
Mass Spectrometry Imaging as a Tool to Investigate Region Specific Lipid Alterations in Symptomatic Human Carotid Atherosclerotic Plaques

 , ,
, ,
Abstract
1. Introduction
2. Results and Discussion
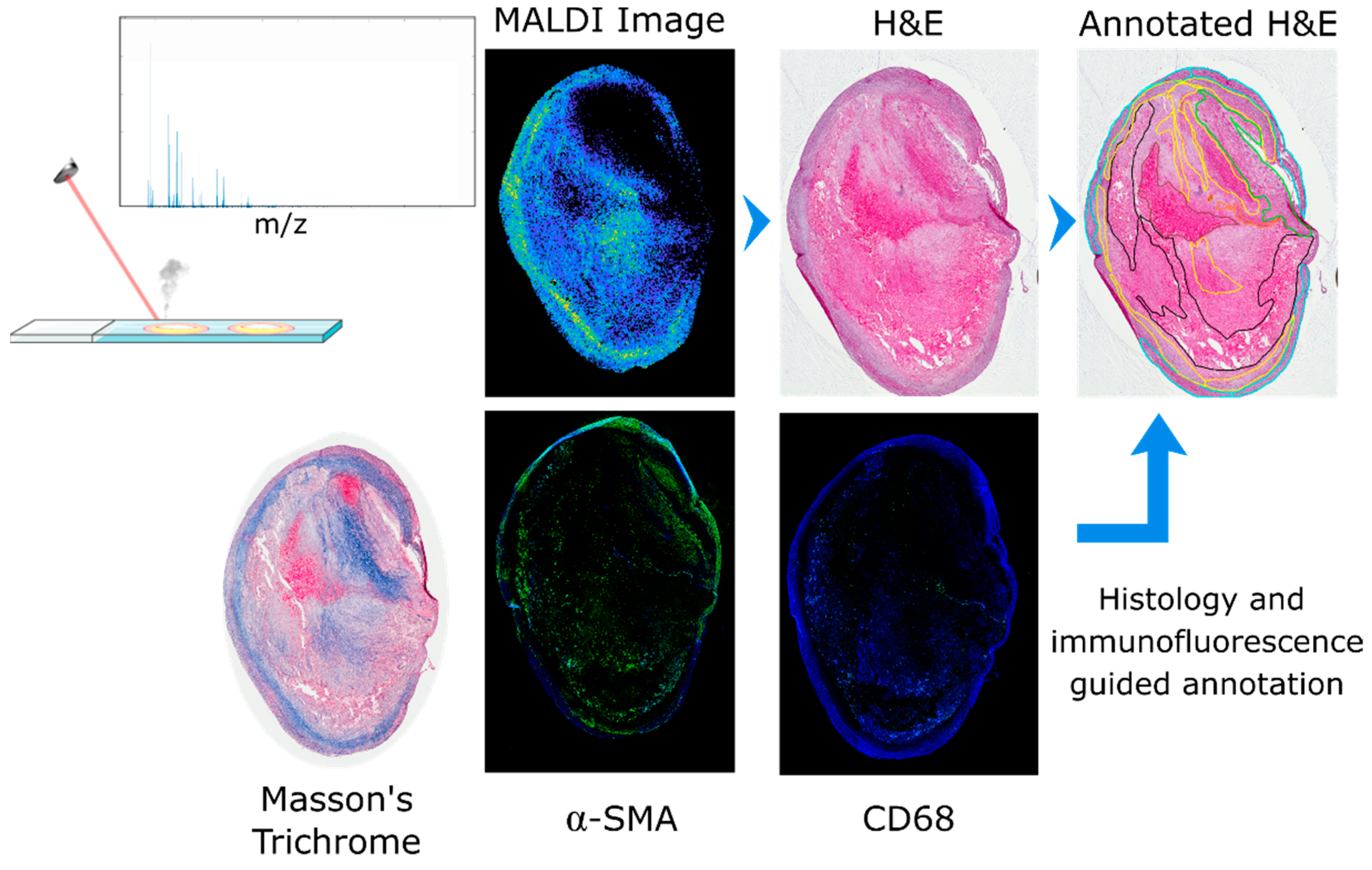
2.1. Image Co-Registration and Region Definition
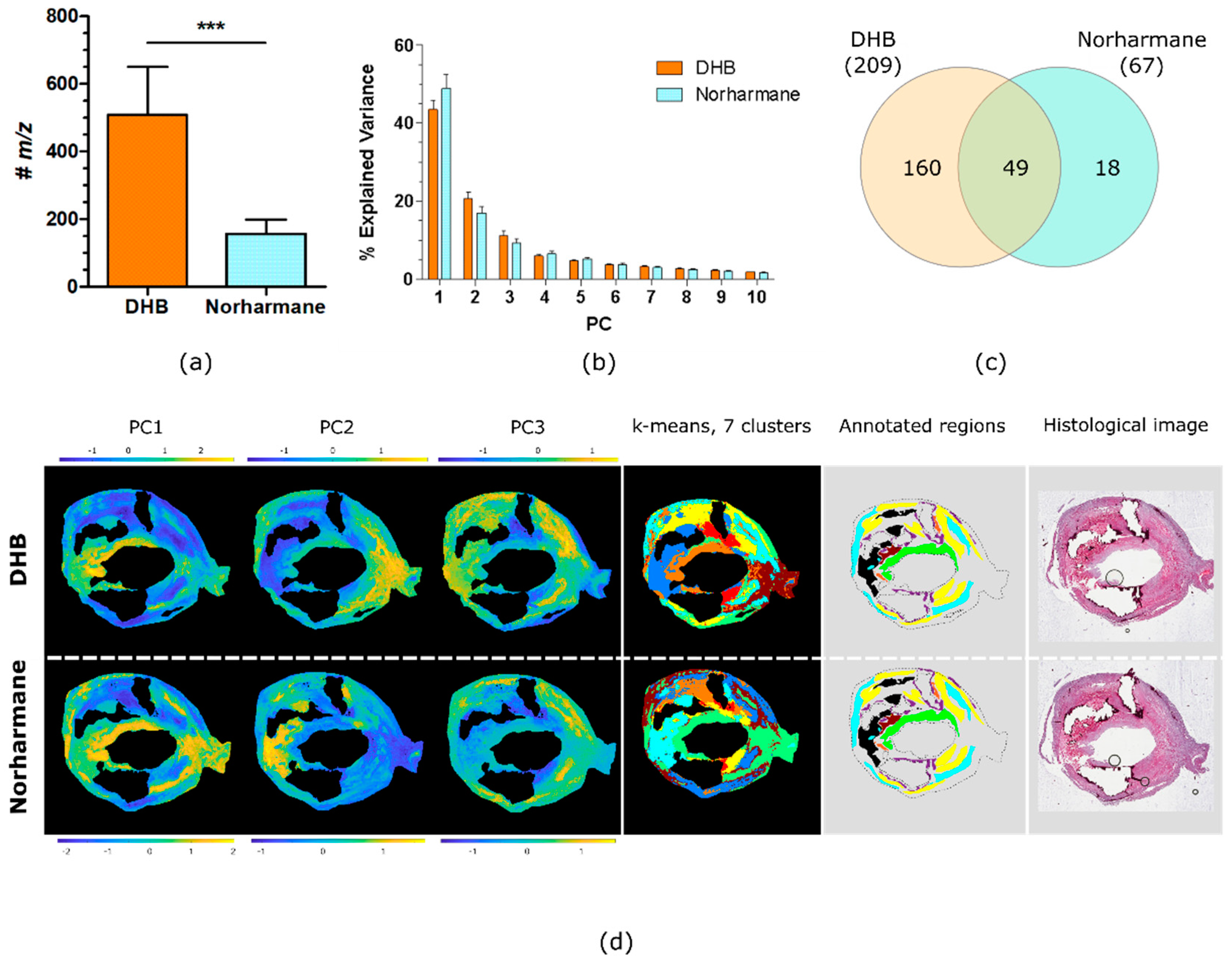
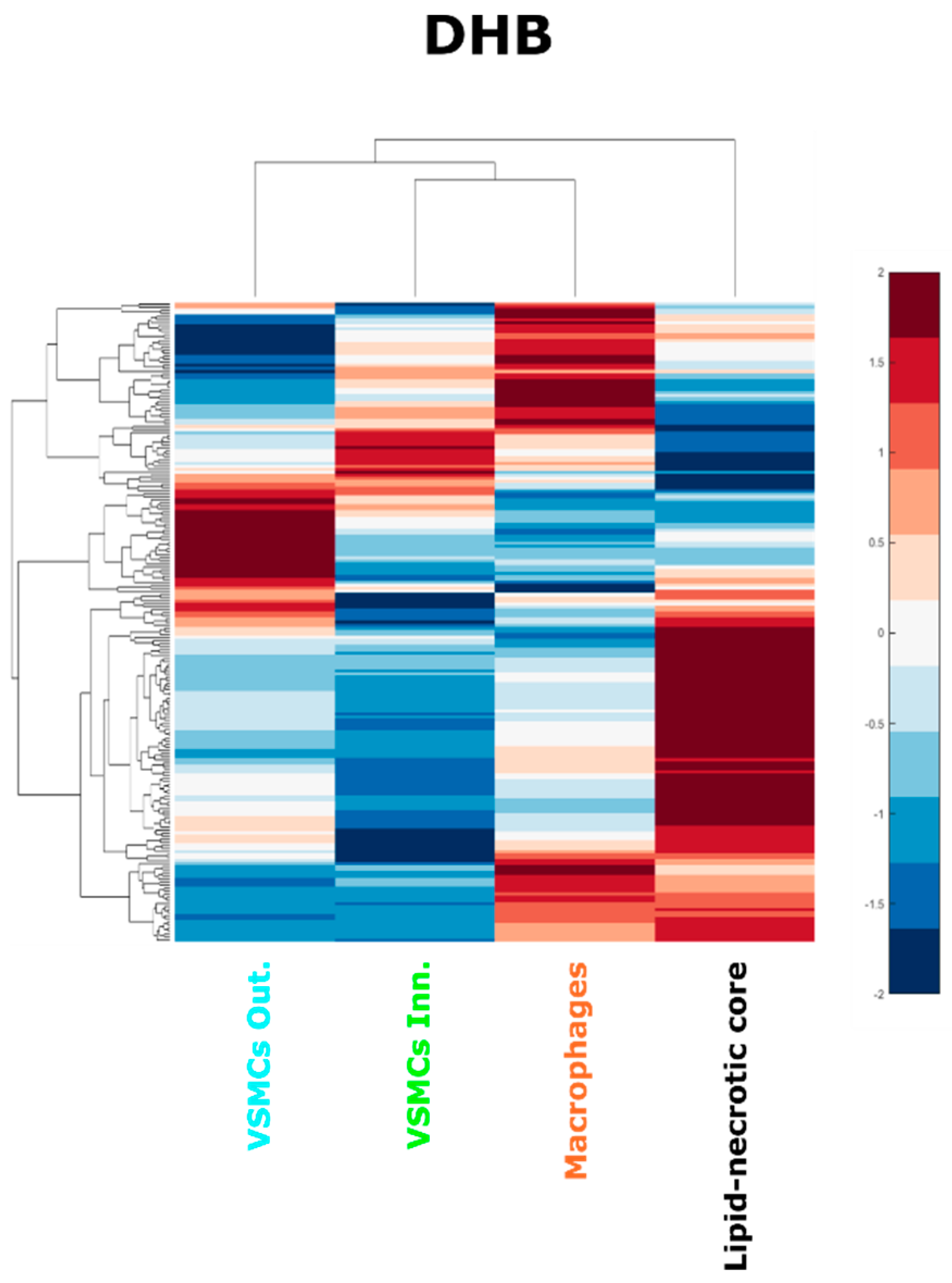
2.2. Comparison of DHB and Norharmane Matrices
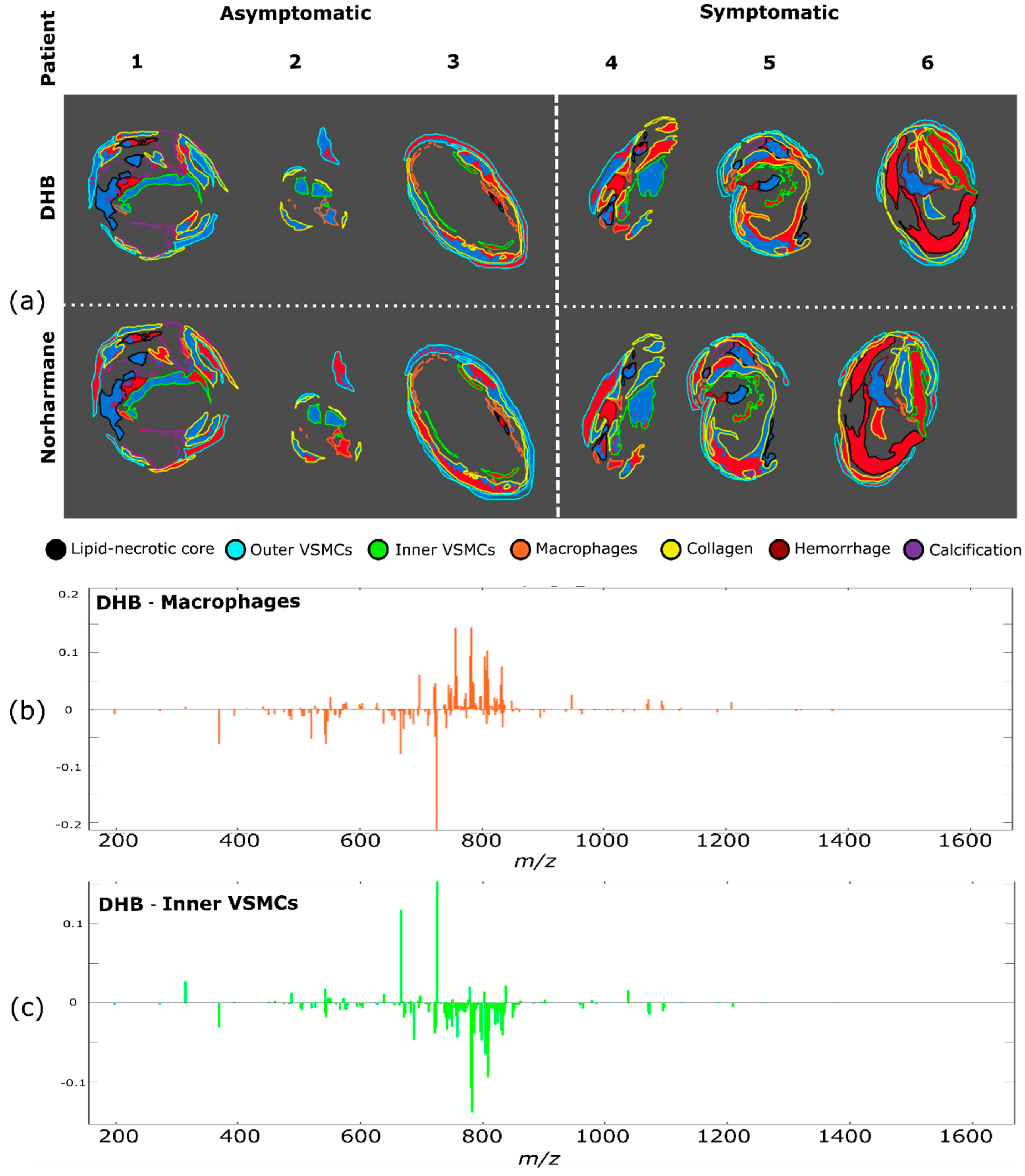
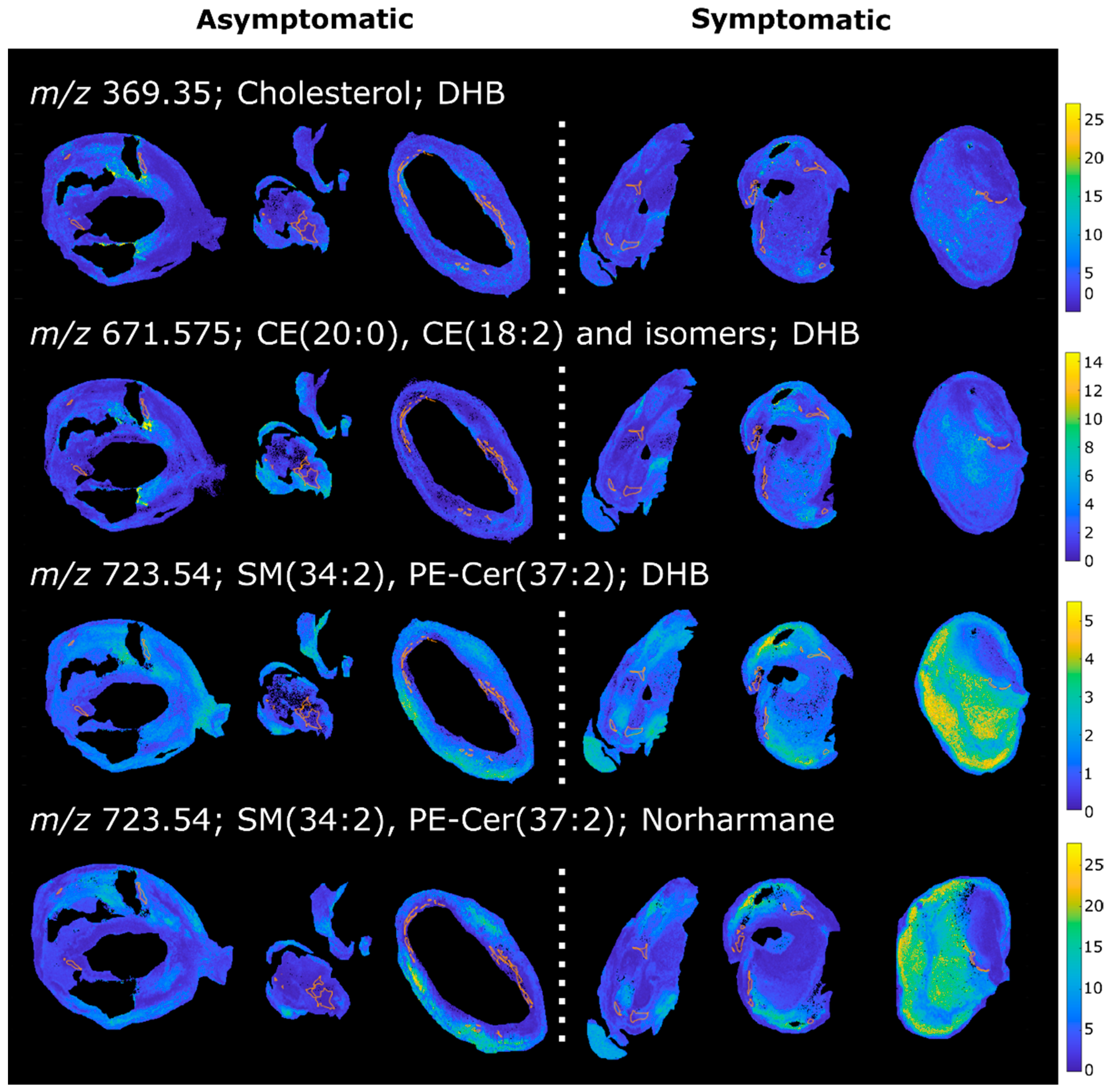
2.3. Patient Series Analyses
2.4. PCA of the Patient Series
2.5. K-Means Cluster Analysis of the Patient Series
2.6. PLS Regression of the Patient Series
2.7. Hierarchical Cluster Analysis
2.8. Study Limitations and Future Perspectives
3. Materials and Methods
3.1. Materials
3.2. Tissue Collection
3.3. Sample Preparation
3.4. MALDI MSI Data Acquisition
3.5. Histological Staining
3.6. Immunofluorescence
3.7. Histological Annotation
3.8. MSI Data Preprocessing
3.9. MSI Data Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kobiyama, K.; Ley, K. Atherosclerosis: A Chronic Inflammatory Disease with an Autoimmune Component. Circ. Res. 2018, 123, 1118–1120. [Google Scholar] [CrossRef]
- Bergheanu, S.C.; Bodde, M.C.; Jukema, J.W. Pathophysiology and treatment of atherosclerosis: Current view and future perspective on lipoprotein modification treatment. Netherlands Hear. J. 2017, 25, 231–242. [Google Scholar] [CrossRef]
- Lusis, A.J. Atherosclerosis. Nature 2010, 407, 233–241. [Google Scholar] [CrossRef]
- Thomas, H.; Diamond, J.; Vieco, A.; Chaudhuri, S.; Shinnar, E.; Cromer, S.; Perel, P.; Mensah, G.A.; Narula, J.; Johnson, C.O.; et al. Global Atlas of Cardiovascular Disease 2000–2016: The Path to Prevention and Control. Glob. Heart 2018, 13, 143–163. [Google Scholar] [CrossRef] [PubMed]
- Hansson, G.; Robertson, A.-K.; Soderberg-Naucler, C. Inflammation and atherosclerosis. Annu. Rev. Pathol. 2006, 1, 297–329. [Google Scholar] [CrossRef]
- Seimon, T.; Tabas, I. Mechanisms and consequences of macrophage apoptosis in atherosclerosis. J. Lipid Res. 2009, 50, S382–S387. [Google Scholar] [CrossRef] [PubMed]
- Chistiakov, D.A.; Orekhov, A.N.; Bobryshev, Y.V. Vascular smooth muscle cell in atherosclerosis. Acta Physiol. 2015, 214, 33–50. [Google Scholar] [CrossRef] [PubMed]
- Basatemur, G.L.; Jørgensen, H.F.; Clarke, M.C.H.; Bennett, M.R.; Mallat, Z. Vascular smooth muscle cells in atherosclerosis. Nat. Rev. Cardiol. 2019, 16, 727–744. [Google Scholar] [CrossRef] [PubMed]
