
Contribution of Metabolomics to the Understanding of NAFLD and NASH Syndromes: A Systematic Review

 ,
,  ,
,  , ,
, ,  and
and 
Abstract
:1. Introduction
1.1. MetS General Characteristics
1.2. NAFLD and NASH
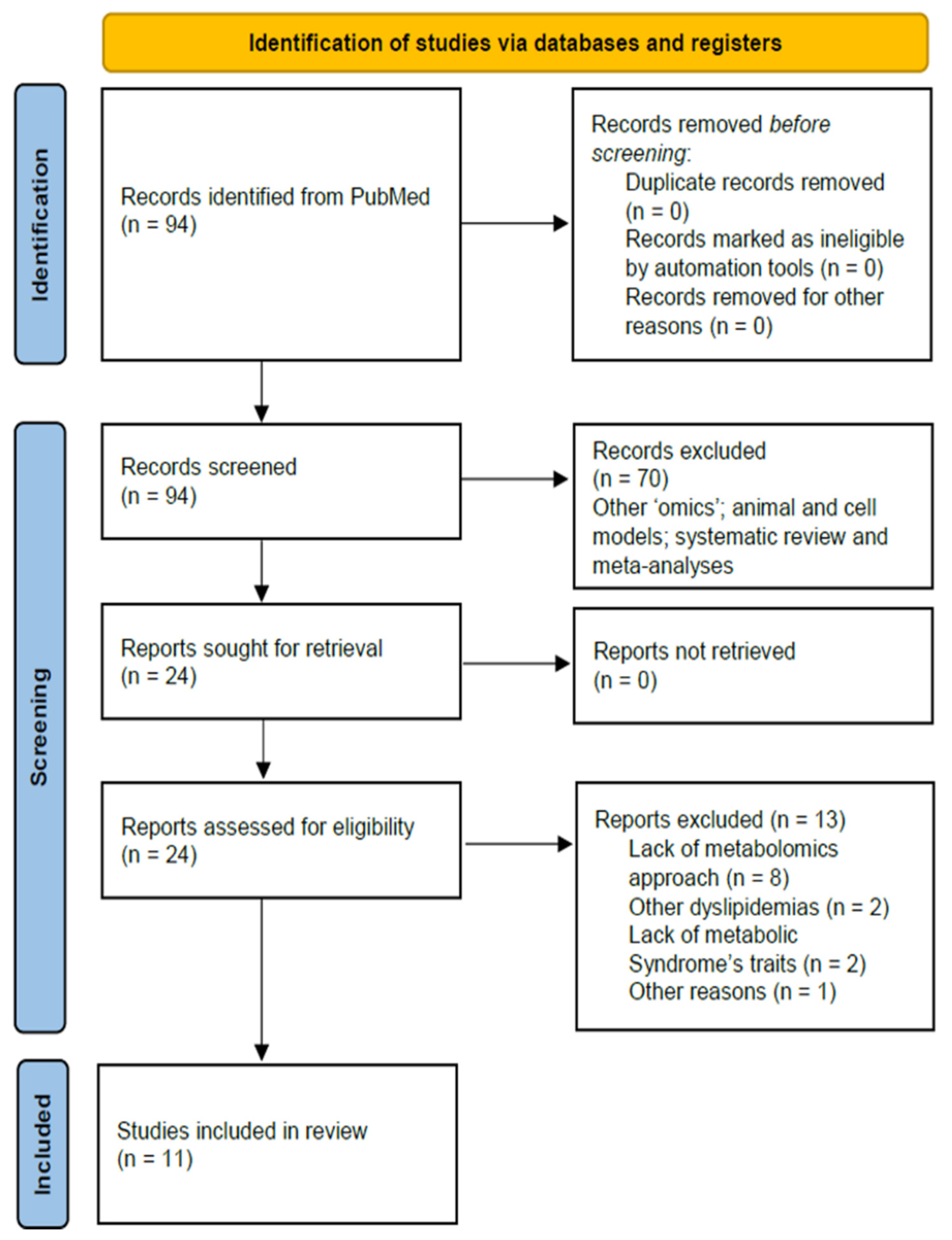
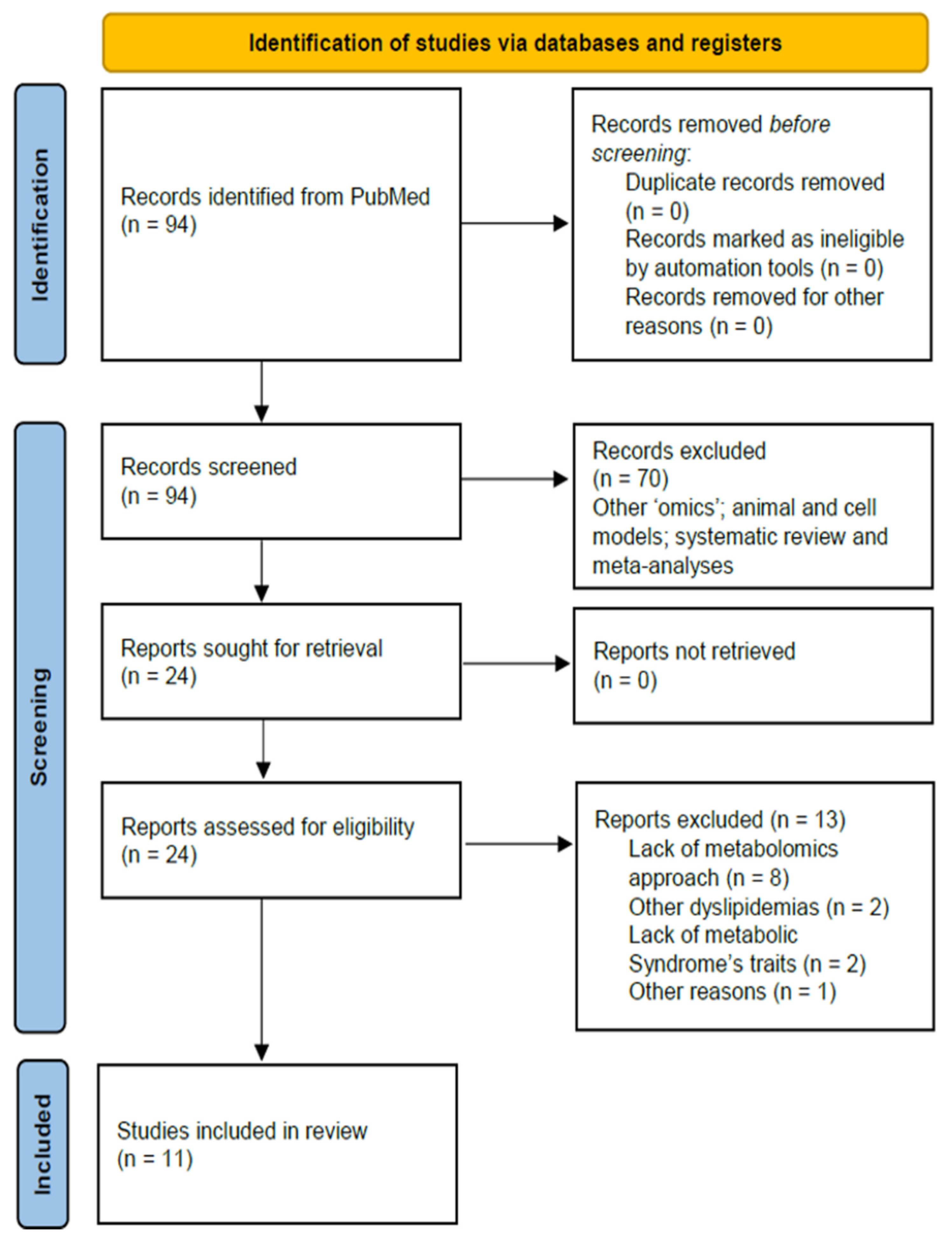
2. Methods
2.1. Search Strategy
2.2. Selection Criteria
2.3. Data Extraction
3. Results
3.1. NAFLD/NASH Biomarkers: Results from Case/Control Studies
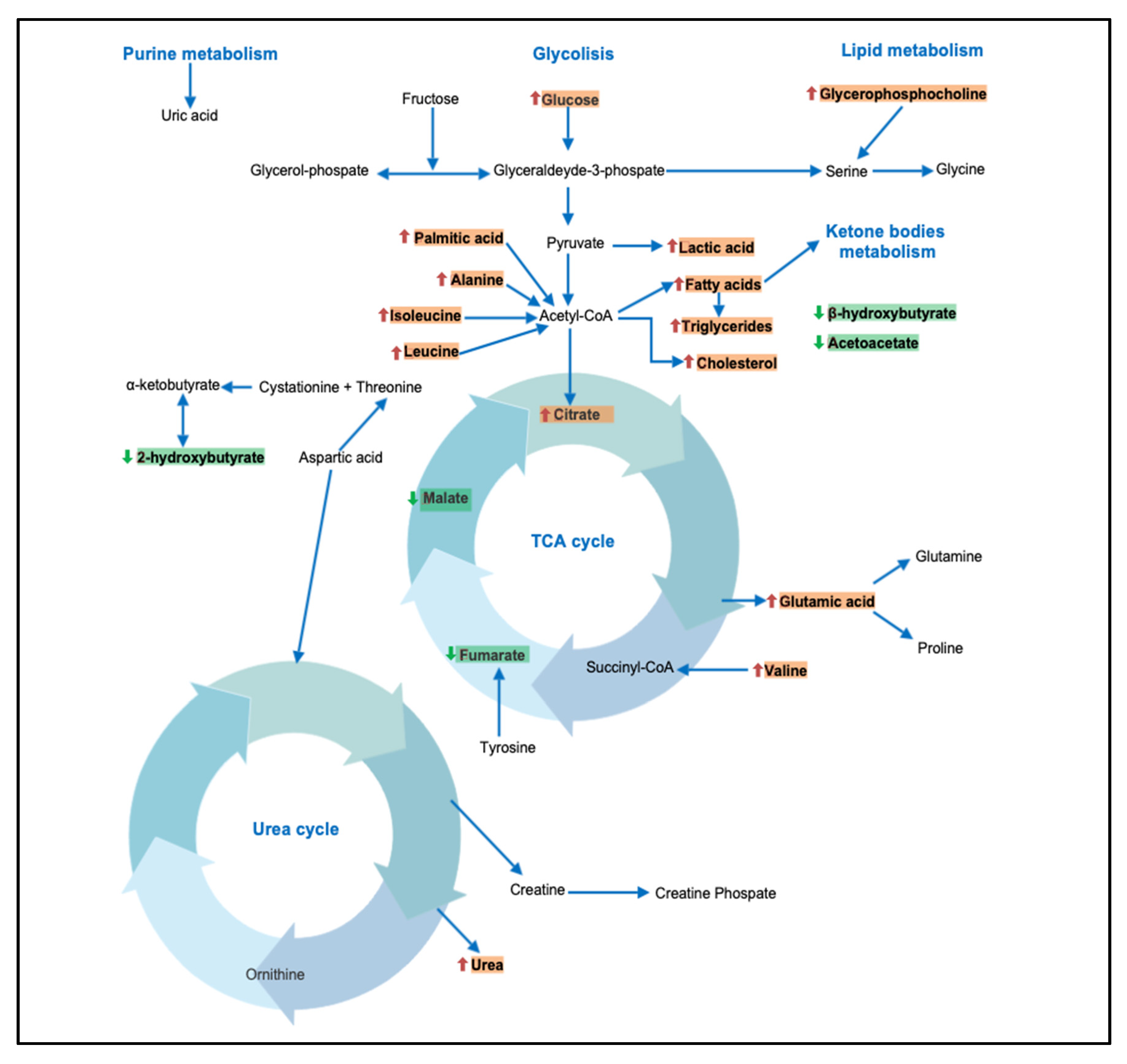
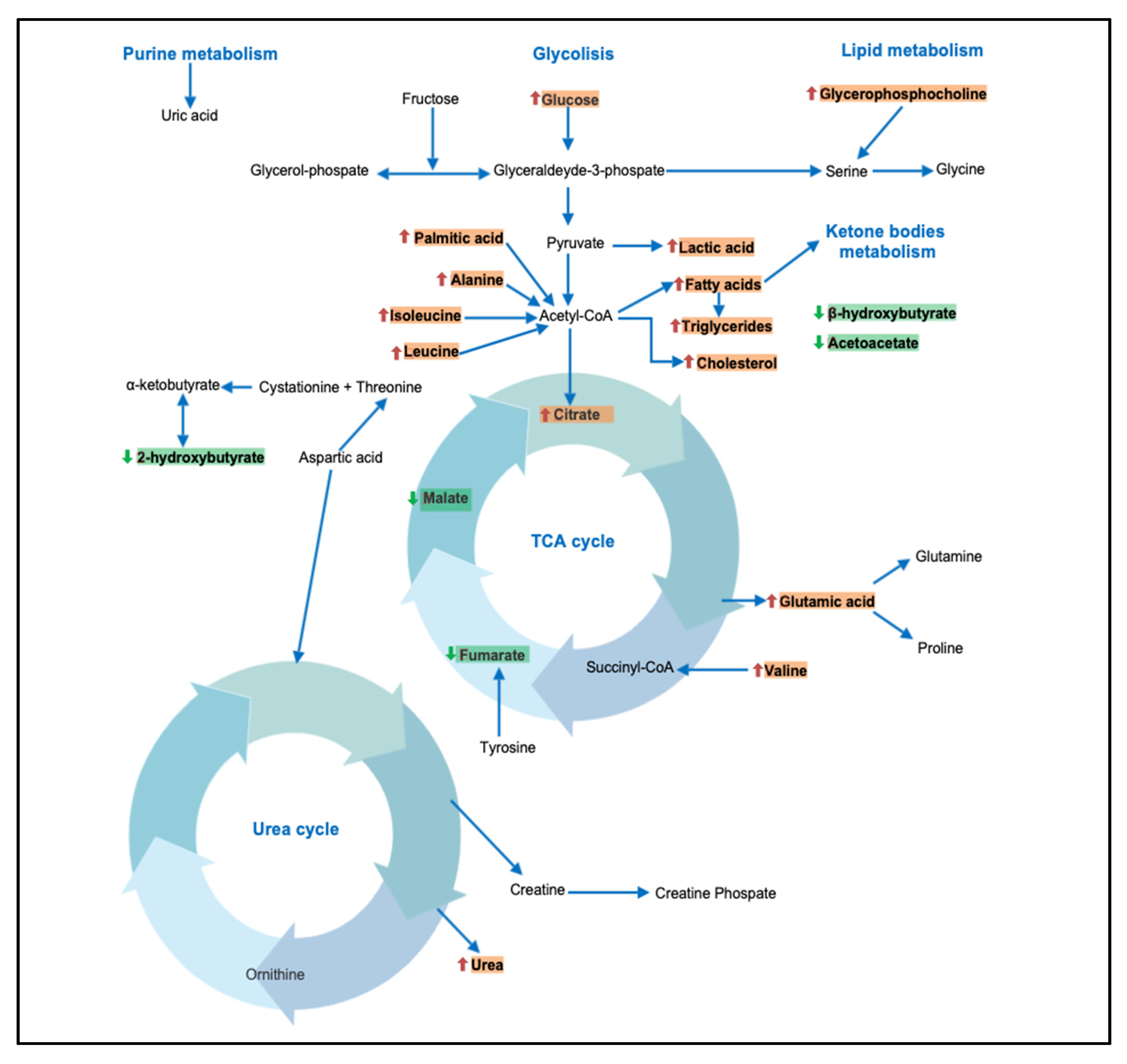
3.2. Metabolism of Amino Acids and Derivatives
3.3. Metabolism of Fatty Acids and Derivatives
3.4. Metabolism of Vitamin and Ketone Bodies
4. Discussion
4.1. BCAAs and Kynurenine
4.2. Carbohydrates
4.3. Fatty Acids and Derivatives
4.4. Other Metabolites
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Lent-Schochet, D.; McLaughlin, M.; Ramakrishnan, N.; Jialal, I. Exploratory metabolomics of metabolic syndrome: A status report. World J. Diabetes 2019, 10, 23–36. [Google Scholar] [CrossRef]
- Alberti, K.G.; Zimmet, P.Z. Definition, diagnosis and classification of diabetes mellitus and its complications, part 1: Diagnosis and classification of diabetes mellitus provisional report of a WHO consultation. Diabet. Med. 1998, 15, 539–553. [Google Scholar] [CrossRef]
- Cornier, M.A.; Dabelea, D.; Hernandez, T.L.; Lindstrom, R.C.; Steig, A.J.; Stob, N.R.; Van Pelt, R.E.; Wang, H.; Eckel, R.H. The Metabolic Syndrome. Endocr. Rev. 2008, 29, 777–822. [Google Scholar] [CrossRef]
- Grund, S.M.; Cleeman, J.I.; Daniels, S.R.; Donato, K.A.; Eckel, R.H.; Franklin, B.A.; Gordon, D.J.; Krauss, R.M.; Savage, P.J.; Smith, S.C.; et al. Diagnosis and management of the metabolic syndrome: An American Heart Association/National Heart, Lung, and Blood Institute scientific statement. Circulation 2005, 112, 2735–2752. [Google Scholar] [CrossRef] [Green Version]
- Cleeman, J.I.; Grundy, S.M.; Becker, D.; Clark, L. Expert Panel on Detection, Evaluation, and Treatment of High Blood Cholesterol in Adults. Executive Summary of The Third Report of The National Cholesterol Education Program (NCEP). Expert Panel on Detection, Evaluation, And Treatment of High Blood Cholesterol in Adults (Adult Treatment Panel III). JAMA 2001, 285, 2486–2497. [Google Scholar]
- Hamaguchi, M.; Kojima, T.; Takeda, N.; Nakagawa, T.; Taniguchi, H.; Fujii, K.; Omatsu, T.; Nakajima, T.; Sarui, H.; Shimazaki, M.; et al. The metabolic syndrome as a predictor of nonalcoholic fatty liver disease. Ann. Intern. Med. 2005, 143, 722–728. [Google Scholar] [CrossRef]
- Hanley, A.J.; Williams, K.; Festa, A.; Wagenknecht, L.E.; D’Agostino, R.B.; Haffner, S.M. Liver markers and development of the metabolic syndrome: The insulin resistance atherosclerosis study. Diabetes 2005, 54, 3140–3147. [Google Scholar] [CrossRef] [Green Version]
- Marra, F.; Svegliati-Baroni, G. Lipotoxicity and the gut-liver axis in NASH pathogenesis. J. Hepatol. 2018, 68, 280–295. [Google Scholar] [CrossRef]
- Sookoian, S.; Pirola, C.J. Genetic predisposition in nonalcoholic fatty liver disease. Clin. Mol. Hepatol. 2017, 23, 1–12. [Google Scholar] [CrossRef]
