D., Krempf, M., Hankard, R., & Tcherkez, G. (2021). Stable Isotope Abundance and Fractionation in Human Diseases. Metabolites, 11(6), 370. https://doi.org/10.3390/metabo11060370