- Halvorsen, B.; Otterdal, K.; Dahl, T.B.; Skjelland, M.; Gullestad, L.; Øie, E.; Aukrust, P. Atherosclerotic Plaque Stability-What Determines the Fate of a Plaque? Prog. Cardiovasc. Dis. 2008, 51, 183–194. [Google Scholar] [CrossRef]
- Nilsson, J. Atherosclerotic plaque vulnerability in the statin era. Eur. Heart J. 2017, 38, 1638–1644. [Google Scholar] [CrossRef]
- Paty, P.S.K.; Adeniyi, J.A.; Mehta, M.; Darling, R.C.; Chang, B.B.; Kreienberg, P.B.; Ozsvath, K.J.; Roddy, S.P.; Shah, D.M. Surgical treatment of internal carotid artery occlusion. J. Vasc. Surg. 2003, 37, 785–788. [Google Scholar] [CrossRef]
- Golledge, J.; Greenhalgh, R.M.; Davies, A.H. The symptomatic carotid plaque. Stroke 2000, 31, 774–781. [Google Scholar] [CrossRef] [PubMed]
- Naylor, A.R.; Ricco, J.B.; de Borst, G.J.; Debus, S.; de Haro, J.; Halliday, A.; Hamilton, G.; Kakisis, J.; Kakkos, S.; Lepidi, S.; et al. Editor’s Choice—Management of Atherosclerotic Carotid and Vertebral Artery Disease: 2017 Clinical Practice Guidelines of the European Society for Vascular Surgery (ESVS). Eur. J. Vasc. Endovasc. Surg. 2018, 55, 3–81. [Google Scholar] [CrossRef] [PubMed]
- Mezger, S.T.P.; Mingels, A.M.A.; Bekers, O.; Cillero-Pastor, B.; Heeren, R.M.A. Trends in mass spectrometry imaging for cardiovascular diseases. Anal. Bioanal. Chem. 2019, 411, 3709–3720. [Google Scholar] [CrossRef] [PubMed]
- Hillenkamp, F. MALDI MS A Practical Guide to Instrumentation, Methods and Applications, 2nd ed.; Blackwell, Wiley: Weinheim, Germany, 2014; ISBN 9780470035245. [Google Scholar]
- Cornett, D.S.; Reyzer, M.L.; Chaurand, P.; Caprioli, R.M. MALDI imaging mass spectrometry: Molecular snapshots of biochemical systems. Nat. Methods 2007, 4, 828–833. [Google Scholar] [CrossRef] [PubMed]
- McDonnell, L.A.; Heeren, R.M.A. Imaging mass spectrometry. Mass Spectrom. Rev. 2007, 26, 606–643. [Google Scholar] [CrossRef] [PubMed]
- Martin-Lorenzo, M.; Alvarez-Llamas, G.; McDonnell, L.A.; Vivanco, F. Molecular histology of arteries: Mass spectrometry imaging as a novel ex vivo tool to investigate Atherosclerosis. Expert Rev. Proteomics 2016, 13, 69–81. [Google Scholar] [CrossRef]
- Pirillo, A.; Bonacina, F.; Norata, G.D.; Catapano, A.L. The Interplay of Lipids, Lipoproteins, and Immunity in Atherosclerosis. Curr. Atheroscler. Rep. 2018, 20. [Google Scholar] [CrossRef]
- Zaima, N.; Sasaki, T.; Tanaka, H.; Cheng, X.W.; Onoue, K.; Hayasaka, T.; Goto-Inoue, N.; Enomoto, H.; Unno, N.; Kuzuya, M.; et al. Imaging mass spectrometry-based histopathologic examination of atherosclerotic lesions. Atherosclerosis 2011, 217, 427–432. [Google Scholar] [CrossRef]