- Lind, L. Genome-Wide Association Study of the Metabolic Syndrome in UK Biobank. Metab. Syndr. Relat. Disord. 2019, 17, 505–511. [Google Scholar] [CrossRef]
- Männistö, V.T.; Simonen, M.; Hyysalo, J.; Soininen, P.; Kangas, A.J.; Kaminska, D.; Matte, A.K.; Venesmaa, S.; Käkelä, P.; Kärjä, V.; et al. Ketone body production is differentially altered in steatosis and non-alcoholic steatohepatitis in obese humans. Liver Int. 2015, 35, 1853–1861. [Google Scholar] [CrossRef]
- Stechemesser, L.; Eder, S.K.; Wagner, A.; Patsch, W.; Feldman, A.; Strasser, M.; Auer, S.; Niederseer, D.; Huber-Schönauer, U.; Paulweber, B.; et al. Metabolomic profiling identifies potential pathways involved in the interaction of iron homeostasis with glucose metabolism. Mol. Metab. 2016, 6, 38–47. [Google Scholar] [CrossRef]
- Sookoian, S.; Castaño, G.O.; Scian, R.; Fernández Gianotti, T.; Dopazo, H.; Rohr, C.; Gaj, G.; San Martino, J.; Sevic, I.; Flichman, D.; et al. Serum aminotransferases in nonalcoholic fatty liver disease are a signature of liver metabolic perturbations at the amino acid and Krebs cycle level. Am. J. Clin. Nutr. 2016, 103, 422–434. [Google Scholar] [CrossRef] [Green Version]
- Bhupathiraju, S.N.; Guasch-Ferré, M.; Gadgil, M.D.; Newgard, C.B.; Bain, J.R.; Muehlbauer, M.J.; Ilkayeva, O.R.; Scholtens, D.M.; Hu, F.B.; Kanaya, A.M.; et al. Dietary Patterns among Asian Indians Living in the United States Have Distinct Metabolomic Profiles That Are Associated with Cardiometabolic Risk. J. Nutr. 2018, 148, 1150–1159. [Google Scholar] [CrossRef]
- Feldman, A.; Eder, S.K.; Felder, T.K.; Paulweber, B.; Zandanell, S.; Stechemesser, L.; Schranz, M.; Strebinger, G.; Huber-Schönauer, U.; Niederseer, D.; et al. Clinical and metabolic characterization of obese subjects without non-alcoholic fatty liver: A targeted metabolomics approach. Diabetes Metab. 2019, 45, 132–139. [Google Scholar] [CrossRef]
- Troisi, J.; Belmonte, F.; Bisogno, A.; Pierri, L.; Colucci, A.; Scala, G.; Cavallo, P.; Mandato, C.; Di Nuzzi, A.; Di Michele, L.; et al. Metabolomic Salivary Signature of Pediatric Obesity Related Liver Disease and Metabolic Syndrome. Nutrients 2019, 26, 274. [Google Scholar] [CrossRef] [Green Version]
- Masarone, M.; Troisi, J.; Aglitti., A.; Torre, P.; Colucci, A.; Dallio, M.; Federico, A.; Balsano, C.; Persico, M. Untargeted metabolomics as a diagnostic tool in NAFLD: Discrimination of statosis, steatohepatitis and cirrhosis. Metabolomics 2021, 17, 1–13. [Google Scholar] [CrossRef]
- Lovric, A.; Granér, M.; Bjornson, E.; Arif, M.; Benfeitas, R.; Nyman, K.; Ståhlman, M.; Pentikäinen, M.O.; Lundbom, J.; Hakkarainen, A.; et al. Characterization of different fat depots in NAFLD using inflammation-associated proteome, lipidome and metabolome. Sci. Rep. 2018, 8, 14200. [Google Scholar] [CrossRef] [Green Version]