- Patterson, N.H.; Doonan, R.J.; Daskalopoulou, S.S.; Dufresne, M.; Lenglet, S.; Montecucco, F.; Thomas, A.; Chaurand, P. Three-dimensional imaging MS of lipids in atherosclerotic plaques: Open-source methods for reconstruction and analysis. Proteomics 2016, 16, 1642–1651. [Google Scholar] [CrossRef]
- Castro-Perez, J.; Hatcher, N.; Kofi Karikari, N.; Wang, S.-P.; Mendoza, V.; Shion, H.; Millar, A.; Shockcor, J.; Towers, M.; McLaren, D.; et al. In vivo isotopically labeled atherosclerotic aorta plaques in ApoE KO mice and molecular profiling by matrix-assisted laser desorption/ionization mass spectrometric imaging. Rapid Commun. Mass Spectrom. 2014, 28, 2471–2479. [Google Scholar] [CrossRef]
- Shen, L.; Yamamoto, T.; Tan, X.W.; Ogata, K.; Ando, E.; Ozeki, E.; Matsuura, E. Identification and visualization of oxidized lipids in atherosclerotic plaques by microscopic imaging mass spectrometry-based metabolomics. Atherosclerosis 2020, 311, 1–12. [Google Scholar] [CrossRef]
- Cao, J.; Goossens, P.; Martin-Lorenzo, M.; Dewez, F.; Claes, B.S.R.; Biessen, E.A.L.; Heeren, R.M.A.; Balluff, B. Atheroma-Specific Lipids in ldlr-/- and apoe-/- Mice Using 2D and 3D Matrix-Assisted Laser Desorption/Ionization Mass Spectrometry Imaging. J. Am. Soc. Mass Spectrom. 2020, 31, 1825–1832. [Google Scholar] [CrossRef] [PubMed]
- Martin-Lorenzo, M.; Balluff, B.; Maroto, A.S.; Carreira, R.J.; van Zeijl, R.J.M.; Gonzalez-Calero, L.; de la Cuesta, F.; Barderas, M.G.; Lopez-Almodovar, L.F.; Padial, L.R.; et al. Molecular anatomy of ascending aorta in atherosclerosis by MS Imaging: Specific lipid and protein patterns reflect pathology. J. Proteom. 2015, 126, 245–251. [Google Scholar] [CrossRef]
- Martin-Lorenzo, M.; Balluff, B.; Maroto, A.S.; Carreira, R.J.; van Zeijl, R.J.M.; Gonzalez-Calero, L.; de la Cuesta, F.; Barderas, M.G.; Lopez-Almodovar, L.F.; Padial, L.R.; et al. Lipid and protein maps defining arterial layers in atherosclerotic aorta. Data Br. 2015, 4, 328–331. [Google Scholar] [CrossRef] [PubMed]
- Visscher, M.; Moerman, A.M.; Burgers, P.C.; Van Beusekom, H.M.M.; Luider, T.M.; Verhagen, H.J.M.; Van der Steen, A.F.W.; Van der Heiden, K.; Van Soest, G. Data Processing Pipeline for Lipid Profiling of Carotid Atherosclerotic Plaque with Mass Spectrometry Imaging. J. Am. Soc. Mass Spectrom. 2019, 30, 1790–1800. [Google Scholar] [CrossRef]
- Moerman, A.M.; Visscher, M.; Slijkhuis, N.; Van Gaalen, K.; Heijs, B.; Klein, T.; Burgers, P.C.; De Rijke, Y.B.; Van Beusekom, H.M.M.; Luider, T.M.; et al. Lipid signature of advanced human carotid atherosclerosis assessed by mass spectrometry imaging. J. Lipid Res. 2021, 62, 100020. [Google Scholar] [CrossRef] [PubMed]
- McMillen, J.C.; Fincher, J.A.; Klein, D.R.; Spraggins, J.M.; Caprioli, R.M. Effect of MALDI matrices on lipid analyses of biological tissues using MALDI-2 postionization mass spectrometry. J. Mass Spectrom. 2020, 55, e4663. [Google Scholar] [CrossRef]
- Eiersbrock, F.B.; Orthen, J.M.; Soltwisch, J. Validation of MALDI-MS imaging data of selected membrane lipids in murine brain with and without laser postionization by quantitative nano-HPLC-MS using laser microdissection. Anal. Bioanal. Chem. 2020, 412, 6875–6886. [Google Scholar] [CrossRef]
- McDonnell, L.A.; van Remoortere, A.; de Velde, N.; van Zeijl, R.J.M.; Deelder, A.M. Imaging mass spectrometry data reduction: Automated feature identification and extraction. J. Am. Soc. Mass Spectrom. 2010, 21, 1969–1978. [Google Scholar] [CrossRef]
- Tabas, I. Macrophage apoptosis in atherosclerosis: Consequences on plaque progression and the role of endoplasmic reticulum stress. Antioxidants Redox Signal. 2009, 11, 2333–2339. [Google Scholar] [CrossRef] [PubMed]
- Clarke, M.C.H.; Figg, N.; Maguire, J.J.; Davenport, A.P.; Goddard, M.; Littlewood, T.D.; Bennett, M.R. Apoptosis of vascular smooth muscle cells induces features of plaque vulnerability in atherosclerosis. Nat. Med. 2006, 12, 1075–1080. [Google Scholar] [CrossRef] [PubMed]
- Matthews, C.; Gorenne, I.; Scott, S.; Figg, N.; Kirkpatrick, P.; Ritchie, A.; Goddard, M.; Bennett, M. Vascular smooth muscle cells undergo telomere-based senescence in human atherosclerosis: Effects of telomerase and oxidative stress. Circ. Res. 2006, 99, 156–164. [Google Scholar] [CrossRef] [PubMed]
- Chakraborty, M.; Jiang, X.-C. Sphingomyelin and Its Role in Cellular Signalling. In Lipid-mediated Protein Signalling. Advances in Experimental Medicine and Biology; Capelluto, D., Ed.; Springer Science+Business Media: Doxrdrecht, The Netherlands, 2013; Volume 991, ISBN 9789400763302. [Google Scholar]
- Slotte, J.P. Biological functions of sphingomyelins. Prog. Lipid Res. 2013, 52, 424–437. [Google Scholar] [CrossRef] [PubMed]
- Augé, N.; Nègre-Salvayre, A.; Salvayre, R.; Levade, T. Sphingomyelin metabolites in vascular cell signaling and atherogenesis. Prog. Lipid Res. 2000, 39, 207–229. [Google Scholar] [CrossRef]
- Park, T.S.; Panek, R.L.; Mueller, S.B.; Hanselman, J.C.; Rosebury, W.S.; Robertson, A.W.; Kindt, E.K.; Homan, R.; Karathanasis, S.K.; Rekhter, M.D. Inhibition of sphingomyelin synthesis reduces atherogenesis in apolipoprotein E-knockout mice. Circulation 2004, 110, 3465–3471. [Google Scholar] [CrossRef]