- Ni, Y.; Zhao, L.; Yu, H.; Ma, X.; Bao, Y.; Rajani, C.; Loo, L.W.; Shvetsov, Y.B.; Yu, H.; Chen, T.; et al. Circulating Unsaturated Fatty Acids Delineate the Metabolic Status of Obese Individuals. EBioMedicine 2015, 2, 1513–1522. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Y.; Orešič, M.; Leivonen, M.; Gopalacharyulu, P.; Hyysalo, J.; Arola, J.; Verrijken, A.; Francque, S.; Van Gaal, L.; Hyötyläinen, T.; et al. Noninvasive Detection of Nonalcoholic Steatohepatitis Using Clinical Markers and Circulating Levels of Lipids and Metabolites. Clin. Gastroenterol. Hepatol. 2016, 14, 1463–1472. [Google Scholar] [CrossRef] [Green Version]
- Zhong, G.; Kirkwood, J.; Won, K.J.; Tjota, N.; Jeong, H.; Isoherranen, N. Characterization of Vitamin A Metabolome in Human Livers with and Without Nonalcoholic Fatty Liver Disease. J. Pharm. Exp. Ther. 2019, 370, 92–103. [Google Scholar] [CrossRef]
- Eicher, T.; Kinnebrew, G.; Patt, A.; Spencer, K.; Ying, K.; Ma, Q.; Machiraju, R.; Mathé, E.A. Metabolomics and Multi-Omics Integration: A Survey of Computational Methods and Resources. Metabolites 2020, 10, 202. [Google Scholar] [CrossRef]
- Haufe, S.; Witt, H.; Engeli, S.; Kaminski, J.; Utz, W.; Fuhrmann, J.C.; Rein, D.; Schulz-Menger, J.; Luft, F.C.; Boschmann, M.; et al. Branched-chain and aromatic amino acids, insulin resistance and liver specific ectopic fat storage in overweight to obese subjects. Nutr. Metab. Cardiovasc. Dis. 2016, 26, 637–642. [Google Scholar] [CrossRef]
- Cheng, S.; Rhee, E.P.; Larson, M.G.; Lewis, G.D.; McCabe, E.L.; Shen, D.; Palma, M.J.; Roberts, L.D.; Dejam, A.; Souza, A.L.; et al. Metabolite profiling identifies pathways associated with metabolic risk in humans. Circulation 2012, 125, 2222–2231. [Google Scholar] [CrossRef] [Green Version]
- Lee, C.C.; Watkins, S.M.; Lorenzo, C.; Wagenknecht, L.E.; Il’yasova, D.; Chen, Y.D.; Haffner, S.M.; Hanley, A.J. Branched-Chain Amino Acids and Insulin Metabolism: The Insulin Resistance Atherosclerosis Study (IRAS). Diabetes Care 2016, 39, 582–588. [Google Scholar] [CrossRef] [Green Version]
- Zhang, F.; Zhao, S.; Yan, W.; Xia, Y.; Chen, X.; Wang, W.; Zhang, J.; Gao, C.; Peng, C.; Yan, F.; et al. Branched Chain Amino Acids Cause Liver Injury in Obese/Diabetic Mice by Promoting Adipocyte Lipolysis and Inhibiting Hepatic Autophagy. EBioMedicine 2016, 13, 157–167. [Google Scholar] [CrossRef] [Green Version]
- Oxenkrug, G.F. Metabolic syndrome, age-associated neuroendocrine disorders, and dysregulation of tryptophan-kynurenine metabolism. Ann. N. Y. Acad. Sci. 2010, 1199, 1–14. [Google Scholar] [CrossRef]
- Park, S.; Sadanala, K.C.; Kim, E.K.A. Metabolomic Approach to Understanding the Metabolic Link between Obesity and Diabetes. Mol. Cells 2015, 38, 587–596. [Google Scholar] [CrossRef] [Green Version]
- Ramachandran, R.; Gravenstein, K.S.; Metter, E.J.; Egan, J.M.; Ferrucci, L.; Chia, C.W. Selective contribution of regional adiposity, skeletal muscle, and adipokines to glucose disposal in older adults. J. Am. Geriatr. Soc. 2012, 60, 707–712. [Google Scholar] [CrossRef] [Green Version]
- Böhm, A.; Halama, A.; Meile, T.; Zdichavsky, M.; Lehmann, R.; Weigert, C.; Fritsche, A.; Stefan, N.; Königsrainer, A.; Häring, H.U.; et al. Metabolic signatures of cultured human adipocytes from metabolically healthy versus unhealthy obese individuals. PLoS ONE 2014, 9, e93148. [Google Scholar] [CrossRef]
- Schäfer, N.; Yu, Z.; Wagener, A.; Millrose, M.K.; Reissmann, M.; Bortfeldt, R.; Dieterich, C.; Adamski, J.; Wang-Sattler, R.; Illig, T.; et al. Changes in metabolite profiles caused by genetically determined obesity in mice. Metabolomics 2014, 10, 461–472. [Google Scholar] [CrossRef] [Green Version]
- Spalding, K.L.; Arner, E.; Westermark, P.O.; Bernard, S.; Buchholz, B.A.; Bergmann, O.; Blomqvist, L.; Hoffstedt, J.; Näslund, E.; Britton, T.; et al. Dynamics of fat cell turnover in humans. Nature 2008, 453, 783–787. [Google Scholar] [CrossRef]
- Hanamatsu, H.; Ohnishi, S.; Sakai, S.; Yuyama, K.; Mitsutake, S.; Takeda, H.; Hashino, S.; Igarashi, Y. Altered levels of serum sphingomyelin and ceramide containing distinct acyl chains in young obese adults. Nutr. Diabetes 2014, 4, e141. [Google Scholar] [CrossRef] [Green Version]
- Gong, J.; Campos, H.; McGarvey, S.; Wu, Z.; Goldberg, R.; Baylin, A. Adipose tissue palmitoleic acid and obesity in humans: Does it behave as a lipokine? Am. J. Clin. Nutr. 2011, 93, 186–191. [Google Scholar] [CrossRef] [Green Version]
- Kurotani, K.; Sato, M.; Ejima, Y.; Nanri, A.; Yi, S.; Pham, N.M.; Akter, S.; Poudel-Tandukar, K.; Kimura, Y.; Imaizumi, K.; et al. High levels of stearic acid, palmitoleic acid, and dihomo-γ-linolenic acid and low levels of linoleic acid in serum cholesterol ester are associated with high insulin resistance. Nutr. Res. 2012, 32, 669–675. [Google Scholar] [CrossRef]
- Micha, R.; Mozaffarian, D. Saturated fat and cardiometabolic risk factors, coronary heart disease, stroke, and diabetes: A fresh look at the evidence. Lipids 2010, 45, 893–905. [Google Scholar] [CrossRef] [Green Version]
- Bechmann, L.P.; Kocabayoglu, P.; Sowa, J.P.; Sydor, S.; Best, J.; Schlattjan, M.; Beilfuss, A.; Schmitt, J.; Hannivoort, R.A.; Kilicarslan, A.; et al. Free fatty acids repress small heterodimer partner (SHP) activation and adiponectin counteracts bile acid-induced liver injury in superobese patients with nonalcoholic steatohepatitis. Hepatology 2013, 57, 1394–1406. [Google Scholar] [CrossRef]
- Fukao, T.; Lopaschuk, G.D.; Mitchell, G.A. Pathways and control of ketone body metabolism: On the fringe of lipid biochemistry. Prostaglandins Leukot. Essent. Fat. Acids 2004, 70, 243–251. [Google Scholar] [CrossRef]
- Mulholland, A.J.; Richards, W.G. Acetyl-CoA enolization in citrate synthase: A quantum mechanical/molecular mechanical (QM/MM) study. Proteins 1997, 27, 9–25. [Google Scholar] [CrossRef]
- Amer, B.; Clausen, M.R.; Bertram, H.C.; Bohl, M.; Nebel, C.; Zheng, H.; Skov, T.; Larsen, M.K.; Gregersen, S.; Hermansen, K.; et al. Consumption of Whey in Combination with Dairy Medium-Chain Fatty Acids (MCFAs) may Reduce Lipid Storage due to Urinary Loss of Tricarboxylic Acid Cycle Intermediates and Increased Rates of MCFAs Oxidation. Mol. Nutr. Food Res. 2017, 61, 1601048. [Google Scholar] [CrossRef] [Green Version]