- Hojjati, M.R.; Li, Z.; Zhou, H.; Tang, S.; Huan, C.; Ooi, E.; Lu, S.; Jiang, X.C. Effect of myriocin on plasma sphingolipid metabolism and atherosclerosis in apoE-deficient mice. J. Biol. Chem. 2005, 280, 10284–10289. [Google Scholar] [CrossRef]
- Li, Y.; Huang, T.; Lou, B.; Ye, D.; Qi, X.; Li, X.; Hu, S.; Ding, T.; Chen, Y.; Cao, Y.; et al. Discovery, synthesis and anti-atherosclerotic activities of a novel selective sphingomyelin synthase 2 inhibitor. Eur. J. Med. Chem. 2019, 163, 864–882. [Google Scholar] [CrossRef]
- Li, Z.; Basterr, M.J.; Hailemariam, T.K.; Hojjati, M.R.; Lu, S.; Liu, J.; Liu, R.; Zhou, H.; Jiang, X.C. The effect of dietary sphingolipids on plasma sphingomyelin metabolism and atherosclerosis. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2005, 1735, 130–134. [Google Scholar] [CrossRef]
- Jiang, X.C.; Paultre, F.; Pearson, T.A.; Reed, R.G.; Francis, C.K.; Lin, M.; Berglund, L.; Tall, A.R. Plasma sphingomyelin level as a risk factor for coronary artery disease. Arterioscler. Thromb. Vasc. Biol. 2000, 20, 2614–2618. [Google Scholar] [CrossRef]
- Liu, J.; Huan, C.; Chakraborty, M.; Zhang, H.; Lu, D.; Kuo, M.-S.; Cao, G.; Jiang, X. Macrophage Sphingomyelin Synthase 2 (SMS2) Deficiency Decreases Atherosclerosis in Mice. Circ. Res. 2009, 105, 295–303. [Google Scholar] [CrossRef]
- Yu, X.H.; Fu, Y.C.; Zhang, D.W.; Yin, K.; Tang, C.K. Foam cells in atherosclerosis. Clin. Chim. Acta 2013, 424, 245–252. [Google Scholar] [CrossRef] [PubMed]
- Guerrini, V.; Gennaro, M.L. Foam Cells: One Size Doesn’t Fit All. Trends Immunol. 2019, 40, 1163–1179. [Google Scholar] [CrossRef]
- Chen, Z.; Ichetovkin, M.; Kurtz, M.; Zycband, E.; Kawka, D.; Woods, J.; He, X.; Plump, A.S.; Hailman, E. Cholesterol in human atherosclerotic plaque is a marker for underlying disease state and plaque vulnerability. Lipids Health Dis. 2010, 9, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, T.; Kobayashi, T.; Kamata, K. Role of Lysophosphatidylcholine (LPC) in Atherosclerosis. Curr. Med. Chem. 2007, 14, 3209–3220. [Google Scholar] [CrossRef] [PubMed]
- Schmitz, G.; Ruebsaamen, K. Metabolism and atherogenic disease association of lysophosphatidylcholine. Atherosclerosis 2010, 208, 10–18. [Google Scholar] [CrossRef]
- Diehl, P.; Nienaber, F.; Zaldivia, M.T.K.; Stamm, J.; Siegel, P.M.; Mellett, N.A.; Wessinger, M.; Wang, X.; McFadyen, J.D.; Bassler, N.; et al. Lysophosphatidylcholine is a Major Component of Platelet Microvesicles Promoting Platelet Activation and Reporting Atherosclerotic Plaque Instability. Thromb. Haemost. 2019, 119, 1295–1310. [Google Scholar] [CrossRef]
- van der Veen, J.N.; Kennelly, J.P.; Wan, S.; Vance, J.E.; Vance, D.E.; Jacobs, R.L. The critical role of phosphatidylcholine and phosphatidylethanolamine metabolism in health and disease. Biochim. Biophys. Acta Biomembr. 2017, 1859, 1558–1572. [Google Scholar] [CrossRef]
- Allahverdian, S.; Chehroudi, A.C.; McManus, B.M.; Abraham, T.; Francis, G.A. Contribution of intimal smooth muscle cells to cholesterol accumulation and macrophage-like cells in human atherosclerosis. Circulation 2014, 129, 1551–1559. [Google Scholar] [CrossRef]
- Wang, Y.; Dubland, J.A.; Allahverdian, S.; Asonye, E.; Sahin, B.; Jaw, J.E.; Sin, D.D.; Seidman, M.A.; Leeper, N.J.; Francis, G.A. Smooth Muscle Cells Contribute the Majority of Foam Cells in ApoE (Apolipoprotein E)-Deficient Mouse Atherosclerosis. Arterioscler. Thromb. Vasc. Biol. 2019, 39, 876–887. [Google Scholar] [CrossRef]
- Stary, H.C.; Chandler, A.B.; Dinsmore, R.E.; Fuster, V.; Glagov, S.; Insull, W.; Rosenfeld, M.E.; Schwartz, C.J.; Wagner, W.D.; Wissler, R.W. A Definition of Advanced Types of Atherosclerotic Lesions and a Histological Classification of Atherosclerosis. Circulation 1995, 92, 1355–1374. [Google Scholar] [CrossRef] [PubMed]
- Stary, H.; Chandler, A.; Glagov, S.; Jr, G.; Insull, W.; Rosenfeld, M.J.; Schaffer, S.; Schwartz, C.; Wagner, W.; Wissler, R. A Definition of Initial, Fatty Streak, and Intermediate Lesions of Atherosclerosis. A report from the Committee on Vascular Lesions of the Council on Atherosclerosis, American Heart Association. Circulation 1994, 89, 2462–2478. [Google Scholar] [CrossRef]
- Pucci, A.; Franzini, M.; Matteucci, M.; Ceragioli, S.; Marconi, M.; Ferrari, M.; Passino, C.; Basolo, F.; Emdin, M.; Paolicchi, A. B-Gamma-glutamyltransferase activity in human vulnerable carotid plaques. Atherosclerosis 2014, 237, 307–313. [Google Scholar] [CrossRef] [PubMed]
- Belov, M.E.; Ellis, S.R.; Dilillo, M.; Paine, M.R.L.; Danielson, W.F.; Anderson, G.A.; De Graaf, E.L.; Eijkel, G.B.; Heeren, R.M.A.; McDonnell, L.A. Design and Performance of a Novel Interface for Combined Matrix-Assisted Laser Desorption Ionization at Elevated Pressure and Electrospray Ionization with Orbitrap Mass Spectrometry. Anal. Chem. 2017, 89, 7493–7501. [Google Scholar] [CrossRef] [PubMed]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of Image Analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef] [PubMed]
- Shoelson, B. Image Registration App 2020. Available online: https://it.mathworks.com/matlabcentral/fileexchange/34510-image-registration-app (accessed on 29 April 2019).