- Martin, D.B.; Vagelos, P.R. The mechanism of tricarboxylic acid cycle regulation of fatty acid synthesis. J. Biol. Chem. 1962, 237, 1787–1792. [Google Scholar] [CrossRef]
- Youm, Y.H.; Nguyen, K.Y.; Grant, R.W.; Goldberg, E.L.; Bodogai, M.; Kim, D.; D’Agostino, D.; Planavsky, N.; Lupfer, C.; Kanneganti, T.D.; et al. The ketone metabolite β-hydroxybutyrate blocks NLRP3 inflammasome-mediated inflammatory disease. Nat. Med. 2015, 21, 263–269. [Google Scholar] [CrossRef] [Green Version]

{kind=link}
{kind=link}
| Author | Year | Title | Subjects | Reasons of Exclusion |
|---|---|---|---|---|
| Amer et al. | 2017 | Consumption of whey in combination with dairy medium-chain fatty acids (MCFAs) may reduce lipid storage due to urinary loss of tricarboxylic acid cycle intermediates and increased rates of MCFAs oxidation | 52 abdominally overweight participants | The work analyses the metabolomics profile of patients subjected to different types of diet without distinguishing between patients with metabolic syndrome (or other traits of the latter) and healthy patients |
| Boden et al. | 2015 | Excessive caloric intake acutely causes oxidative stress, GLUT4 carbonylation, and insulin resistance in healthy men | 3 subjects normal weight (BMI 23.0, 23.9, and 24.9) and 3 overweight (BMI 26.8, 28.7, and 28.1) | The study is not using a metabolomics approach to identify compounds |
| Chen et al. | 2018 | Serum metabolomics model and its metabolic characteristics in patients with different syndromes of dyslipidemia based on nuclear magnetic resonance | 60 dyslipidemia patients (30 patients with SKYD and 30 patients with PDR) and 20 healthy subjects | Despite the study using a metabolomics approach, exclusively analyses patients with secondary dyslipidemia in SKYD (Spleen and Kidney Yang Deficiency) and PDR (Phlegm-Dampness Retention) |
| Chen et al. | 2019 | Trimethylamine N-Oxide Binds and Activates PERK to Promote Metabolic Dysfunction | Mouse model and cell culture | The study is not using a metabolomics approach to identify compounds; The study was not conducted on humans |
| Grün et al. | 2018 | High-Density Lipoprotein Reduction Differentially Modulates to Classical and Nonclassical Monocyte Subpopulations in Metabolic Syndrome Patients and in LPS-Stimulated Primary Human Monocytes In Vitro | 86 women and men between 20 and 60 years old | The study analyses the differences in monocyte subpopulations in patients with metabolic syndrome using flow cytometry (absence of a metabolomics approach) |
| Heidenreich et al. | 2017 | Retinol saturase coordinates liver metabolism by regulating ChREBP activity | Mouse model and cell culture | The study was not conducted on humans |
| Kempinska et al. | 2019 | The Association between SOCS11656G>A Polymorphism, Insulin Resistance and Obesity in Nonalcoholic Fatty Liver Disease (NAFLD) Patients | 138 patients with features of NAFLD 1000 controls | The study analyses the presence of genetic polymorphisms associated with metabolic syndrome and insulin resistance without using a metabolomics approach for the identification of metabolites |