- He, L.; Diedrich, J.; Chu, Y.-Y.; Yates, J.R. Extracting Accurate Precursor Information for Tandem Mass Spectra by RawConverter. Anal. Chem. 2015, 87, 11361–11367. [Google Scholar] [CrossRef]
- Alexandrov, T.; Becker, M.; Deininger, S.O.; Ernst, G.; Wehder, L.; Grasmair, M.; Von Eggeling, F.; Thiele, H.; Maass, P. Spatial segmentation of imaging mass spectrometry data with edge-preserving image denoising and clustering. J. Proteome Res. 2010, 9, 6535–6546. [Google Scholar] [CrossRef] [PubMed]
- Lourakis, M. TV-L1 Image Denoising Algorithm. Available online: https://www.mathworks.com/matlabcentral/fileexchange/57604-tv-l1-image-denoising-algorithm (accessed on 15 May 2019).
- Heberle, H.; Meirelles, V.G.; da Silva, F.R.; Telles, G.P.; Minghim, R. InteractiVenn: A web-based tool for the analysis of sets through Venn diagrams. BMC Bioinform. 2015, 16, 1–7. [Google Scholar] [CrossRef]
- Tran, T.; Szymańska, E.; Gerretzen, J.; Buydens, L.; Afanador, N.L.; Blanchet, L. Weight randomization test for the selection of the number of components in PLS models. J. Chemom. 2017, 31, 1–15. [Google Scholar] [CrossRef]
- Farrés, M.; Platikanov, S.; Tsakovski, S.; Tauler, R. Comparison of the variable importance in projection (VIP) and of the selectivity ratio (SR) methods for variable selection and interpretation. J. Chemom. 2015, 29, 528–536. [Google Scholar] [CrossRef]
- Smith, C.A.; Maille, G.O.; Want, E.J.; Qin, C.; Trauger, S.A.; Brandon, T.R.; Custodio, D.E.; Abagyan, R.; Siuzdak, G. Metlin: A Metabolite Mass Spectral Database. Ther. Drug Monit. 2005, 27, 747–751. [Google Scholar] [CrossRef] [PubMed]
- Liebisch, G.; Vizcaíno, J.A.; Köfeler, H.; Trötzmüller, M.; Griffiths, W.J.; Schmitz, G.; Spener, F.; Wakelam, M.J.O. Shorthand notation for lipid structures derived from mass spectrometry. J. Lipid Res. 2013, 54, 1523–1530. [Google Scholar] [CrossRef] [PubMed]
- Perez-Riverol, Y.; Csordas, A.; Bai, J.; Bernal-Llinares, M.; Hewapathirana, S.; Kundu, D.J.; Inuganti, A.; Griss, J.; Mayer, G.; Eisenacher, M.; et al. The PRIDE database and related tools and resources in 2019: Improving support for quantification data. Nucleic Acids Res. 2019, 47, D442–D450. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Matrix | Dysregulation in Symptomatic | m/z | Δppm | Adduct | Assignment |
|---|---|---|---|---|---|
| DHB | ↑ | 369.35 | 5 | [M+H-H2O]+ | Cholesterol |
| DHB | ↑ | 520.34 | 0 | [M+H]+ | LPC(18:2); |
| DHB | ↑ | 542.32 | 3 | [M+Na]+ | LPC(18:2) |
| DHB | ↑ | 544.34 | 0 | [M+H]+ | LPC(20:4) |
| DHB | ↑ | 544.34 | 0 | [M+H-H2O]+ | PC(20:2) |
| DHB | ↑ | 666.48 | |||
| DHB | ↑ | 671.575 | 1 | [M+H]+ | CE(20:5) |
| DHB | ↑ | 671.575 | 1 | [M+Na]+ | CE(18:2), zymosteryl oleate, 16:1 Stigmasteryl ester, 16:2 Sitosteryl ester |
| DHB | ↑ | 711.54 | 1 | [M+Na]+ | Etn-1-P-Cer(22:1), SM(33:1), PE-Cer(36:1) |
| DHB | ↑ | 723.54 | 1 | [M+Na]+ | SM(34:2), PE-Cer(37:2) |
| DHB | ↑ | 725.555 | 2 | [M+Na]+ | SM(34:1), PE-Cer(37:1) |
| DHB | ↑ | 741.53 | |||
| DHB | ↑ | 807.635 | 0 | [M+Na]+ | SM(40:2) |
| DHB | ↑ | 833.65 | 0 | [M+Na]+ | SM(42:3) |
| Nor. | ↑ | 723.54 | 1 | [M+Na]+ | SM(34:2), PE-Cer(37:2) |
| Nor. | ↑ | 835.67 | |||
| DHB | ↓ | 697.475 | 4 | [M+Na]+ | PA(34:1) |
| DHB | ↓ | 723.49 | 4 | [M+Na]+ | PA(36:2) |
| DHB | ↓ | 756.55 | 1 | [M+Na]+ | PC(32:0), PE(35:0) |
| DHB | ↓ | 780.55 | 1 | [M+Na]+ | PC(34:2), PE(37:2) |
| DHB | ↓ | 782.565 | 2 | [M+Na]+ | PC(34:1), PE(37:1) |
| DHB | ↓ | 804.55 | 1 | [M+Na]+ | PC(36:4), PE(39:4) |
| DHB | ↓ | 806.565 | 2 | [M+Na]+ | PC(36:3), PE(39:3) |
| DHB | ↓ | 808.58 | 3 | [M+Na]+ | PC(36:2), PE(39:2) |
| DHB | ↓ | 832.58 | 3 | [M+Na]+ | PC(38:4), PE(41:4) |
| Nor. | ↓ | 758.57 | 0 | [M+H]+ | PC(34:2), PE(37:2) |
| Nor. | ↓ | 758.57 | 0 | [M+H-H2O]+ | PS(O-36:0) |
| Nor. | ↓ | 782.57 | 0 | [M+H]+ | PC(36:4), PE(39:4) |
| Nor. | ↓ | 782.57 | 0 | [M+H-H2O]+ | PS(O-38:3), PS(P-38:2) |
| Nor. | ↓ | 808.585 | 0 | [M+H]+ | PC(38:5), PE(41:5) |
| Nor. | ↓ | 808.585 | 0 | [M+H-H2O]+ | PS(O-40:4), PS(P-40:3) |
| Nor. | ↓ | 808.585 | 2 | [M+Na]+ | PC(36:2), PE(39:2) |
| Matrix | Dysregulation in Symptomatic | m/z | Δppm | Adduct | Assignment |
|---|---|---|---|---|---|
| DHB | ↑ | 369.35 | 5 | [M+H-H2O]+ | Cholesterol |
| DHB | ↑ | 671.575 | 1 | [M+H]+ | CE(20:5) |
| DHB | ↑ | 671.575 | 1 | [M+Na]+ | CE(18:2), zymosteryl oleate, 16:1 Stigmasteryl ester, 16:2 Sitosteryl ester |
| DHB | ↑ | 687.55 | 4 | [M+Na]+ | TG(38:1) |
| DHB | ↑ | 725.555 | 2 | [M+Na]+ | SM(34:1), PE-Cer(37:1) |
| DHB | ↑ | 741.53 | |||
| DHB | ↑ | 758.57 | 0 | [M+H]+ | PC(34:2), PE(37:2) |
| DHB | ↑ | 780.55 | 1 | [M+Na]+ | PC(34:2), PE(37:2) |
| DHB | ↑ | 782.565 | 2 | [M+Na]+ | PC(34:1), PE(37:1) |
| DHB | ↑ | 796.525 | 0 | [M+Na]+ | PE(P-40:7),1-(8-[3]-ladderane-octanoyl)-2-(8-[3]-ladderane-octanyl)-sn-glycerophosphoethanolamine |
| DHB | ↑ | 798.54 | 1 | [M+Na]+ | PE(P-40:6) |
| DHB | ↑ | 808.58 | 3 | [M+Na]+ | PC(36:2), PE(39:2) |
| Nor. | ↑ | 780.555 | 1 | [M+H]+ | PC(36:5), PE(39:5) |
| Nor. | ↑ | 780.555 | 4 | [M+Na]+ | PC(34:2), PE(37:2) |
| Nor. | ↑ | 808.585 | 0 | [M+H]+ | PC(38:5), PE(41:5) |
| Nor. | ↑ | 808.585 | 0 | [M+H-H2O]+ | PS(O-40:4), PS(P-40:3) |
| Nor. | ↓ | 808.585 | 2 | [M+Na]+ | PC(36:2), PE(39:2) |
| DHB | ↓ | 697.475 | 4 | [M+Na]+ | PA(34:1) |
| DHB | ↓ | 773.505 | 1 | [M+H-H2O]+ | all-trans-nonaprenyl diphosphate |
| DHB | ↓ | 778.605 | |||
| DHB | ↓ | 832.58 | 3 | [M+Na]+ | PC(38:4), PE(41:4) |
| DHB | ↓ | 837.68 | 2 | [M+Na]+ | SM(42:1) |
| DHB | ↓ | 946.615 | |||
| Nor. | ↓ | 723.54 | 1 | [M+Na]+ | SM(34:2), PE-Cer(37:2) |
| Nor. | ↓ | 835.67 | 4 | [M+Na]+ | SM(42:2) |
| Patient | Age | Sex | Symptoms | Plaque AHA Classification |
|---|---|---|---|---|
| 1 | 66 | F | None | Type VI |
| 2 | 84 | F | None | Type Va |
| 3 | 84 | M | None | Type III |
| 4 | 68 | M | Transient ischemic attack | Type VI |
| 5 | 75 | M | Amaurosis | Type VI |
| 6 | 73 | F | Transient ischemic attack | Type VI |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Greco, F.; Quercioli, L.; Pucci, A.; Rocchiccioli, S.; Ferrari, M.; Recchia, F.A.; McDonnell, L.A. Mass Spectrometry Imaging as a Tool to Investigate Region Specific Lipid Alterations in Symptomatic Human Carotid Atherosclerotic Plaques. Metabolites 2021, 11, 250. https://doi.org/10.3390/metabo11040250
Greco F, Quercioli L, Pucci A, Rocchiccioli S, Ferrari M, Recchia FA, McDonnell LA. Mass Spectrometry Imaging as a Tool to Investigate Region Specific Lipid Alterations in Symptomatic Human Carotid Atherosclerotic Plaques. Metabolites. 2021; 11(4):250. https://doi.org/10.3390/metabo11040250
Chicago/Turabian StyleGreco, Francesco, Laura Quercioli, Angela Pucci, Silvia Rocchiccioli, Mauro Ferrari, Fabio A. Recchia, and Liam A. McDonnell. 2021. "Mass Spectrometry Imaging as a Tool to Investigate Region Specific Lipid Alterations in Symptomatic Human Carotid Atherosclerotic Plaques" Metabolites 11, no. 4: 250. https://doi.org/10.3390/metabo11040250
APA StyleGreco, F., Quercioli, L., Pucci, A., Rocchiccioli, S., Ferrari, M., Recchia, F. A., & McDonnell, L. A. (2021). Mass Spectrometry Imaging as a Tool to Investigate Region Specific Lipid Alterations in Symptomatic Human Carotid Atherosclerotic Plaques. Metabolites, 11(4), 250. https://doi.org/10.3390/metabo11040250