| Kozyra et al. | 2018 | Human hepatic 3D spheroids as a model for steatosis and insulin resistance | Human hepatocyte 3D spheroid cultures | Study conducted on primary human hepatocyte 3D spheroid cultures |
| Leboucher et al. | 2019 | The translational regulator FMRP controls lipid and glucose metabolism in mice and humans | 25 fragile X patients and 29 sex- and age-matched healthy subjects | The study analyses exclusively people with fragile X syndrome to assess how the absence of specific genes may influence the metabolic homeostasis; No metabolomics approaches were used |
| Silvestri et al. | 2015 | Two non-psychoactive cannabinoids reduce intracellular lipid levels and inhibit hepatosteatosis | Mouse model and cell culture | The study was not conducted on humans |
| Teslovich et al. | 2018 | Identification of seven novel loci associated with amino acid levels using single-variant and gene-based tests in 8545 Finnish men from the METSIM study | 8545 non-diabetic men of mean age 57.3 ± 7.1 years | The study identifies genetic variants related to amino acid alterations in patients with metabolic syndrome |
| Zimmermann et al. | 2010 | Alterations in lipid, carbohydrate, and iron metabolism in patients with non-alcoholic steatohepatitis (NASH) and metabolic syndrome | 37 patients with metabolic syndrome (25 NASH and 12 non-NASH) 37 controls | The study is not using a metabolomics approach to identify compounds |
| Author | Years | Sample Size | Biofluid | Technique | Biomarkers Candidates | Results Increase (↑) or Decrease (↓) Biomarker Concentration Compared to the Control Group | Comments |
|---|---|---|---|---|---|---|---|
| Bhupathiraju et al. [14] | 2018 | 145 participants (aged 45 to 79 years) Control: Vegetarian dietary pattern | Serum | TQD MS/MS | Leucine; Valine; Tyrosine Methionine; Medium (C8–C14) to long-chain (C16–C20) acylcarnitines; Short-chain (C2 and C5) acylcarnitines | BCAAs ↑ Tyrosine ↓ Methionine ↓ Medium (C8–C14) to long-chain (C16–C20) acylcarnitines ↑ Short-chain (C2 and C5) acylcarnitines ↑ | Higher scores on the BCAA, aromatic amino acid, and short-chain AC metabolomic pattern were significantly associated with higher fasting insulin and 2-h insulin concentrations |
| Feldman et al. [15] | 2018 | 183 subjects: 62 obese NAFLD Controls: 69 lean controls 50 obese healthy | Serum | LC-MS/MS | Isoleucine; Leucine; Valine; Sphingolipids (SM); Phosphatidylcholine | In obese NAFLD: Isoleucine ↑ Leucine ↑ Valine ↑ Sphingolipids ↓ Phosphatidylcholine ↓ | SM OH C14:1 SM OH C16:1 SM OH C22:2 SM C16:0 |
| Lovric et al. [18] | 2018 | 37 subjects with MetS No Control Group | Plasma | UPLC/MS/MS GC/MS | Cholesterol ester (CE); Triacylglycerol (TAG); Phosphatidylcholine (PC); Glucose; Lysophosphatidylcholine (LPC); Sphingomyelin (SM); DAG (16:0/18:2) *; PE (16:0/18:1) **; PI (18:0/20:4) *** | CE 18:2 ↓ CE 20:3 ↑ DAG (16:0/18:2) ↑ SM (18:0) ↑ PC (38:3) ↑ PE (16:0/18:1) ↑ PI (18:0/20:4) ↑ TAG (50:4) ↓ TAGs containing FA with C48 to C50 ↑ TAGs containing FA with C52 to C58 ↓ Glucose ↑ | Ectopic fat (myocardial, epicardial, pericardial, and liver) was measured in non-diabetic male subjects with NAFLD Trend of plasma metabolites correlated with the increase in fat levels in ectopic fat deposits |
| Männisto et al. [11] | 2014 | 76 obese individuals: 32 with normal liver, 19 with simple steatosis 25 with NASH Controls: 32 with normal liver | Serum | 1H-NMR | Alanine; Histidine; Isoleucine; Leucine; Phenylalanine; Tyrosine; Valine; Citrate; β-hydroxybutyrte (β-HB); Acetoacetate | Alanine ↑ (NASH) Histidine ↑ (NASH) Isoleucine ↑ (NASH) Leucine ↑ (NASH) Phenylalanine ↑ (NASH) Valine ↑ (NASH) Citrate ↑ (NASH) β-HB ↓ (NASH) Acetoacetate ↓ (NASH) | β-HB decreases in the NASH compared to NAFLD Low levels of ketone bodies were associated with liver cell injury (ballooning) |
| Masarone et al. [17] | 2021 | 69 Controls 78 NAFLD 23 NASH 15 NASH-cirrhosis | Serum | GC-MS | Glycocholic acid; Taurocholic acid; Phenylalanine; BCAAs; Glutathione | Glycocholic acid ↑ (NASH) Taurocholic acid ↑ (NASH) Phenylalanine ↑ (NAFLD and NASH) BCAAs ↑ (NAFLD and NASH and NASH-cirrhosis) Glutathione ↓ (NAFLD and NASH) | |
| Ni et al. [19] | 2015 | 132 healthy subjects with normal weight (NW). 107 overweight/obese metabolically healthy (HO) 73 overweight/obese diabetic individuals (UO) | Serum | UPLC-QTOF-MS | Palmitoleic acid (PA) Dihomo-γ-linolenic acid (DGLA) | PA ↑ DGLA ↑ (UO) | Palmitoleic acid increase in UO subjects compared to HO subjects |
| Sookoian et al. [13] | 2016 | 32 patients with NAFLD 16 healthy controls | Serum | HPLC-MS | Alanine:pyruvate ratio; Kynurenine; Methionine; Taurine; Glucose; Glucose-6-phosphate; Lactic acid; Citraconic acid; Fumaric acid; Methyladenosine; N2, dimethylguanosine | Alanine:pyruvate ratio ↑ Kynurenine ↑ Methionine ↑ Taurine ↓ Glucose ↑ Glucose-6-phosphate ↓ Lactic acid ↑ Citraconic acid ↓ Fumaric acid ↓ Methyladenosine ↑ N2, dimethylguanosine ↑ | |
| Stechemesser et al. [12] | 2016 | 53 healthy controls 54 MetS without hyperferritinemia (MetS-Fe) 56 MetS with hyperferritinemia (MetS+Fe) | Serum | LC-MS/MS | Alanine; Citrulline; Glutamate; Kynurenine; Leucine; Sarcosine; Valine; Long-chain PCs; | Alanine ↑ Citrulline ↑ (MetS+Fe) Glutamate ↑ Kynurenine ↑ Leucine ↑ Sarcosine ↑ (MetS+Fe) Valine ↑ Long-chain PCs ↑ (MetS+Fe) | PC 40:2 PC 40:3 PC 40:4 PC 42:1 |
| Troisi et al. [16] | 2019 | 23 obese patients (15 with hepatic steatosis (St+); 8 without hepatic steatosis (St–); 10 with MetS; 13 without MetS) 18 normal weight healthy controls | Saliva | GC-MS | Isoleucine; Urea; Aconitic acid; Erythrose; Gluconic acid; Maltose; N-acetylgalactosamine; Lauric acid; Palmitic acid; Citraconic acid; Hydroxybutyric acid; Malic acid; Methylmalonic acid; Myristic acid | Isoleucine ↑ Urea ↑ Aconitic acid ↑ (Obese with MetS) Erythrose ↓ (Obese with MetS) Gluconic acid ↑ Maltose ↓ (with NAFLD) N-acetylgalactosamine ↑ Lauric acid ↓ (with NAFLD) Palmitic acid ↑ Citraconic acid ↓ Hydroxybutyric acid ↓ (without NAFLD) Malic acid ↓ (without NAFLD) Methylmalonic acid ↓ (Obese with MetS) Myristic acid ↑ | |
| Zhong et al. [21] | 2019 | 22 livers with NAFLD or NASH 50 normal liver | Liver Samples | LC-MS/MS | all-trans-retinoic acid (atRA); 13-cisRA; 4-oxo-arRA; all-trans-retinyl palmitate-d4 (RP) | RP ↓ (NASH) atRA ↓ (NASH) 13-cisRA ↓ 4-oxo-arRA ↓ | 4-oxo-arRA decrease in both NASH and NAFLD |
| Zhou et al. [20] | 2016 | 117 with NAFLD 69 NASH Control group | Plasma | UPLC-MS | Lysophosphatidylcholine 16:0 (LysoPC 16:0); Sphingomyelin; Unsaturated triacylglycerols (UnTGs) | LysoPC 16:0 ↓ (NASH) Sphingomyelin ↓ (NASH) UnTGs ↑(NASH) |
| Metabolites | Biofluids | Potential Effects |
|---|---|---|
| BCCAs (isoleucine, leucine, and valine) | Serum | Potential predictors of insulin resistance, fasting blood glucose level, and TG concentration |
| Kynurenine | Serum | The increase of Kynurenine associated with apoptosis and pro-oxidant effect (inflammation) |
| Glucose | Serum | The hyperglycemia condition is one of the main causes in the development of insulin resistance |
| Lactate | Serum | Precursor of gluconeogenesis during anaerobic glycolysis. Pathway overexpressed in the metabolic syndrome |
| Phosphatidylcoline (PC) | Serum | The reduction in CP levels appears to be caused by an increase in the turnover and size of adipocytes |
| Sphingolipids (sphingomyelin, ceramides) | Serum | The alteration of the sphingolipids seems to promote cellular stress, mitochondrial dysfunction, and alteration of the insulin-signalling pathway |
| DGLA (dihomo-gamma-linolenic acid) | Serum | Obese status was positively correlated with DGLA. DGLA is a good inflammation marker, useful in monitoring the metabolic status of overweight/obese individuals |
| Palmitoleic acid (PA) | Serum | An increase in PA is associated with an increase in de novo lipogenesis, which leads to an increase in DAG synthesis |
| Ketone bodies | Serum | A reduction in β-hydroxybutyrate levels seems to inhibit the NLRP3 inflammasome and reduce the production of IL1beta and IL-18. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Piras, C.; Noto, A.; Ibba, L.; Deidda, M.; Fanos, V.; Muntoni, S.; Leoni, V.P.; Atzori, L. Contribution of Metabolomics to the Understanding of NAFLD and NASH Syndromes: A Systematic Review. Metabolites 2021, 11, 694. https://doi.org/10.3390/metabo11100694
Piras C, Noto A, Ibba L, Deidda M, Fanos V, Muntoni S, Leoni VP, Atzori L. Contribution of Metabolomics to the Understanding of NAFLD and NASH Syndromes: A Systematic Review. Metabolites. 2021; 11(10):694. https://doi.org/10.3390/metabo11100694
Chicago/Turabian StylePiras, Cristina, Antonio Noto, Luciano Ibba, Martino Deidda, Vassilios Fanos, Sandro Muntoni, Vera Piera Leoni, and Luigi Atzori. 2021. "Contribution of Metabolomics to the Understanding of NAFLD and NASH Syndromes: A Systematic Review" Metabolites 11, no. 10: 694. https://doi.org/10.3390/metabo11100694
APA StylePiras, C., Noto, A., Ibba, L., Deidda, M., Fanos, V., Muntoni, S., Leoni, V. P., & Atzori, L. (2021). Contribution of Metabolomics to the Understanding of NAFLD and NASH Syndromes: A Systematic Review. Metabolites, 11(10), 694. https://doi.org/10.3390/metabo11100694